
Bacillus subtilis PS-216 Antagonistic Activities against Campylobacter jejuni NCTC 11168 Are Modulated by Temperature, Oxygen, and Growth Medium


 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Cocultivation of B. subtilis with C. jejuni in Mueller–Hinton Broth at Different Starting Ratios
2.3. Cocultivation of B. subtilis with C. jejuni in Mueller–Hinton Broth at Different Temperatures
2.4. Cocultivation of B. subtilis with C. jejuni in Mueller–Hinton Broth under Aerobic Conditions
2.5. Cocultivation of B. subtilis PS-216 and C. jejuni NCTC 11168 in Broiler Chicken Intestinal Content Medium
2.6. Growth of B. subtilis PS-216 in Nonsterile Broiler Chicken Intestinal Content Medium
2.7. Statistical Analysis
3. Results
3.1. B. subtilis PS-216 Reduces C. jejuni Growth
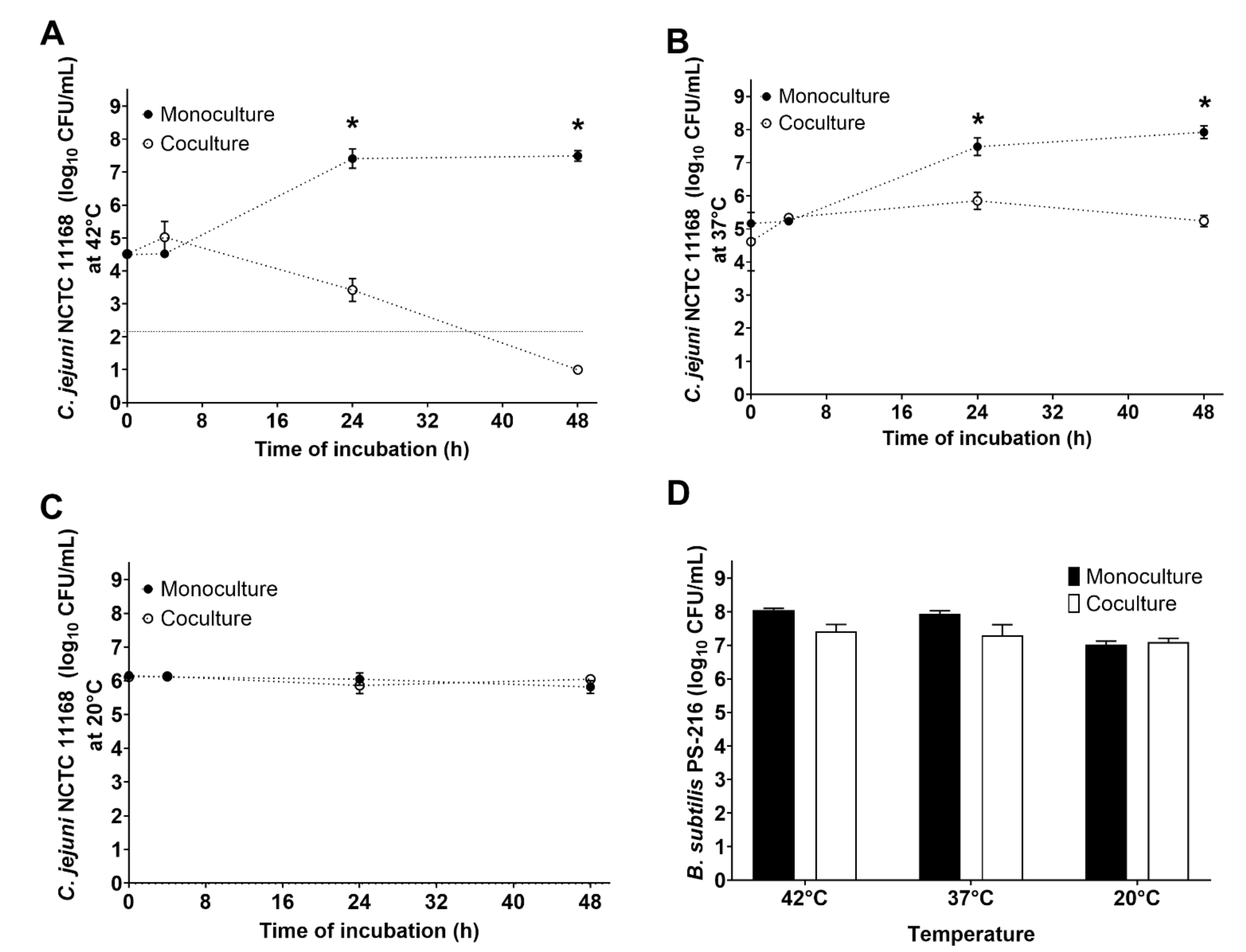
3.2. B. subtilis PS-216 Anti-Campylobacter Effects Increase at Higher Temperatures
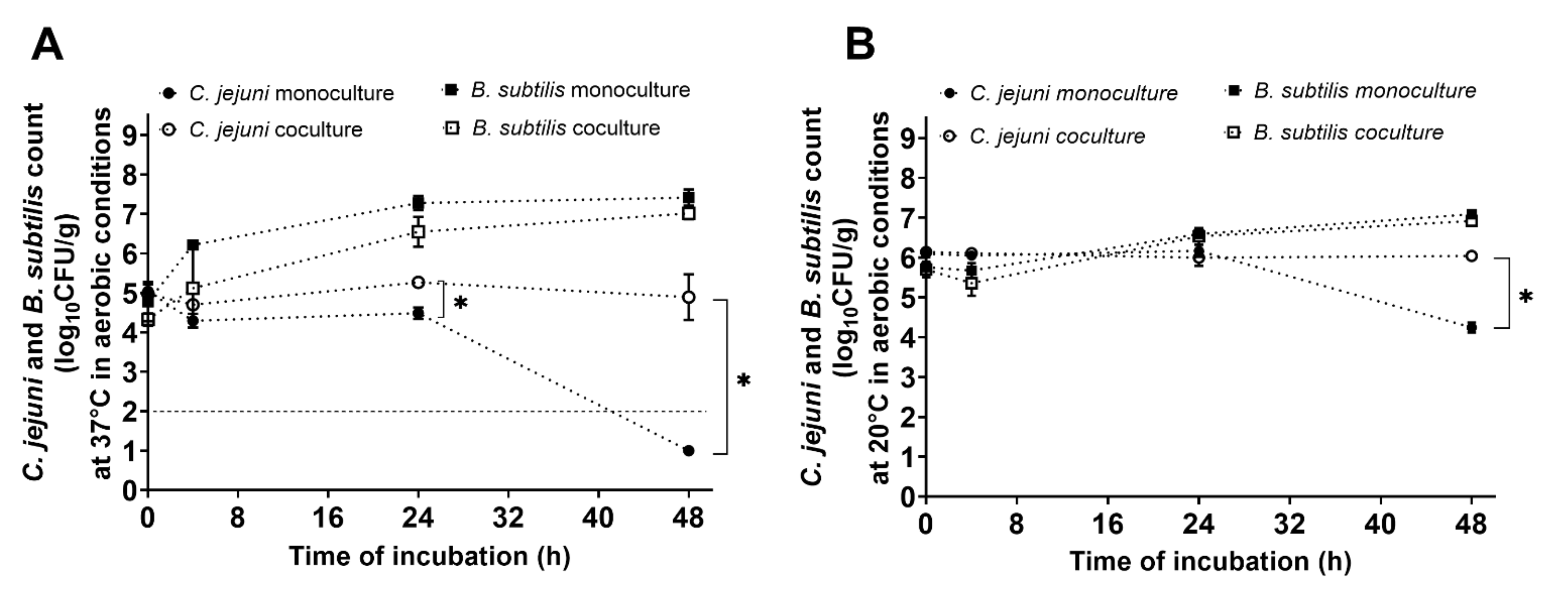
3.3. Protective Effects of B. subtilis PS-216 for C. jejuni under Aerobic Conditions
3.4. Anti-Campylobacter Effects of B. subtilis PS-216 Are More Prominent in Chicken Litter Than in Intestinal Content
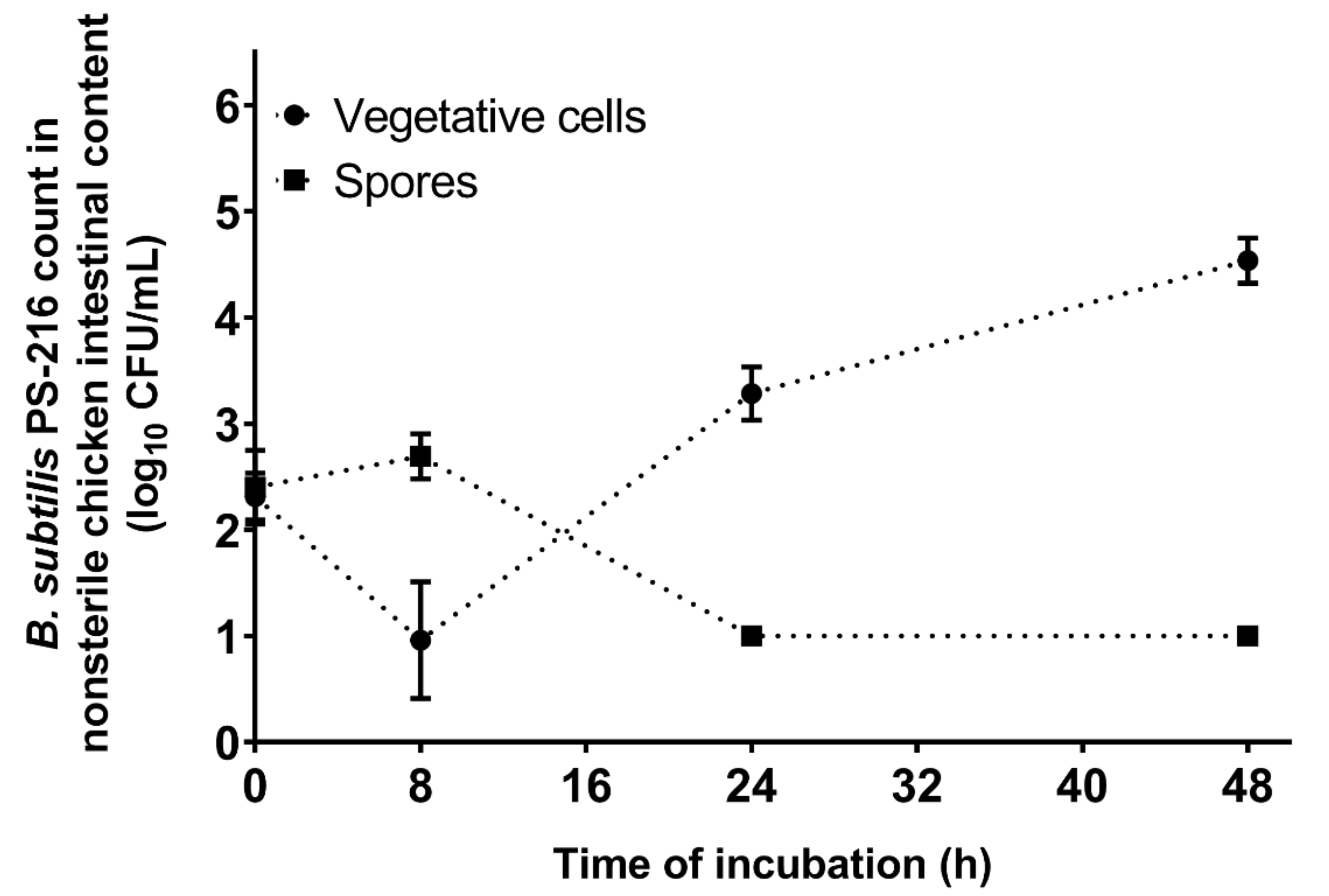
3.5. B. subtilis PS-216 Growth and Sporulation Are Limited in Nonsterile Chicken Intestinal Content Medium
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA. ECDC The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA and ECDC European Food Safety Authority and European Centre for Disease Prevention and Control. The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar] [CrossRef]
- Ellyn, P.; Marder, M.P.H.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Hurd, S.; Jervis, R.; Lathrop, S.; Muse, A.; Ryan, P.; et al. Preliminary Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2006–2017. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 324–328. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Davies, R.; Cesare, A.D.; Herman, L.; Hilbert, F.; Lindqvist, R.; et al. Update and Review of Control Options for Campylobacter in Broilers at Primary Production. EFSA J. 2020, 18, e06090. [Google Scholar] [CrossRef]
