
Fluoroquinolone Persistence in Escherichia coli Requires DNA Repair despite Differing between Starving Populations


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Chemicals and Media
2.3. Culture Conditions
2.4. Growth Assay
2.5. Minimum Inhibitory Concentration Assay
2.6. Persister Assay
2.7. Statistical Analysis
3. Results
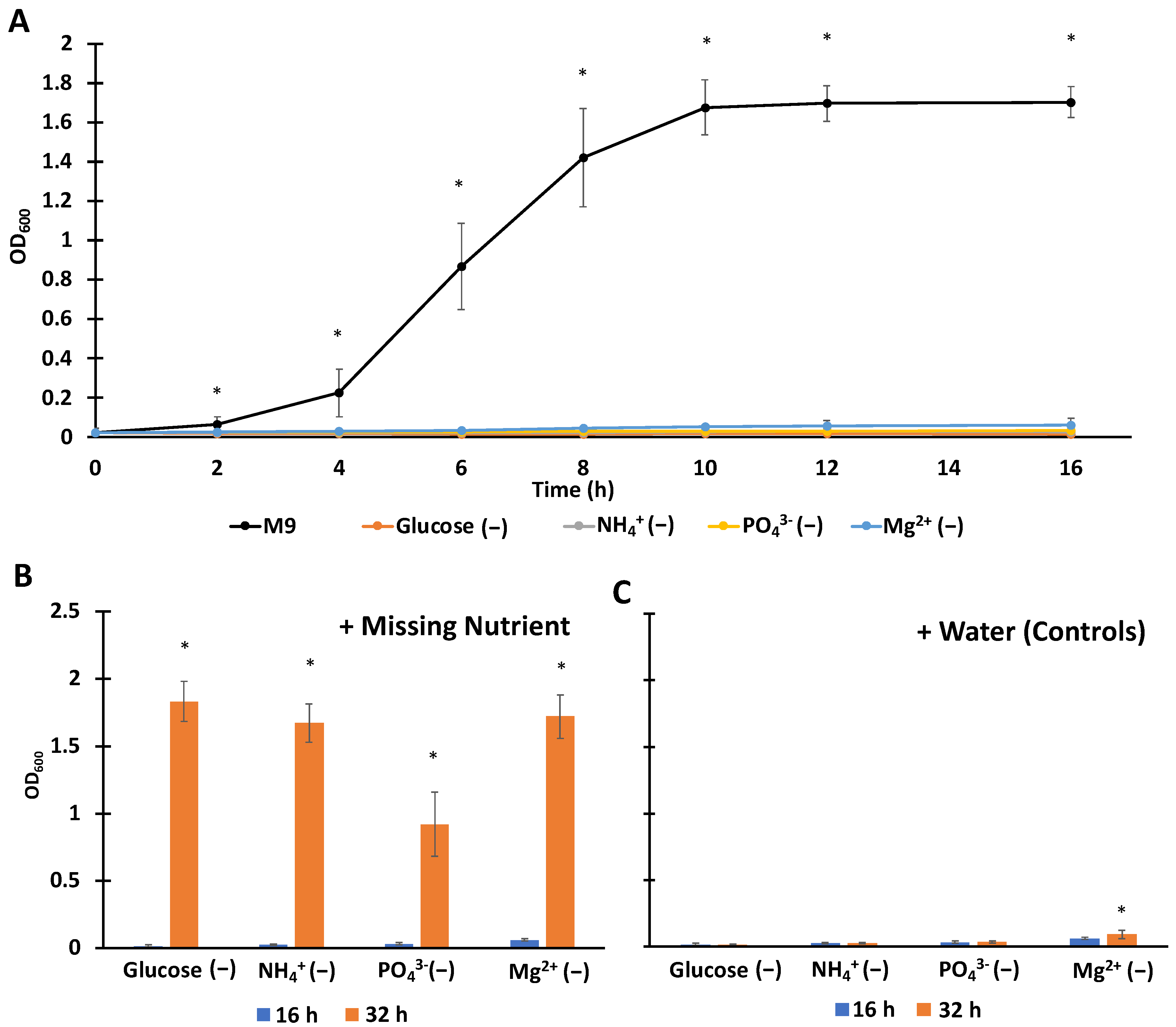
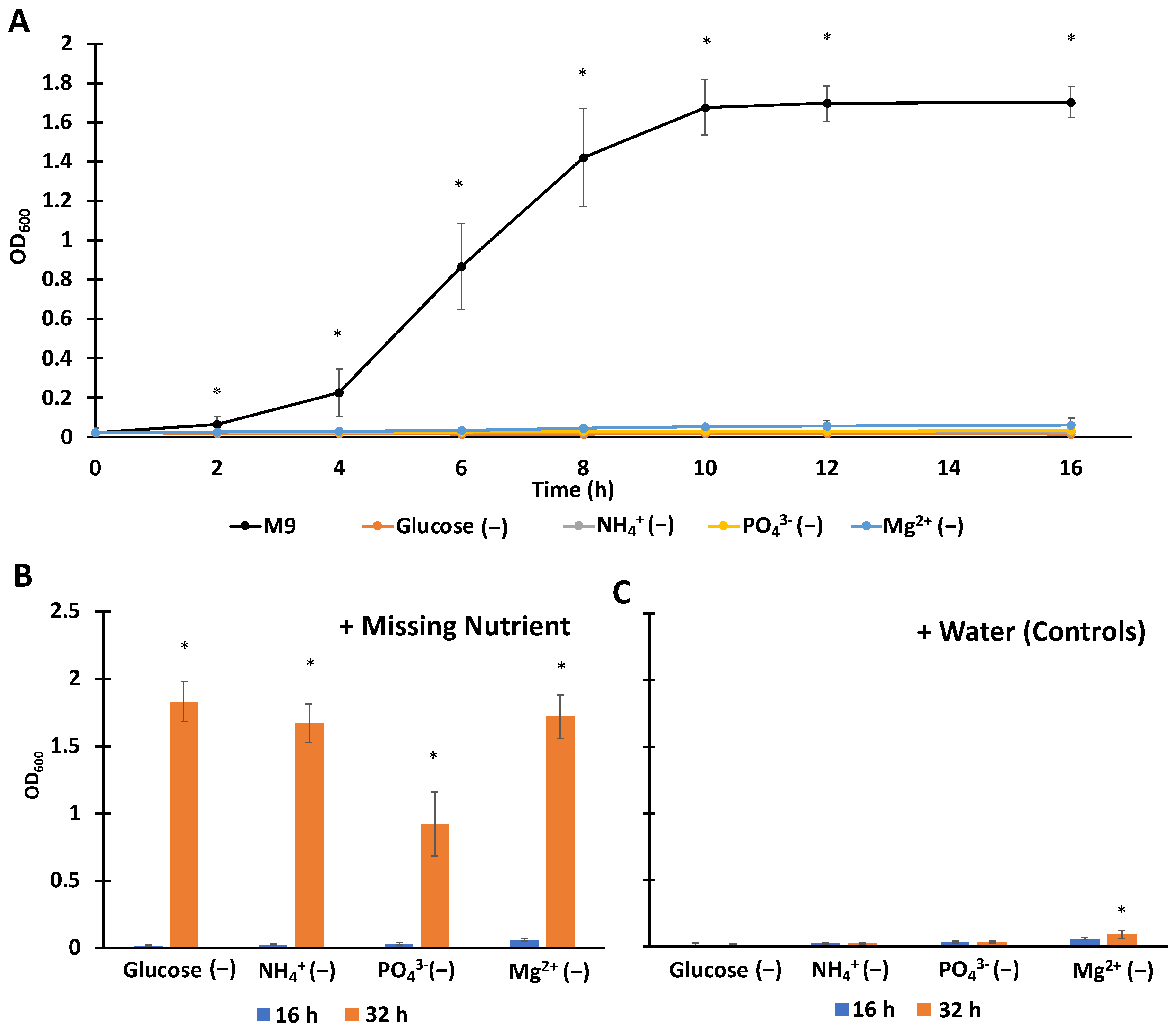
3.1. Establishing Single-Nutrient Starved Culturing Conditions with Complete Media Controls
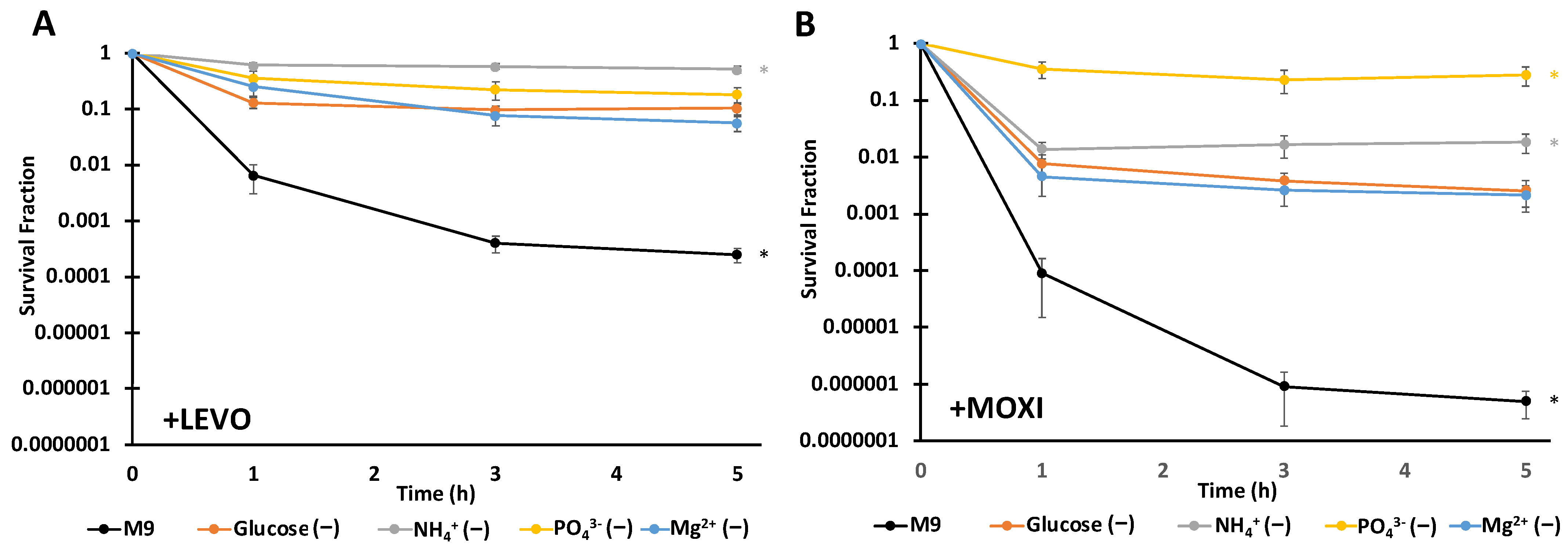
3.2. FQ Persistence Levels Depend on the Type of Starvation and FQ Used
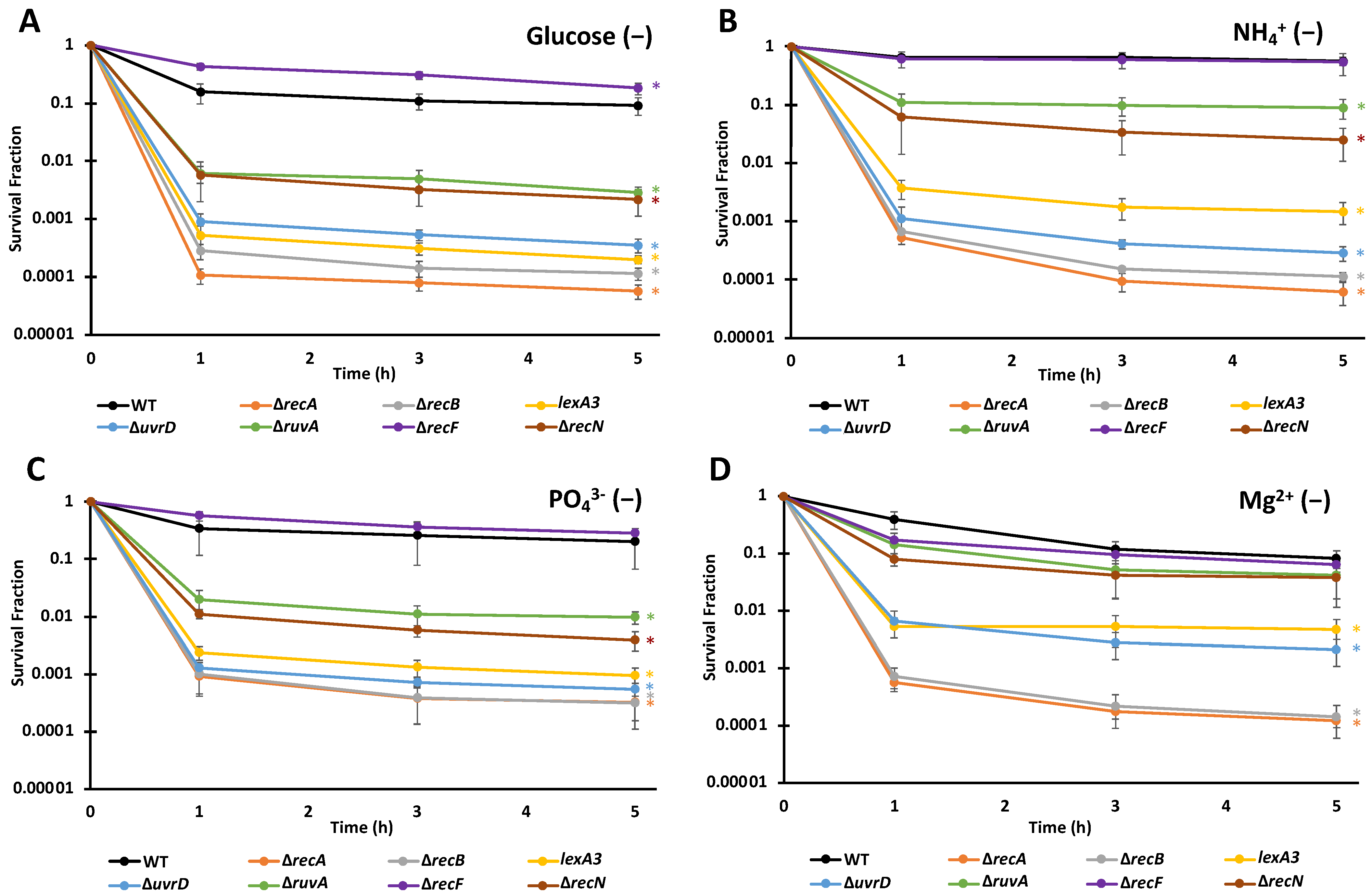
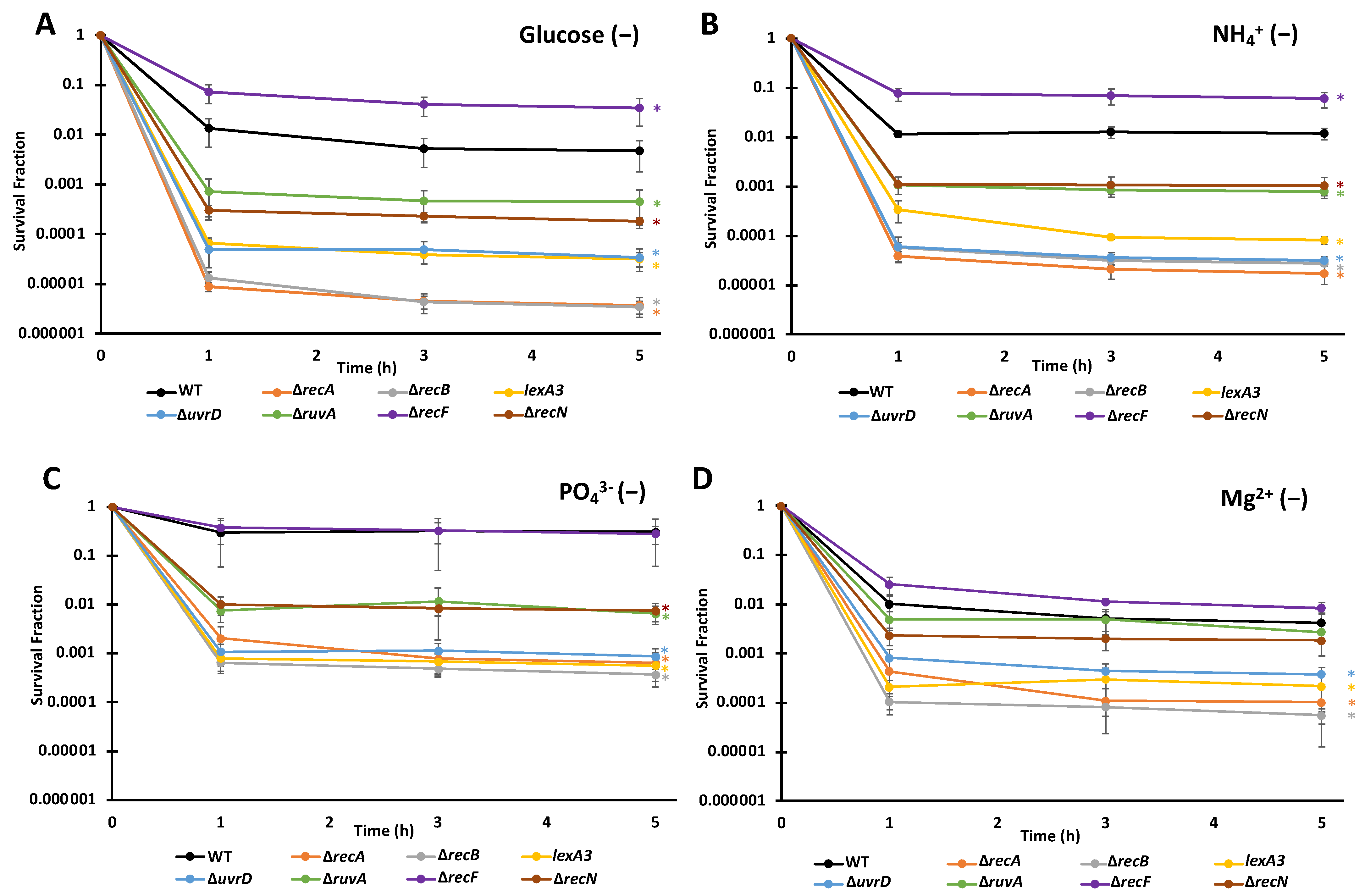
3.3. FQ Persisters Depend on Similar DNA Repair Systems When Deprived of Different Nutrients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alteri, C.J.; Mobley, H.L.T. Escherichia coli Physiology and Metabolism Dictates Adaptation to Diverse Host Microenvironments. Curr. Opin. Microbiol. 2012, 15, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appelberg, R. Macrophage nutriprive antimicrobial mechanisms. J. Leukoc. Biol. 2006, 79, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.A.; Palmer, K.L.; Whiteley, M. Revisiting the host as a growth medium. Nat. Rev. Microbiol. 2008, 6, 657–666. [Google Scholar] [CrossRef] [Green Version]
- Foxman, B. Epidemiology of urinary tract infections: Incidence, morbidity, and economic costs. Am. J. Med. 2002, 113 (Suppl. 1A), 5S–13S. [Google Scholar] [CrossRef]
- Brooks, T.; Keevil, C.W. A simple artificial urine for the growth of urinary pathogens. Lett. Appl. Microbiol. 1997, 24, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, M.; Kasper, L.; Hensel, M.; Hube, B. Metabolic adaptation of intracellular bacteria and fungi to macrophages. Int. J. Med. Microbiol. 2018, 308, 215–227. [Google Scholar] [CrossRef]
- Sivaloganathan, D.M.; Brynildsen, M.P. Phagosome–Bacteria Interactions from the Bottom Up. Annu. Rev. Chem. Biomol. Eng. 2021, 12, 309–331. [Google Scholar] [CrossRef]
- Walters, M.C.; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Stewart, P.S. Contributions of Antibiotic Penetration, Oxygen Limitation, and Low Metabolic Activity to Tolerance of Pseudomonas aeruginosa Biofilms to Ciprofloxacin and Tobramycin. Antimicrob. Agents Chemother. 2003, 47, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Stewart, P.S.; Franklin, M.J. Physiological heterogeneity in biofilms. Nat. Rev. Microbiol. 2008, 6, 199–210. [Google Scholar] [CrossRef]
