
Molecular Characterization of the 2020 Outbreak of Lumpy Skin Disease in Nepal

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Sample Collection
2.2. Sample Processing and DNA Extraction
2.3. Molecular Detection and Differential Diagnosis
2.4. Amplification of Selected Genes and Sequencing
2.5. Sequence and Phylogenetic Analysis
2.6. Targeted Nanopore Sequencing of Short PCR Fragments in the Buffalo Serum
3. Results
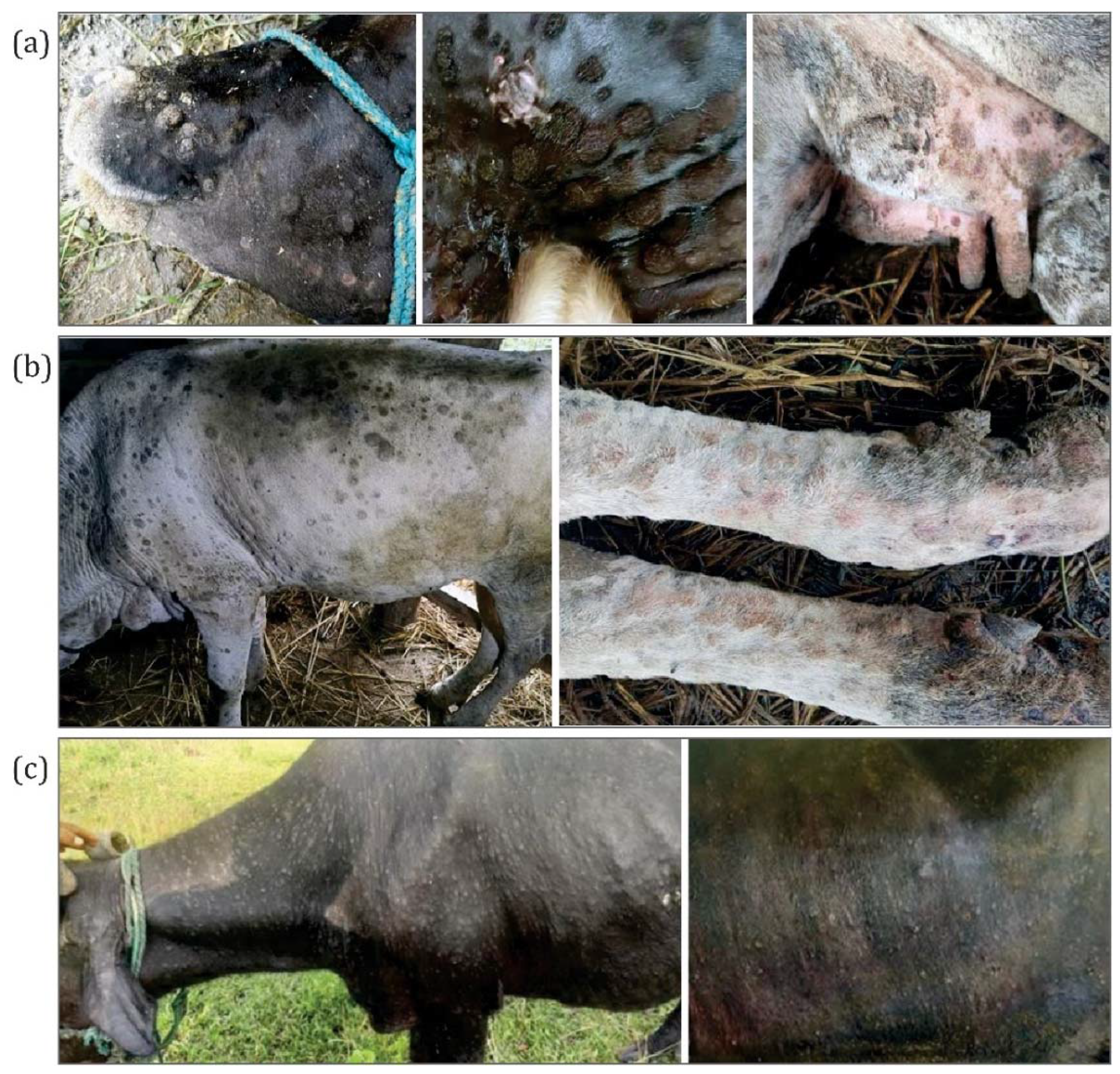
3.1. Clinical Signs and Symptoms and Outbreak Investigations
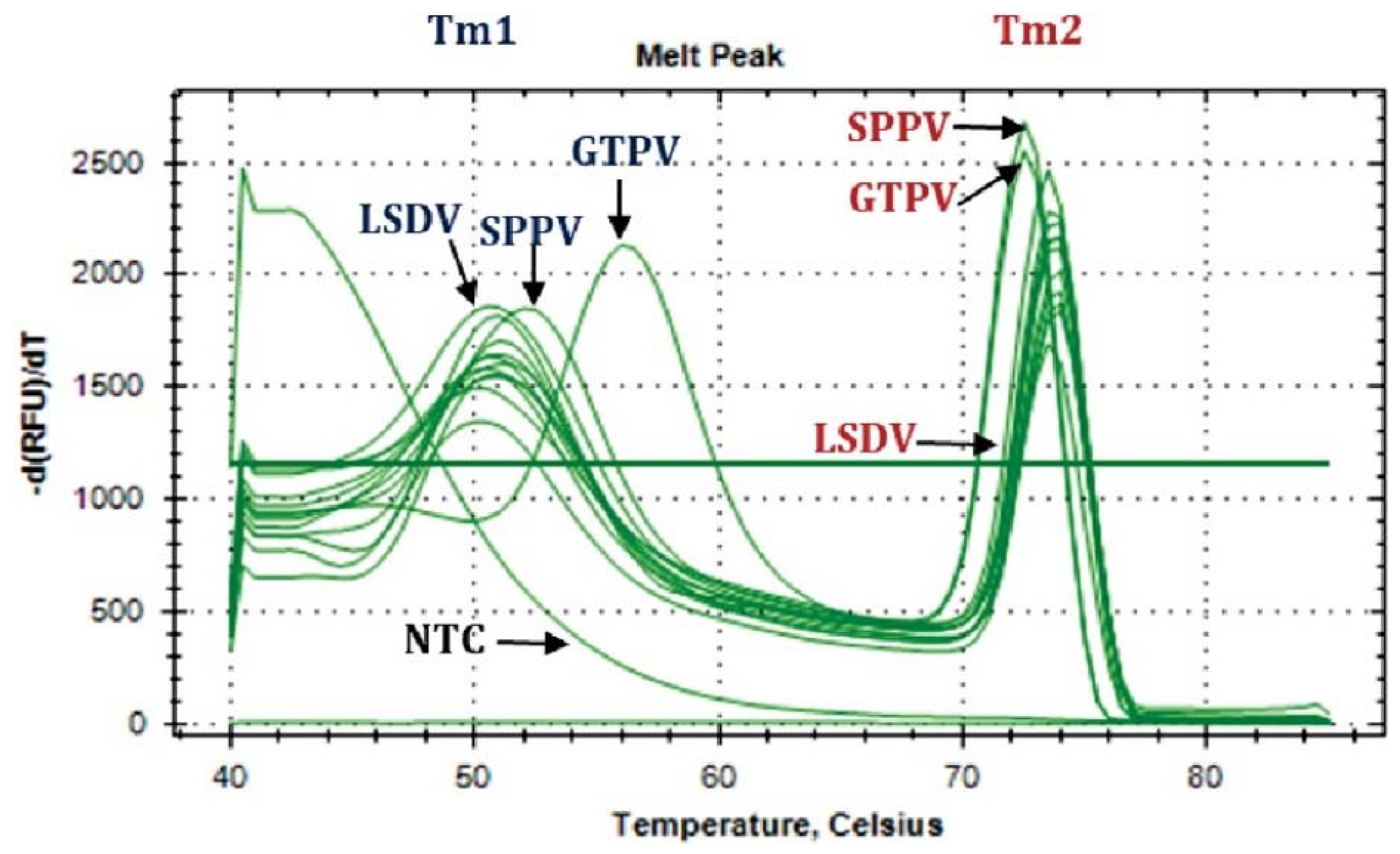
3.2. Molecular Detection and Diagnosis of LSDV
3.3. Amplification and Sequencing
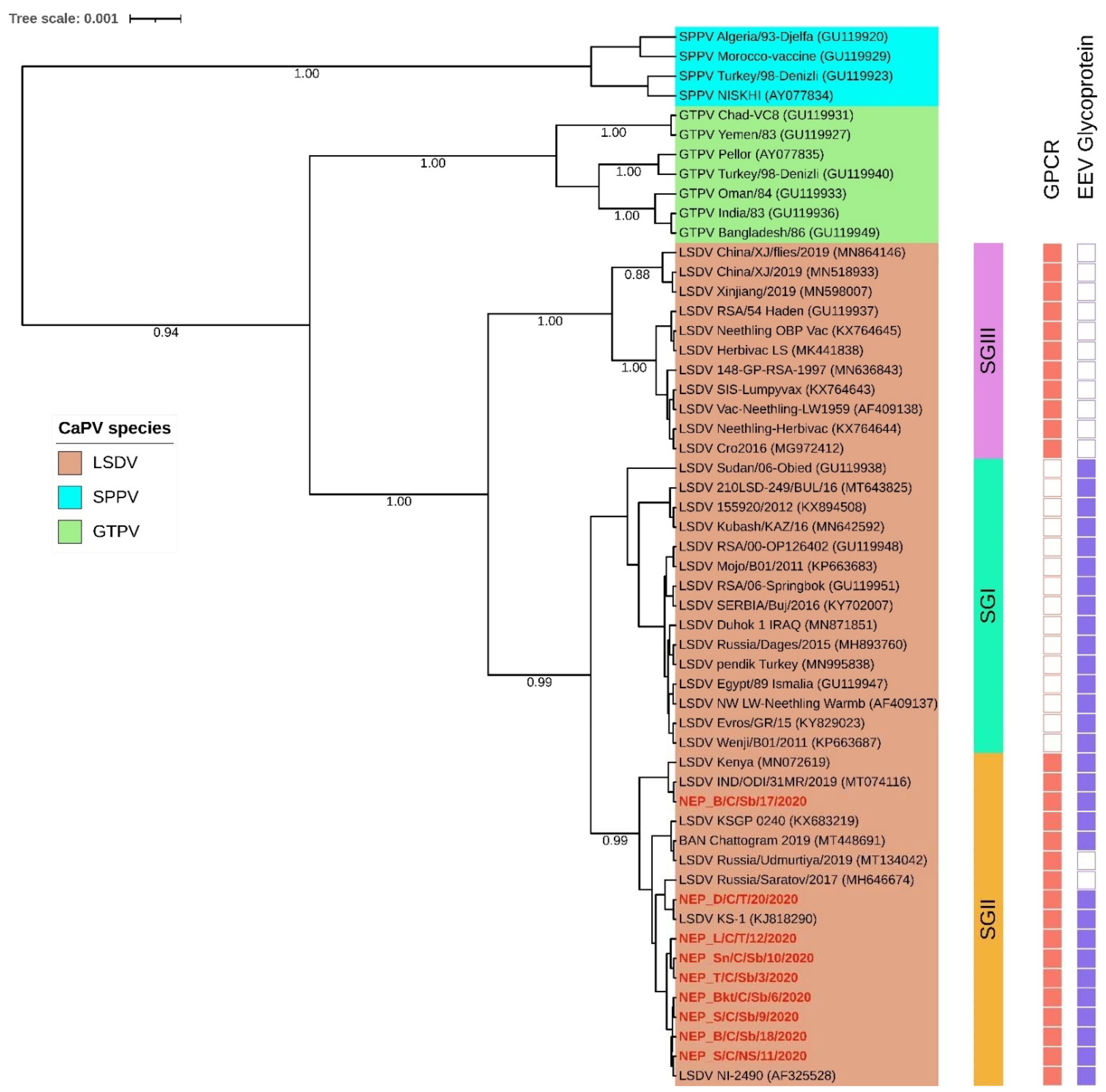
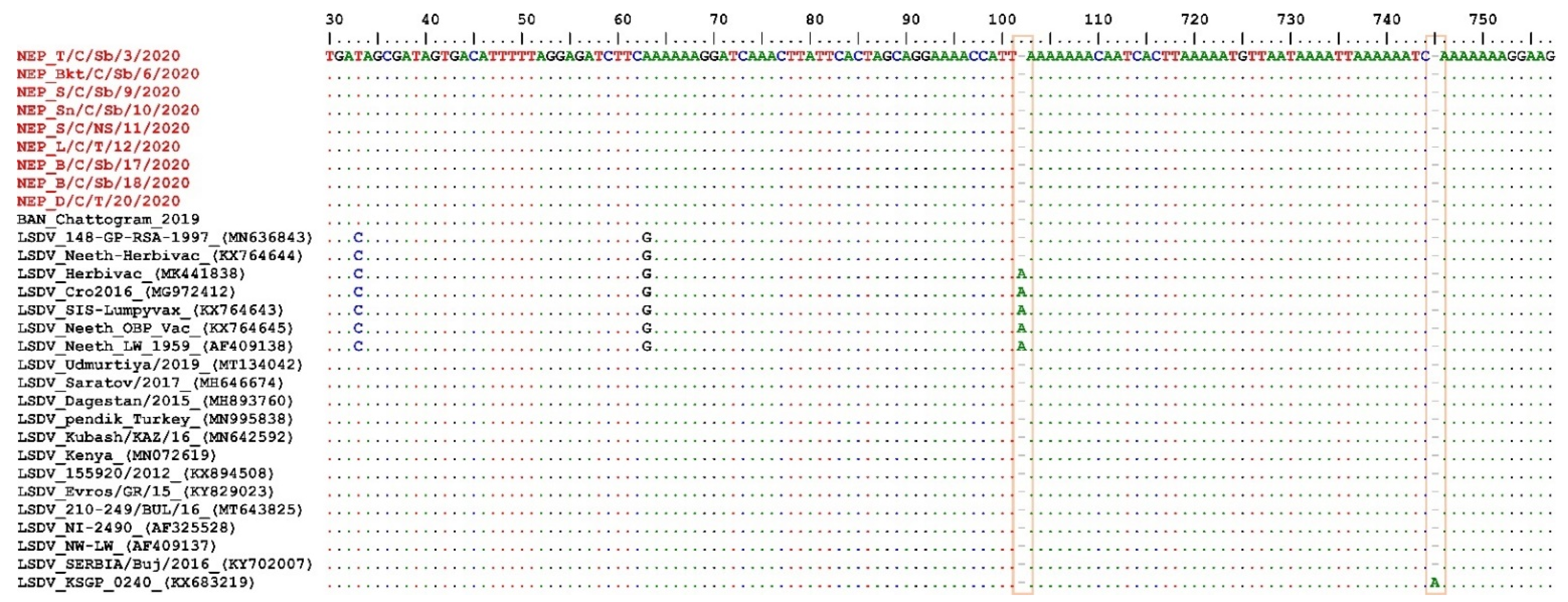
3.4. Sequence and Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Statement
References
- Gari, G.; Bonnet, P.; Roger, F.; Waret-Szkuta, A. Epidemiological aspects and financial impact of lumpy skin disease in Ethiopia. Prev. Vet. Med. 2011, 102, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Davies, F.G. Lumpy skin disease, an African capripox virus disease of cattle. Br. Vet. J. 1991, 147, 489–503. [Google Scholar] [CrossRef]
- Abutarbush, S.M.; Tuppurainen, E.S.M. Serological and clinical evaluation of the Yugoslavian RM65 sheep pox strain vaccine use in cattle against lumpy skin disease. Transbound. Emerg. Dis. 2018, 65, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- Gelaye, E.; Lamien, C.E. Lumpy skin disease and vectors of LSDV. In Transboundary Animal Diseases in Sahelian Africa and Connected Regions; Springer: Cham, Switzerland, 2019; pp. 267–288. ISBN 9783030253851. [Google Scholar]
- Sprygin, A.; Pestova, Y.; Wallace, D.B.; Tuppurainen, E.; Kononov, A.V. Transmission of lumpy skin disease virus: A short review. Virus Res. 2019, 269, 197637. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.S.M.; Stoltsz, W.H.; Troskie, M.; Wallace, D.B.; Oura, C.A.L.; Mellor, P.S.; Coetzer, J.A.W.; Venter, E.H. A Potential Role for Ixodid (Hard) Tick Vectors in the Transmission of Lumpy Skin Disease Virus in Cattle. Transbound. Emerg. Dis. 2011, 58, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.S.M.; Venter, E.H.; Shisler, J.L.; Gari, G.; Mekonnen, G.A.; Juleff, N.; Lyons, N.A.; De Clercq, K.; Upton, C.; Bowden, T.R.; et al. Review: Capripoxvirus Diseases: Current Status and Opportunities for Control. Transbound. Emerg. Dis. 2017, 64, 729–745. [Google Scholar] [CrossRef]