- Meunier, M.; Guyard-Nicodème, M.; Dory, D.; Chemaly, M. Control Strategies against Campylobacter at the Poultry Production Level: Biosecurity Measures, Feed Additives and Vaccination. J. Appl. Microbiol. 2016, 120, 1139–1173. [Google Scholar] [CrossRef] [Green Version]
- Dogan, O.B.; Clark, J.; Mattos, F.; Wang, B. A Quantitative Microbial Risk Assessment Model of Campylobacter in Broiler Chickens: Evaluating Processing Interventions. Food Control 2019, 100, 97–110. [Google Scholar] [CrossRef]
- Foster, K.R.; Bell, T. Competition, Not Cooperation, Dominates Interactions among Culturable Microbial Species. Curr. Biol. 2012, 22, 1845–1850. [Google Scholar] [CrossRef] [Green Version]
- Thomrongsuwannakij, T.; Chuanchuen, R.; Chansiripornchai, N. Identification of Competitive Exclusion and Its Ability to Protect against Campylobacter jejuni in Broilers. Thai J. Vet. Med. 2016, 46, 279–286. [Google Scholar]
- Schouls, L.M.; Reulen, S.; Duim, B.; Wagenaar, J.A.; Willems, R.J.L.; Dingle, K.E.; Colles, F.M.; Van Embden, J.D.A. Comparative Genotyping of Campylobacter jejuni by Amplified Fragment Length Polymorphism, Multilocus Sequence Typing, and Short Repeat Sequencing: Strain Diversity, Host Range, and Recombination. J. Clin. Microbiol. 2003, 41, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Stern, N.J. Mucosal Competitive Exclusion to Diminish Colonization of Chickens by Campylobacter jejuni. Poult. Sci. 1994, 73, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ma, L.; Doyle, M.P. Potential Competitive Exclusion Bacteria from Poultry Inhibitory to Campylobacter jejuni and Salmonella. J. Food Prot. 2007, 70, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Saint-Cyr, M.J.; Guyard-Nicodème, M.; Messaoudi, S.; Chemaly, M.; Cappelier, J.-M.; Dousset, X.; Haddad, N. Recent Advances in Screening of Anti-Campylobacter Activity in Probiotics for Use in Poultry. Front. Microbiol. 2016, 7, 553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mañes-Lázaro, R.; Van Diemen, P.M.; Pin, C.; Mayer, M.J.; Stevens, M.P.; Narbad, A. Administration of Lactobacillus Johnsonii FI9785 to Chickens Affects Colonisation by Campylobacter jejuni and the Intestinal Microbiota. Br. Poult. Sci. 2017, 58, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Wang, W.; Zhang, H.; Wang, J.; Zhang, W.; Gao, J.; Wu, S.; Qi, G. Supplemental Bacillus subtilis DSM 32315 Manipulates Intestinal Structure and Microbial Composition in Broiler Chickens. Sci. Rep. 2018, 8, 15358. [Google Scholar] [CrossRef] [Green Version]