- Eng, R.H.; Padberg, F.T.; Smith, S.M.; Tan, E.N.; Cherubin, C.E. Bactericidal effects of antibiotics on slowly growing and nongrowing bacteria. Antimicrob. Agents Chemother. 1991, 35, 1824–1828. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Coates, A. Nonmultiplying Bacteria are Profoundly Tolerant to Antibiotics. In Antibiotic Resistance; Coates, A.R.M., Ed.; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2012; pp. 99–119. ISBN 978-3-642-28951-4. [Google Scholar]
- Fung, D.K.C.; Chan, E.W.C.; Chin, M.L.; Chan, R.C.Y. Delineation of a Bacterial Starvation Stress Response Network Which Can Mediate Antibiotic Tolerance Development. Antimicrob. Agents Chemother. 2010, 54, 1082–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontes, M.H.; Groisman, E.A. Slow growth determines nonheritable antibiotic resistance in Salmonella enterica. Sci. Signal. 2019, 12, eaax3938. [Google Scholar] [CrossRef] [PubMed]
- Mok, W.W.K.; Brynildsen, M.P. Nutrient Depletion and Bacterial Persistence. In Persister Cells and Infectious Disease; Lewis, K., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 99–132. ISBN 978-3-030-25241-0. [Google Scholar]
- Brauer, M.J.; Yuan, J.; Bennett, B.D.; Lu, W.; Kimball, E.; Botstein, D.; Rabinowitz, J.D. Conservation of the metabolomic response to starvation across two divergent microbes. Proc. Natl. Acad. Sci. USA 2006, 103, 19302–19307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chubukov, V.; Sauer, U. Environmental Dependence of Stationary-Phase Metabolism in Bacillus subtilis and Escherichia coli. Appl. Environ. Microbiol. 2014, 80, 2901–2909. [Google Scholar] [CrossRef] [Green Version]
- Peterson, C.N.; Mandel, M.J.; Silhavy, T.J. Escherichia coli Starvation Diets: Essential Nutrients Weigh in Distinctly. J. Bacteriol. 2005, 187, 7549–7553. [Google Scholar] [CrossRef] [Green Version]