- Babiuk, S.; Bowden, T.R.; Boyle, D.B.; Wallace, D.B.; Kitching, R.P. Capripoxviruses: An emerging worldwide threat to sheep, goats and cattle. Transbound. Emerg. Dis. 2008, 55, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Molla, W.; de Jong, M.C.M.; Gari, G.; Frankena, K. Economic impact of lumpy skin disease and cost effectiveness of vaccination for the control of outbreaks in Ethiopia. Prev. Vet. Med. 2017, 147, 100–107. [Google Scholar] [CrossRef]
- Alkhamis, M.A.; VanderWaal, K. Spatial and temporal epidemiology of lumpy skin disease in the Middle East, 2012–2015. Front. Vet. Sci. 2016, 3, 19. [Google Scholar] [CrossRef]
- Van Schalkwyk, A.; Byadovskaya, O.; Shumilova, I.; Wallace, D.B.; Sprygin, A. Estimating evolutionary changes between highly passaged and original parental lumpy skin disease virus strains. Transbound. Emerg. Dis. 2021, in press. [Google Scholar] [CrossRef]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. Genome of Lumpy Skin Disease Virus. J. Virol. 2001, 75, 7122–7130. [Google Scholar] [CrossRef] [PubMed]
- Douglass, N.; Van Der Walt, A.; Omar, R.; Munyanduki, H.; Williamson, A.L. The complete genome sequence of the lumpy skin disease virus vaccine Herbivac LS reveals a mutation in the superoxide dismutase gene homolog. Arch. Virol. 2019, 164, 3107–3109. [Google Scholar] [CrossRef] [PubMed]
- Sprygin, A.; Babin, Y.; Pestova, Y.; Kononova, S.; Wallace, D.B.; Van Schalkwyk, A.; Byadovskaya, O.; Diev, V.; Lozovoy, D.; Kononov, A. Analysis and insights into recombination signals in lumpy skin disease virus recovered in the field. PLoS ONE 2018, 13, e0207480. [Google Scholar] [CrossRef]
- Sprygin, A.; Pestova, Y.; Bjadovskaya, O.; Prutnikov, P.; Zinyakov, N.; Kononova, S.; Ruchnova, O.; Lozovoy, D.; Chvala, I.; Kononov, A. Evidence of recombination of vaccine strains of lumpy skin disease virus with field strains, causing disease. PLoS ONE 2020, 15, e0207480. [Google Scholar] [CrossRef] [PubMed]