- Neijat, M.; Shirley, R.B.; Welsher, A.; Barton, J.; Thiery, P.; Kiarie, E. Growth Performance, Apparent Retention of Components, and Excreta Dry Matter Content in Shaver White Pullets (5 to 16 Week of Age) in Response to Dietary Supplementation of Graded Levels of a Single Strain Bacillus subtilis Probiotic. Poult. Sci. 2019, 98, 3777–3786. [Google Scholar] [CrossRef]
- Tamehiro, N.; Okamoto-Hosoya, Y.; Okamoto, S.; Ubukata, M.; Hamada, M.; Naganawa, H.; Ochi, K. Bacilysocin, a Novel Phospholipid Antibiotic Produced by Bacillus subtilis 168. Antimicrob. Agents Chemother. 2002, 46, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.; Dang, S.; Gupta, S.; Gabrani, R. Antibacterial Activity, Cytotoxicity, and the Mechanism of Action of Bacteriocin from Bacillus subtilis GAS101. Med. Princ. Pract. 2018, 27, 186–192. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus subtilis Group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Khare, A.; Tavazoie, S. Multifactorial Competition and Resistance in a Two-Species Bacterial System. PLoS Genet. 2015, 11, e1005715. [Google Scholar] [CrossRef]
- Erega, A.; Stefanic, P.; Dogsa, I.; Danevčič, T.; Simunovic, K.; Klančnik, A.; Možina, S.S.; Mulec, I.M. Bacillaene Mediates the Inhibitory Effect of Bacillus subtilis on Campylobacter jejuni Biofilms. Appl. Environ. Microbiol. 2021, 87, e0295520. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Miller, W.G.; De Reuse, H.; Mendz, G.L. Oxygen Requirement and Tolerance of Campylobacter jejuni. Res. Microbiol. 2007, 158, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Culotti, A.; Packman, A.I. Pseudomonas Aeruginosa Facilitates Campylobacter jejuni Growth in Biofilms under Oxic Flow Conditions. FEMS Microbiol. Ecol. 2015, 91, fiv136. [Google Scholar] [CrossRef] [PubMed]
- Hilbert, F.; Scherwitzel, M.; Paulsen, P.; Szostak, M.P. Survival of Campylobacter jejuni under Conditions of Atmospheric Oxygen Tension with the Support of Pseudomonas Spp. Appl. Environ. Microbiol. 2010, 76, 5911–5917. [Google Scholar] [CrossRef] [Green Version]