- Notley-McRobb, L.; Death, A.; Ferenci, T. The relationship between external glucose concentration and cAMP levels inside Escherichia coli: Implications for models of phosphotransferase-mediated regulation of adenylate cyclase. Microbiology (Reading) 1997, 143 Pt 6, 1909–1918. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Kalman, M.; Ikehara, K.; Zemel, S.; Glaser, G.; Cashel, M. Residual guanosine 3′,5′-bispyrophosphate synthetic activity of relA null mutants can be eliminated by spoT null mutations. J. Biol. Chem. 1991, 266, 5980–5990. [Google Scholar] [CrossRef]
- Ballesteros, M.; Fredriksson, A.; Henriksson, J.; Nyström, T. Bacterial senescence: Protein oxidation in non-proliferating cells is dictated by the accuracy of the ribosomes. EMBO J. 2001, 20, 5280–5289. [Google Scholar] [CrossRef] [Green Version]
- Moreau, P.L. Protective role of the RpoE (σE) and Cpx envelope stress responses against gentamicin killing of nongrowing Escherichia coli incubated under aerobic, phosphate starvation conditions. FEMS Microbiol. Lett. 2014, 357, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Aedo, S.J.; Tang, J.; Brynildsen, M.P. Metabolites Potentiate Nitrofurans in Nongrowing Escherichia coli. Antimicrob. Agents Chemother. 2021, 65, e00858-20. [Google Scholar] [CrossRef]
- Völzing, K.G.; Brynildsen, M.P. Stationary-Phase Persisters to Ofloxacin Sustain DNA Damage and Require Repair Systems Only during Recovery. mBio 2015, 6, e00731-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mok, W.W.K.; Brynildsen, M.P. Timing of DNA damage responses impacts persistence to fluoroquinolones. Proc. Natl. Acad. Sci. USA 2018, 115, E6301–E6309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keren, I.; Kaldalu, N.; Spoering, A.; Wang, Y.; Lewis, K. Persister cells and tolerance to antimicrobials. FEMS Microbiol. Lett. 2004, 230, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Drlica, K.; Zhao, X. DNA gyrase, topoisomerase IV, and the 4-quinolones. Microbiol Mol. Biol. Rev. 1997, 61, 377–392. [Google Scholar]
- Berger, J.M.; Gamblin, S.J.; Harrison, S.C.; Wang, J.C. Structure and mechanism of DNA topoisomerase II. Nature 1996, 379, 225–232. [Google Scholar] [CrossRef]
- Sifaoui, F.; Lamour, V.; Varon, E.; Moras, D.; Gutmann, L. ATP-Bound Conformation of Topoisomerase IV: A Possible Target for Quinolones in Streptococcus pneumoniae. J. Bacteriol. 2003, 185, 6137–6146. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.V.; Maxwell, A. Identification of a residue involved in transition-state stabilization in the ATPase reaction of DNA gyrase. Biochemistry 1998, 37, 9658–9667. [Google Scholar] [CrossRef]