- Sprygin, A.; Van Schalkwyk, A.; Shumilova, I.; Nesterov, A.; Kononova, S.; Prutnikov, P.; Byadovskaya, O.; Kononov, A. Full-length genome characterization of a novel recombinant vaccine-like lumpy skin disease virus strain detected during the climatic winter in Russia, 2019. Arch. Virol. 2020, 165, 2675–2677. [Google Scholar] [CrossRef]
- Kononova, S.; Kononov, A.; Shumilova, I.; Byadovskaya, O.; Nesterov, A.; Prutnikov, P.; Babiuk, S.; Sprygin, A. A lumpy skin disease virus which underwent a recombination event demonstrates more aggressive growth in primary cells and cattle than the classical field isolate. Transbound. Emerg. Dis. 2021, 68, 1377–1383. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.M.; Oura, C.A.L. Review: Lumpy Skin Disease: An Emerging Threat to Europe, the Middle East and Asia. Transbound. Emerg. Dis. 2012, 59, 40–48. [Google Scholar] [CrossRef]
- Gelaye, E.; Mach, L.; Kolodziejek, J.; Grabherr, R.; Loitsch, A.; Achenbach, J.E.; Nowotny, N.; Diallo, A.; Lamien, C.E. A novel HRM assay for the simultaneous detection and differentiation of eight poxviruses of medical and veterinary importance. Sci. Rep. 2017, 7, 42892. [Google Scholar] [CrossRef]
- Lamien, C.E.; Lelenta, M.; Goger, W.; Silber, R.; Tuppurainen, E.; Matijevic, M.; Luckins, A.G.; Diallo, A. Real time PCR method for simultaneous detection, quantitation and differentiation of capripoxviruses. J. Virol. Methods 2011, 171, 134–140. [Google Scholar] [CrossRef]
- Haegeman, A.; Zro, K.; Vandenbussche, F.; Demeestere, L.; Van Campe, W.; Ennaji, M.M.; De Clercq, K. Development and validation of three Capripoxvirus real-time PCRs for parallel testing. J. Virol. Methods 2013, 193, 446–451. [Google Scholar] [CrossRef]
- Stubbs, S.; Oura, C.A.L.; Henstock, M.; Bowden, T.R.; King, D.P.; Tuppurainen, E.S.M. Validation of a high-throughput real-time polymerase chain reaction assay for the detection of capripoxviral DNA. J. Virol. Methods 2012, 179, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus tissue tropism and shedding: A quantitative study in experimentally infected sheep and goats. Virology 2008, 371, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Ireland, D.C.; Binepal, Y.S. Improved detection of capripoxvirus in biopsy samples by PCR. J. Virol. Methods 1998, 74, 1–7. [Google Scholar] [CrossRef]