- Rasschaert, G.; De Zutter, L.; Herman, L.; Heyndrickx, M. Campylobacter Contamination of Broilers: The Role of Transport and Slaughterhouse. Int. J. Food Microbiol. 2020, 322, 108564. [Google Scholar] [CrossRef]
- Nakano, M.M.; Zuber, P. Anaerobic Growth of a “Strict Aerobe” (Bacillus subtilis). Annu. Rev. Microbiol. 1998, 52, 165–190. [Google Scholar] [CrossRef]
- Nakano, M.M.; Hulett, F.M. Adaptation of Bacillus subtilis to Oxygen Limitation. FEMS Microbiol. Lett. 1997, 157, 1–7. [Google Scholar] [CrossRef]
- Hazeleger, W.C.; Wouters, J.A.; Rombouts, F.M.; Abee, T. Physiological Activity of Campylobacter jejuni Far below the Minimal Growth Temperature. Appl. Environ. Microbiol. 1998, 64, 3917–3922. [Google Scholar] [CrossRef] [Green Version]
- Satapute, P.; Olekar, H.S.; Shetti, A.; Kulkarni, A.; Hiremath, G.; Patagundi, B.I.; Shivsharan, C.T.; Kaliwal, B.B. Isolation and Characterization of Nitrogen Fixing Bacillus subtilis Strain As-4 from Agricultural Soil. Int. J. Recent Sci. Res. 2012, 3, 762–765. [Google Scholar]
- Isnawati; Trimulyono, G. Temperature Range and Degree of Acidity Growth of Isolate of Indigenous Bacteria on Fermented Feed “Fermege”. J. Phys. Conf. Ser. 2018, 953, 012209. [Google Scholar] [CrossRef]
- Perez, C.; Suarez, C.; Castro, G.R. Production of Antimicrobials by Bacillus subtilis MIR 15. J. Biotechnol. 1992, 26, 331–336. [Google Scholar] [CrossRef]
- Jiménez-Delgadillo, R.; Valdés-Rodríguez, S.E.; Olalde-Portugal, V.; Abraham-Juárez, R.; García-Hernández, J.L.; Jiménez-Delgadillo, R.; Valdés-Rodríguez, S.E.; Olalde-Portugal, V.; Abraham-Juárez, R.; García-Hernández, J.L. Effect of PH and Temperature on the Growth and Antagonistic Activity of Bacillus subtilis on Rhizoctonia Solani. Rev. Mex. Fitopatol. 2018, 36, 256–275. [Google Scholar] [CrossRef] [Green Version]
- Štefanič, P.; Mandič-Mulec, I. Social Interactions and Distribution of Bacillus subtilis Pherotypes at Microscale. J. Bacteriol. 2009, 191, 1756–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, N.J.; Robach, M.C. Enumeration of Campylobacter spp. in Broiler Feces and in Corresponding Processed Carcasses. J. Food Prot. 2003, 66, 1557–1563. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, G.; Troncoso, M.; López, C.; Rivas, P.; Toro, M. Occurrence and Enumeration of Campylobacter spp. during the Processing of Chilean Broilers. BMC Microbiol. 2009, 9, 94. [Google Scholar] [CrossRef] [Green Version]