- Balaban, N.Q.; Merrin, J.; Chait, R.; Kowalik, L.; Leibler, S. Bacterial persistence as a phenotypic switch. Science 2004, 305, 1622–1625. [Google Scholar] [CrossRef] [Green Version]
- Jõers, A.; Kaldalu, N.; Tenson, T. The Frequency of Persisters in Escherichia coli Reflects the Kinetics of Awakening from Dormancy. J. Bacteriol. 2010, 192, 3379–3384. [Google Scholar] [CrossRef] [Green Version]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Byrd, B.A.; Zenick, B.; Rocha-Granados, M.C.; Englander, H.E.; Hare, P.J.; LaGree, T.J.; DeMarco, A.M.; Mok, W.W.K. The AcrAB-TolC Efflux Pump Impacts Persistence and Resistance Development in Stationary-Phase Escherichia coli following Delafloxacin Treatment. Antimicrob. Agents Chemother. 2021, 65, e0028121. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.C.; Mok, W.W.K.; Murawski, A.M.; Brynildsen, M.P. Enhanced antibiotic resistance development from fluoroquinolone persisters after a single exposure to antibiotic. Nat. Commun. 2019, 10, 1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murawski, A.M.; Brynildsen, M.P. Ploidy is an important determinant of fluoroquinolone persister survival. Curr. Biol. 2021, 31, 2039–2050.e7. [Google Scholar] [CrossRef] [PubMed]
- Henry, T.C.; Brynildsen, M.P. Development of Persister-FACSeq: A method to massively parallelize quantification of persister physiology and its heterogeneity. Sci. Rep. 2016, 6, 25100. [Google Scholar] [CrossRef]
- Theodore, A.; Lewis, K.; Vulic, M. Tolerance of Escherichia coli to fluoroquinolone antibiotics depends on specific components of the SOS response pathway. Genetics 2013, 195, 1265–1276. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S.; Lewis, K.; Vulić, M. Role of Global Regulators and Nucleotide Metabolism in Antibiotic Tolerance in Escherichia coli. Antimicrob. Agents Chemother. 2008, 52, 2718–2726. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, Y. PhoU is a persistence switch involved in persister formation and tolerance to multiple antibiotics and stresses in Escherichia coli. Antimicrob. Agents Chemother. 2007, 51, 2092–2099. [Google Scholar] [CrossRef] [Green Version]
- Sezonov, G.; Joseleau-Petit, D.; D’Ari, R. Escherichia coli Physiology in Luria-Bertani Broth. J. Bacteriol. 2007, 189, 8746–8749. [Google Scholar] [CrossRef] [Green Version]