- Heine, H.G.; Stevens, M.P.; Foord, A.J.; Boyle, D.B. A capripoxvirus detection PCR and antibody ELISA based on the major antigen P32, the homolog of the vaccinia virus H3L gene. J. Immunol. Methods 1999, 227, 187–196. [Google Scholar] [CrossRef]
- Lamien, C.E.; Le Goff, C.; Silber, R.; Wallace, D.B.; Gulyaz, V.; Tuppurainen, E.; Madani, H.; Caufour, P.; Adam, T.; El Harrak, M.; et al. Use of the Capripoxvirus homologue of Vaccinia virus 30kDa RNA polymerase subunit (RPO30) gene as a novel diagnostic and genotyping target: Development of a classical PCR method to differentiate Goat poxvirus from Sheep poxvirus. Vet. Microbiol. 2011, 149, 30–39. [Google Scholar] [CrossRef]
- Le Goff, C.; Lamien, C.E.; Fakhfakh, E.; Chadeyras, A.; Aba-Adulugba, E.; Libeau, G.; Tuppurainen, E.; Wallace, D.B.; Adam, T.; Silber, R.; et al. Capripoxvirus G-protein-coupled chemokine receptor: A host-range gene suitable for virus animal origin discrimination. J. Gen. Virol. 2009, 90, 1967–1977. [Google Scholar] [CrossRef]
- Chibssa, T.R.; Settypalli, T.B.K.; Berguido, F.J.; Grabherr, R.; Loitsch, A.; Tuppurainen, E.; Nwankpa, N.; Tounkara, K.; Madani, H.; Omani, A.; et al. An HRM Assay to Differentiate Sheeppox Virus Vaccine Strains from Sheeppox Virus Field Isolates and other Capripoxvirus Species. Sci. Rep. 2019, 9, 6646. [Google Scholar] [CrossRef]
- Gelaye, E.; Lamien, C.E.; Silber, R.; Tuppurainen, E.S.M.; Grabherr, R.; Diallo, A. Development of a Cost-Effective Method for Capripoxvirus Genotyping Using Snapback Primer and dsDNA Intercalating Dye. PLoS ONE 2013, 8, e75971. [Google Scholar] [CrossRef]
- Menasherow, S.; Erster, O.; Rubinstein-Giuni, M.; Kovtunenko, A.; Eyngor, E.; Gelman, B.; Khinich, E.; Stram, Y. A high-resolution melting (HRM) assay for the differentiation between Israeli field and Neethling vaccine lumpy skin disease viruses. J. Virol. Methods 2016, 232, 12–15. [Google Scholar] [CrossRef]
- Acharya, K.P.; Subedi, D. First outbreak of lumpy skin disease in Nepal. Transbound. Emerg. Dis. 2020, 67, 2280–2281. [Google Scholar] [CrossRef]