- Ica, T.; Caner, V.; Istanbullu, O.; Nguyen, H.D.; Ahmed, B.; Call, D.R.; Beyenal, H. Characterization of Mono- and Mixed-Culture Campylobacter jejuni Biofilms. Appl. Environ. Microbiol. 2012, 78, 1033–1038. [Google Scholar] [CrossRef] [Green Version]
- Fritts, C.A.; Kersey, J.H.; Motl, M.A.; Kroger, E.C.; Yan, F.; Si, J.; Jiang, Q.; Campos, M.M.; Waldroup, A.L.; Waldroup, P.W. Bacillus subtilis C-3102 (Calsporin) Improves Live Performance and Microbiological Status of Broiler Chickens. J. Appl. Poult. Res. 2000, 9, 149–155. [Google Scholar] [CrossRef]
- Guyard-Nicodeme, M.; Keita, A.; Quesne, S.; Amelot, M.; Poezevara, T.; Le Berre, B.; Sanchez, J.; Vesseur, P.; Martin, A.; Medel, P.; et al. Efficacy of Feed Additives against Campylobacter in Live Broilers during the Entire Rearing Period. Poult. Sci. 2016, 95, 298–305. [Google Scholar] [CrossRef]
- Gracia, M.I.; Millan, C.; Sanchez, J.; Guyard-Nicodeme, M.; Mayot, J.; Carre, Y.; Csorbai, A.; Chemaly, M.; Medel, P. Efficacy of Feed Additives against Campylobacter in Live Broilers during the Entire Rearing Period: Part B. Poult. Sci. 2016, 95, 886–892. [Google Scholar] [CrossRef]
- Harada, T.; Tokai, Y.; Kimura, A.; Ikeda, S.; Matsumura, M. Hydrolysis of Crystalline Cellulose to Glucose in an Autoclave Containing Both Gaseous and Liquid Water. RSC Adv. 2014, 4, 26838–26842. [Google Scholar] [CrossRef]
- Manhar, A.K.; Bashir, Y.; Saikia, D.; Nath, D.; Gupta, K.; Konwar, B.K.; Kumar, R.; Namsa, N.D.; Mandal, M. Cellulolytic Potential of Probiotic Bacillus subtilis AMS6 Isolated from Traditional Fermented Soybean (Churpi): An in-Vitro Study with Regards to Application as an Animal Feed Additive. Microbiol. Res. 2016, 186–187, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-K.; Lee, S.-C.; Cho, Y.-Y.; Oh, H.-J.; Ko, Y.H. Isolation of Cellulolytic Bacillus subtilis Strains from Agricultural Environments. ISRN Microbiol. 