- Roca, A.I.; Cox, M.M. RecA Protein: Structure, Function, and Role in Recombinational DNA Repair. In Progress in Nucleic Acid Research and Molecular Biology; Cohn, W.E., Moldave, K., Eds.; Academic Press: Cambridge, MA, USA, 1997; Volume 56, pp. 129–223. [Google Scholar]
- Chaudhury, A.M.; Smith, G.R. A new class of Escherichia coli recBC mutants: Implications for the role of RecBC enzyme in homologous recombination. Proc. Natl. Acad. Sci. USA 1984, 81, 7850–7854. [Google Scholar] [CrossRef] [Green Version]
- Simmons, L.A.; Foti, J.J.; Cohen, S.E.; Walker, G.C. The SOS Regulatory Network. Ecosal Plus 2008, 2008. [Google Scholar] [CrossRef] [Green Version]
- Moolenaar, G.F.; Moorman, C.; Goosen, N. Role of the Escherichia coli Nucleotide Excision Repair Proteins in DNA Replication. J. Bacteriol. 2000, 182, 5706–5714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, M.C.; Jordan, J.R.; Matson, S.W. Evidence for a physical interaction between the Escherichia coli methyl-directed mismatch repair proteins MutL and UvrD. EMBO J. 1998, 17, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.N.; Courcelle, C.T.; Courcelle, J. UvrD Participation in Nucleotide Excision Repair Is Required for the Recovery of DNA Synthesis following UV-Induced Damage in Escherichia coli. J. Nucleic Acids 2012, 2012, e271453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, H.; Takahagi, M.; Nakata, A.; Shinagawa, H. Escherichia coli RuvA and RuvB proteins specifically interact with Holliday junctions and promote branch migration. Genes Dev. 1992, 6, 2214–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uranga, L.A.; Reyes, E.D.; Patidar, P.L.; Redman, L.N.; Lusetti, S.L. The Cohesin-like RecN Protein Stimulates RecA-Mediated Recombinational Repair of DNA Double-Strand Breaks. Nat. Comm. 2017, 8, 15282. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [Green Version]
- Thomason, L.C.; Costantino, N.; Court, D.L. E. coli genome manipulation by P1 transduction. Curr. Protoc. Mol. Biol. 2007. [Google Scholar] [CrossRef]
- Little, J.W.; Harper, J.E. Identification of the lexA gene product of Escherichia coli K-12. Proc. Natl. Acad. Sci. USA 1979, 76, 6147–6151. [Google Scholar] [CrossRef] [Green Version]