- Badhy, S.C.; Chowdhury, M.G.A.; Settypalli, T.B.K.; Cattoli, G.; Lamien, C.E.; Fakir, M.A.U.; Akter, S.; Osmani, M.G.; Talukdar, F.; Begum, N.; et al. Molecular characterization of lumpy skin disease virus (LSDV) emerged in Bangladesh reveals unique genetic features compared to contemporary field strains. BMC Vet. Res. 2021, 17, 61. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Xie, J.; Luo, J.; Shao, R.; Jia, K.; Li, S. Lumpy skin disease outbreaks in China, since 3 August 2019. Transbound. Emerg. Dis. 2021, 68, 216–219. [Google Scholar] [CrossRef]
- Sudhakar, S.B.; Mishra, N.; Kalaiyarasu, S.; Jhade, S.K.; Hemadri, D.; Sood, R.; Bal, G.C.; Nayak, M.K.; Pradhan, S.K.; Singh, V.P. Lumpy skin disease (LSD) outbreaks in cattle in Odisha state, India in August 2019: Epidemiological features and molecular studies. Transbound. Emerg. Dis. 2020, 67, 2408–2422. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Das, M.; Chowdhury, M.S.R.; Akter, S.; Mondal, A.K.; Uddin, M.J.; Rahman, M.M.; Rahman, M.M. An updated review on lumpy skin disease: Perspective of southeast asian countries. J. Adv. Biotechnol. Exp. Ther. 2021, 4, 322–333. [Google Scholar] [CrossRef]
- Arjkumpa, O.; Suwannaboon, M.; Boonrod, M.; Punyawan, I.; Liangchaisiri, S.; Laobannue, P.; Lapchareonwong, C.; Sansri, C.; Kuatako, N.; Panyasomboonying, P.; et al. The First Lumpy Skin Disease Outbreak in Thailand (2021): Epidemiological Features and Spatio-Temporal Analysis. Front. Vet. Sci. 2022, 8, 799065. [Google Scholar] [CrossRef]
- Aleksandr, K.; Pavel, P.; Olga, B.; Svetlana, K.; Vladimir, R.; Yana, P.; Alexander, S. Emergence of a new lumpy skin disease virus variant in Kurgan Oblast, Russia, in 2018. Arch. Virol. 2020, 165, 1343–1356. [Google Scholar] [CrossRef]
- Tasioudi, K.E.; Antoniou, S.E.; Iliadou, P.; Sachpatzidis, A.; Plevraki, E.; Agianniotaki, E.I.; Fouki, C.; Mangana-Vougiouka, O.; Chondrokouki, E.; Dile, C. Emergence of Lumpy Skin Disease in Greece, 2015. Transbound. Emerg. Dis. 2016, 63, 260–265. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.M.; Venter, E.H.; Coetzer, J.A.W. The detection of lumpy skin disease virus in samples of experimentally infected cattle using different diagnostic techniques. Onderstepoort J. Vet. Res. 2005, 72, 153–164. [Google Scholar] [CrossRef]
- Zeynalova, S.; Asadov, K.; Guliyev, F.; Vatani, M.; Aliyev, V. Epizootology and molecular diagnosis of lumpy skin disease among livestock in Azerbaijan. Front. Microbiol. 2016, 7, 1022. [Google Scholar] [CrossRef]