2012, 2012, 650563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattossovich, R.; Iacono, R.; Cangiano, G.; Cobucci-Ponzano, B.; Isticato, R.; Moracci, M.; Ricca, E. Conversion of Xylan by Recyclable Spores of Bacillus subtilis Displaying Thermophilic Enzymes. Microbial. Cell Factories 2017, 16, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez-Lorenzo, L.; Porras-Domínguez, J.R.; Raga-Carbajal, E.; Olvera, C.; Rodríguez-Alegría, M.E.; Carrillo-Nava, E.; Costas, M.; Munguía, A.L. Intrinsic Levanase Activity of Bacillus subtilis 168 Levansucrase (SacB). PLoS ONE 2015, 10, e0143394. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Yu, Z. Intestinal Microbiome of Poultry and Its Interaction with Host and Diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Cartman, S.T.; Ragione, R.M.L.; Woodward, M.J. Bacillus subtilis Spores Germinate in the Chicken Gastrointestinal Tract. Appl. Environ. Microbiol. 2008, 74, 5254–5258. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šimunović, K.; Stefanic, P.; Klančnik, A.; Erega, A.; Mandic Mulec, I.; Smole Možina, S. Bacillus subtilis PS-216 Antagonistic Activities against Campylobacter jejuni NCTC 11168 Are Modulated by Temperature, Oxygen, and Growth Medium. Microorganisms 2022, 10, 289. https://doi.org/10.3390/microorganisms10020289
Šimunović K, Stefanic P, Klančnik A, Erega A, Mandic Mulec I, Smole Možina S. Bacillus subtilis PS-216 Antagonistic Activities against Campylobacter jejuni NCTC 11168 Are Modulated by Temperature, Oxygen, and Growth Medium. Microorganisms. 2022; 10(2):289. https://doi.org/10.3390/microorganisms10020289
Chicago/Turabian StyleŠimunović, Katarina, Polonca Stefanic, Anja Klančnik, Andi Erega, Ines Mandic Mulec, and Sonja Smole Možina. 2022. "Bacillus subtilis PS-216 Antagonistic Activities against Campylobacter jejuni NCTC 11168 Are Modulated by Temperature, Oxygen, and Growth Medium" Microorganisms 10, no. 2: 289. https://doi.org/10.3390/microorganisms10020289
APA StyleŠimunović, K., Stefanic, P., Klančnik, A., Erega, A., Mandic Mulec, I., & Smole Možina, S. (2022). Bacillus subtilis PS-216 Antagonistic Activities against Campylobacter jejuni NCTC 11168 Are Modulated by Temperature, Oxygen, and Growth Medium. Microorganisms, 10(2), 289. https://doi.org/10.3390/microorganisms10020289