- Kühnau, S.; Reyes, M.; Sievertsen, A.; Shuman, H.A.; Boos, W. The activities of the Escherichia coli MalK protein in maltose transport, regulation, and inducer exclusion can be separated by mutations. J. Bacteriol. 1991, 173, 2180–2186. [Google Scholar] [CrossRef] [Green Version]
- Mount, D.W. A mutant of Escherichia coli showing constitutive expression of the lysogenic induction and error-prone DNA repair pathways. Proc. Natl. Acad. Sci. USA 1977, 74, 300–304. [Google Scholar] [CrossRef] [Green Version]
- Goormaghtigh, F.; Van Melderen, L. Single-cell imaging and characterization of Escherichia coli persister cells to ofloxacin in exponential cultures. Sci. Adv. 2019, 5, eaav9462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, D.C.; Jacoby, G.A. Topoisomerase Inhibitors: Fluoroquinolone Mechanisms of Action and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.L.; Baranowski, J.; Pernet, A.G. Mechanism of inhibition of DNA gyrase by quinolone antibacterials: Specificity and cooperativity of drug binding to DNA. Biochemistry 1989, 28, 3879–3885. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.F.; Wang, J.C. Supercoiling of the DNA template during transcription. Proc. Natl. Acad. Sci. USA 1987, 84, 7024–7027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorman, C.J. Regulation of Transcription in Bacteria by DNA Supercoiling. In Bacterial Physiology: A Molecular Approach; El-Sharoud, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 155–178. ISBN 978-3-540-74921-9. [Google Scholar]
- Kato, J.; Nishimura, Y.; Imamura, R.; Niki, H.; Hiraga, S.; Suzuki, H. New topoisomerase essential for chromosome segregation in E. coli. Cell 1990, 63, 393–404. [Google Scholar] [CrossRef]
- Gefen, O.; Fridman, O.; Ronin, I.; Balaban, N.Q. Direct observation of single stationary-phase bacteria reveals a surprisingly long period of constant protein production activity. Proc. Natl. Acad. Sci. USA 2014, 111, 556–561. [Google Scholar] [CrossRef] [Green Version]
- Kolter, R.; Siegele, D.A.; Tormo, A. The Stationary Phase of the Bacterial Life Cycle. Annu. Rev. Microbiol. 1993, 47, 855–874. [Google Scholar] [CrossRef]