- Yeruham, I.; Perl, S.; Nyska, A.; Abraham, A.; Davidson, M.; Haymovitch, M.; Zamir, O.; Grinstein, H. Adverse reactions in cattle to a capripox vaccine. Vet. Rec. 1994, 135, 330–332. [Google Scholar] [CrossRef]
- Abutarbush, S.M. Adverse reactions to field vaccination against lumpy skin disease in cattle. Int. J. Infect. Dis. 2016, 45, 164. [Google Scholar] [CrossRef][Green Version]
- Roche, X.; Rozstalnyy, A.; TagoPacheco, D.; Kamata, A.; Pittiglio, C.; Beltran-alcrudo, D.; Bisht, K.; Karki, S.; Kayamori, J.; Larfaoui, F.; et al. Introduction and Spread of Lumpy Skin Disease in South, East and Southeast Asia: Qualitative Risk Assessment and Management, FAO Animal Production and Health Papers; Paper 183; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| District | Administration Region | No. of Susceptible Cases | No. of Clinically Affected Cases | No. of Dead Cases | Morbidity Rate (%) | Species | Type of Sample | Collection Date | Sample ID |
|---|---|---|---|---|---|---|---|---|---|
| Sunsari | Province I | 3780 | 8 | 0 | 0.21 | Cattle | Scab | October 2020 | Sn/C/Sb/10/2020 |
| Sunsari | 728 | 2 | 0 | 0.27 | Buffalo | Serum | October 2020 | Sn/B/S/15/2020 | |
| Panchthar | 238 | 3 | 0 | 1.26 | Cattle | Serum | October 2020 | P/C/S/7/2020 | |
| Kathmandu | Bagmati Province | 809 | 4 | 0 | 0.49 | Cattle | Nasal swab | August 2020 | Kt/C/NS/4/2020 |
| Kathmandu | Cattle | Nasal swab | August 2020 | Kt/C/NS/5/2020 | |||||
| Bhaktapur | 758 | 2 | 0 | 0.26 | Cattle | Scab | October 2020 | Bkt/C/Sb/6/2020 | |
| Sindhuli | 1969 | 15 | 0 | 0.76 | Cattle | Scab | September 2020 | S/C/Sb/9/2020 | |
| Sindhuli | Cattle | Nasal swab | September 2020 | S/C/NS/11/2020 | |||||
| Kavre | 3987 | 3 | 0 | 0.075 | Buffalo | Serum | September 2020 | Kr/B/S/8/2020 | |
| Kaski | Gandaki Province | 4682 | 4 | 0 | 0.08 | Buffalo | Scab | August 2020 | K/B/Sb/1/2020 |
| Kaski | Buffalo | Scab | August 2020 | K/B/Sb/2/2020 | |||||
| Kaski | 3170 | 12 | 0 | 0.38 | Cattle | Serum | September 2020 | K/C/S/16/2020 | |
| Tanahu | 4164 | 2 | 0 | 0.05 | Cattle | Scab | August 2020 | T/C/Sb/3/2020 | |
| Lamjung | 1100 | 1 | 0 | 0.09 | Cattle | Tissue | September 2020 | L/C/T/12/2020 | |
| Nawalpur | 1799 | 9 | 0 | 0.50 | Cattle | Serum | September 2020 | N/C/S/14/2020 | |