- Brown, D.R. Nitrogen Starvation Induces Persister Cell Formation in Escherichia coli. J. Bacteriol. 2019, 201, e00622-18. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Wang, X.; Meng, L.; Zhu, M.; Wu, J.; Xu, Y.; Zhang, Y.; Zhang, W. Magnesium Links Starvation-Mediated Antibiotic Persistence to ATP. mSphere 2020, 5, e00862-19. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Chan, E.W.C.; Wan, Y.; Wong, M.H.-Y.; Chen, S. Active maintenance of proton motive force mediates starvation-induced bacterial antibiotic tolerance in Escherichia coli. Commun. Biol. 2021, 4, 1068. [Google Scholar] [CrossRef]
- Lemma, A.S.; Brynildsen, M.P. Toxin Induction or Inhibition of Transcription or Translation Posttreatment Increases Persistence to Fluoroquinolones. mBio 2021, 12, e0198321. [Google Scholar] [CrossRef] [PubMed]
- Kowalczykowski, S.C. Initiation of genetic recombination and recombination-dependent replication. Trends Biochem. Sci. 2000, 25, 156–165. [Google Scholar] [CrossRef]
- Dörr, T.; Vulić, M.; Lewis, K. Ciprofloxacin Causes Persister Formation by Inducing the TisB toxin in Escherichia coli. PLoS Biol. 2010, 8, e1000317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dörr, T.; Lewis, K.; Vulić, M. SOS Response Induces Persistence to Fluoroquinolones in Escherichia coli. PLoS Genet. 2009, 5, e1000760. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Glucose (−) | NH4 + (-) | PO4 3− (−) | Mg 2+ (−) | M9 |
|---|---|---|---|---|---|
| WT | 9.26 × 10−2 (+/− 3.09 × 10−2) | 5.58 × 10−1 (+/− 8.82 × 10−2) | 2.02 × 10−1 (+/− 1.36 × 10−1) | 8.28 × 10−2 (+/− 2.77 × 10−2) | 2.50 × 10−4 (+/− 7.10 × 10−5) |
| ∆recA | 5.75 × 10−5 (+/− 1.63 × 10−5) | 6.23 × 10−5 (+/− 2.61 × 10−5) | 3.25 × 10−4 (+/− 1.69 × 10−4) | 1.22 × 10−4 (+/− 2.74 × 10−5) | |
| ∆recB | 1.16 × 10−4 (+/− 2.84 × 10−5) | 1.13 × 10−4 (+/− 1.75 × 10−5) | 3.08 × 10−4 (+/− 2.00 × 10−4) | 1.44 × 10−4 (+/− 8.21 × 10−5) | |
| lexA3 | 2.00 × 10−4 (+/− 2.92 × 10−5) | 1.48 × 10−3 (+/− 6.00 × 10−4) | 9.35 × 10−4 (+/− 3.45 × 10−4) | 4.71 × 10−3 (+/− 2.41 × 10−3) | |
| ∆uvrD | 3.53 × 10−4 (+/− 9.52 × 10−5) | 2.86 × 10−4 (+/− 8.02 × 10−5) | 5.48 × 10−4 (+/− 1.37 × 10−4) | 2.14 × 10−3 (+/− 1.04 × 10−3) | |
| ∆ruvA | 2.85 × 10−3 (+/− 7.19 × 10−4) | 8.99 × 10−2 (+/− 3.30 × 10−2) | 9.77 × 10−3 (+/− 2.37 × 10−3) | 4.17 × 10−2 (+/− 3.03 × 10−2) | |
| ∆recF | 1.81 × 10−1 (+/− 4.20 × 10−2) | 5.42 × 10−1 (+/− 2.22 × 10−1) | 2.81 × 10−1 (+/− 6.08 × 10−2) | 6.36 × 10−2 (+/− 1.65 × 10−2) | |
| ∆recN | 2.16 × 10−3 (+/− 1.04 × 10−3) | 2.54 × 10−2 (+/− 1.46 × 10−2) | 3.92 × 10−3 (+/− 1.42 × 10−3) | 3.82 × 10−2 (+/− 2.18 × 10−2) |