| Rupandehi | Lumbini Province | 5500 | 5 | 0 | 0.09 | Cattle | Ocular swab | September 2020 | R/C/OS/13/2020 |
| Banke | 1623 | 28 | 0 | 1.72 | Cattle | Scab swab | October 2020 | B/C/Sb/17/2020 | |
| Banke | Cattle | Scab swab | October 2020 | B/C/Sb/18/2020 | |||||
| Dang | 5958 | 3 | 0 | 0.05 | Cattle | Tissue (Skin nodules) | October 2020 | D/C/T/19/2020 | |
| Dang | Cattle | Tissue (Skin nodules) | October 2020 | D/C/T/20/2020 | |||||
| Total | 30868 | 92 | 0 | 0.3 | Cattle | ||||
| 9397 | 9 | 0 | 0.09 | Buffalo | |||||
| Target Gene | Primer | Amplicon Size |
|---|---|---|
| RPO30 Snapback | Cp-HRM-SBF-5′-ggTGTAGTACGTATAAGATTATCGTATAGAAACAAGCCTTTA-3′ | SNAP BACK assay |
| Cp-HRM1R-5′-AATTTCTTTCTCTGTTCCATTTG-3′ | ||
| GPCR_ FRET | CpRt Forward- 5′-GATAGTATCGCTAAACAATGG-3′ | 200 bp |
| CpRt Reverse- 5′-ATCCAAACCACCATACTAAG-3′ | ||
| RPO30 | Cp-OL1F 5′-CAGCTGTTTGTTTACATTTGATTTTT-3′ | 554 bp |
| Cp-OL1R 5′-TCGTATAGAAACAAGCCTTTAATAGA-3′ | ||
| Cp-OL2F 5′-TTTGAACACATTTTATTCCAAAAAG-3′ | 520 bp | |
| Cp-OL2R 5′-AACCTACATGCATAAACAGAAGC-3′ | ||
| GPCR | CpGPCR-OL1F-5′-TGAAAAATTAATCCATTCTTCTAAACA-3′ | 684 bp |
| CpGPCR-OL1R-5′-TCATGTATTTTATAACGATAATGCAAA-3′ | ||
| CpGPCR-OL2F-5′-TTAGCGGTATAATCATTCCAAATA -3′ | 603 bp | |
| CpGPCR-OL2R-5′-GCGATGATTATGATGATTATGAAGTG-3‘ | ||
| CpGPCR-OL3F-5′-CACAATTATATTTCCAAATAATCCAA -3′ | 617 bp | |
| CpGPCR-OL3R-5′-TGTACATGTGTAATTTTAATGTTCGTA-3′ | ||
| EEV glycoprotein gene (ORF LSDV126) | EEVGly F- 5′-ATGGGAATAGTATCTGTTGTATACG-3′ | 250 bp |
| EEVGly R-5′-CGAACCCCTATTTACTTGAGAA-3′ | ||
| B22R | B22R F- 5′-TCATTTTCTTCTAGTTCCGACGA-3′ | 863 bp |
| B22R R- 5′-TTCGTTGATGATAAATAACTGGAAA-3′ |
| S. No. | Sample ID | Host | SNAP BACK Assay Analysis | Remarks | ||
|---|---|---|---|---|---|---|
| Cq | Tm1 (°C) | Tm2 (°C) | ||||
| 1 | K/B/Sb/1/2020 | Buffalo | N/A | N/A | N/A | Negative |
| 2 | K/B/Sb/2/2020 | Buffalo | N/A | N/A | N/A | Negative |
| 3 | T/C/Sb/3/2020 | Cattle | 20.18 | 51.00 | 74.00 | LSDV |
| 4 | Kt/C/NS/4/2020 | Cattle | 28.73 | 50.50 | 74.00 | LSDV |
| 5 | Kt/C/NS/5/2020 | Cattle | N/A | N/A | N/A | Negative |
| 6 | Bkt/C/Sb/6/2020 | Cattle | 24.25 | 51.00 | 74.00 | LSDV |
| 7 | P/C/S/7/2020 | Cattle | 37.38 | 50.50 | 73.50 | LSDV |
| 8 | Kr/B/S/8/2020 | Buffalo | 24.86 | 50.00 | 73.50 | LSDV |
| 9 | S/C/Sb/9/2020 | Cattle | 22.48 | 51.00 | 74.00 | LSDV |
| 10 | Sn/C/Sb/10/2020 | Cattle | 19.83 | 51.00 | 74.00 | LSDV |
| 11 | S/C/NS/11/2020 | Cattle | 24.83 | 51.00 | 74.00 | LSDV |
| 12 | L/C/T/12/2020 | Cattle | 25.14 | 51.00 | 74.00 | LSDV |
| 13 | R/C/OS/13/2020 | Cattle | N/A | N/A | N/A | Negative |
| 14 | N/C/S/14/2020 | Cattle | N/A | N/A | N/A | Negative |
| 15 | Sn/B/S/15/2020 | Buffalo | 36.34 | 51.00 | 73.50 | LSDV |
| 16 | K/C/S/16/2020 | Cattle | N/A | N/A | N/A | Negative |
| 17 | B/C/Sb/17/2020 | Cattle | 24.97 | 51.00 | 74.00 | LSDV |
| 18 | B/C/Sb/18/2020 | Cattle | 29.28 | 50.50 | 74.00 | LSDV |
| 19 | D/C/T/19/2020 | Cattle | 31.02 | 50.00 | 73.50 | LSDV |
| 20 | D/C/T/20/2020 | Cattle | 22.87 | 50.50 | 74.00 | LSDV |
| GTPV | GTPV positive control | N/A | 27.82 | 56.00 | 72.50 | GTPV |
| SPPV | SPPV positive control | N/A | 27.88 | 52.00 | 72.50 | SPPV |
| LSDV | LSDV positive control | N/A | 24.90 | 50.50 | 73.50 | LSDV |
| NTC | RNAse free water | N/A | N/A | N/A | N/A | Negative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koirala, P.; Meki, I.K.; Maharjan, M.; Settypalli, B.K.; Manandhar, S.; Yadav, S.K.; Cattoli, G.; Lamien, C.E. Molecular Characterization of the 2020 Outbreak of Lumpy Skin Disease in Nepal. Microorganisms 2022, 10, 539. https://doi.org/10.3390/microorganisms10030539
Koirala P, Meki IK, Maharjan M, Settypalli BK, Manandhar S, Yadav SK, Cattoli G, Lamien CE. Molecular Characterization of the 2020 Outbreak of Lumpy Skin Disease in Nepal. Microorganisms. 2022; 10(3):539. https://doi.org/10.3390/microorganisms10030539
Chicago/Turabian StyleKoirala, Pragya, Irene Kasindi Meki, Manju Maharjan, Bharani Kumar Settypalli, Salina Manandhar, Sanjay Kumar Yadav, Giovanni Cattoli, and Charles Euloge Lamien. 2022. "Molecular Characterization of the 2020 Outbreak of Lumpy Skin Disease in Nepal" Microorganisms 10, no. 3: 539. https://doi.org/10.3390/microorganisms10030539
APA StyleKoirala, P., Meki, I. K., Maharjan, M., Settypalli, B. K., Manandhar, S., Yadav, S. K., Cattoli, G., & Lamien, C. E. (2022). Molecular Characterization of the 2020 Outbreak of Lumpy Skin Disease in Nepal. Microorganisms, 10(3), 539. https://doi.org/10.3390/microorganisms10030539