| Strain | Glucose (−) | NH4 + (−) | PO4 3− (−) | Mg 2+ (−) | M9 |
|---|---|---|---|---|---|
| WT | 4.76 × 10−3 (+/− 2.98 × 10−3) | 1.19 × 10−2 (+/− 3.11 × 10−3) | 3.07 × 10−1 (+/− 2.47 × 10−1) | 4.28 × 10−3 (+/− 2.49 × 10−3) | 5.00 × 10−7 (+/− 2.51 × 10−7) |
| ∆recA | 3.71 × 10−6 (+/− 1.56 × 10−6) | 1.73 × 10−5 (+/− 6.92 × 10−6) | 6.46 × 10−4 (+/− 3.72 × 10−4) | 1.03 × 10−4 (+/− 9.00 × 10−5) | |
| ∆recB | 3.48 × 10−6 (+/− 1.05 × 10−6) | 2.75 × 10−5 (+/− 9.98 × 10−6) | 3.70 × 10−4 (+/− 1.63 × 10−4) | 5.54 × 10−5 (+/− 1.93 × 10−5) | |
| lexA3 | 3.20 × 10−5 (+/− 1.03 × 10−5) | 8.09 × 10−5 (+/− 1.43 × 10−5) | 5.65 × 10−4 (+/− 2.92 × 10−4) | 2.19 × 10−4 (+/− 1.53 × 10−4) | |
| ∆uvrD | 3.42 × 10−5 (+/− 1.65 × 10−5) | 3.14 × 10−5 (+/− 6.45 × 10−6) | 8.72 × 10−4 (+/− 3.99 × 10−4) | 3.80 × 10−4 (+/− 1.47 × 10−4) | |
| ∆ruvA | 4.54 × 10−4 (+/− 3.24 × 10−4) | 7.89 × 10−4 (+/−1.53 × 10−4) | 6.55 × 10−3 (+/− 2.60 × 10−3) | 2.74 × 10−3 (+/− 1.04 × 10−3) | |
| ∆recF | 3.46 × 10−2 (+/− 1.96 × 10−2) | 5.96 × 10−2 (+/− 2.02 × 10−2) | 2.82 × 10−1 (+/− 1.14 × 10−1) | 8.47 × 10−3 (+/− 2.20 × 10−3) | |
| ∆recN | 1.83 × 10−4 (+/− 2.94 × 10−5) | 1.03 × 10−3 (+/− 4.66 × 10−4) | 7.57 × 10−3 (+/− 3.02 × 10−3) | 1.84 × 10−3 (+/− 9.36 × 10−4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemma, A.S.; Soto-Echevarria, N.; Brynildsen, M.P. Fluoroquinolone Persistence in Escherichia coli Requires DNA Repair despite Differing between Starving Populations. Microorganisms 2022, 10, 286. https://doi.org/10.3390/microorganisms10020286
Lemma AS, Soto-Echevarria N, Brynildsen MP. Fluoroquinolone Persistence in Escherichia coli Requires DNA Repair despite Differing between Starving Populations. Microorganisms. 2022; 10(2):286. https://doi.org/10.3390/microorganisms10020286
Chicago/Turabian StyleLemma, Annabel S., Nashaly Soto-Echevarria, and Mark P. Brynildsen. 2022. "Fluoroquinolone Persistence in Escherichia coli Requires DNA Repair despite Differing between Starving Populations" Microorganisms 10, no. 2: 286. https://doi.org/10.3390/microorganisms10020286
APA StyleLemma, A. S., Soto-Echevarria, N., & Brynildsen, M. P. (2022). Fluoroquinolone Persistence in Escherichia coli Requires DNA Repair despite Differing between Starving Populations. Microorganisms, 10(2), 286. https://doi.org/10.3390/microorganisms10020286