
A Novel Genospecies of Borrelia burgdorferi Sensu Lato Associated with Cricetid Rodents in Brazil


Abstract
:1. Introduction
2. Materials and Methods
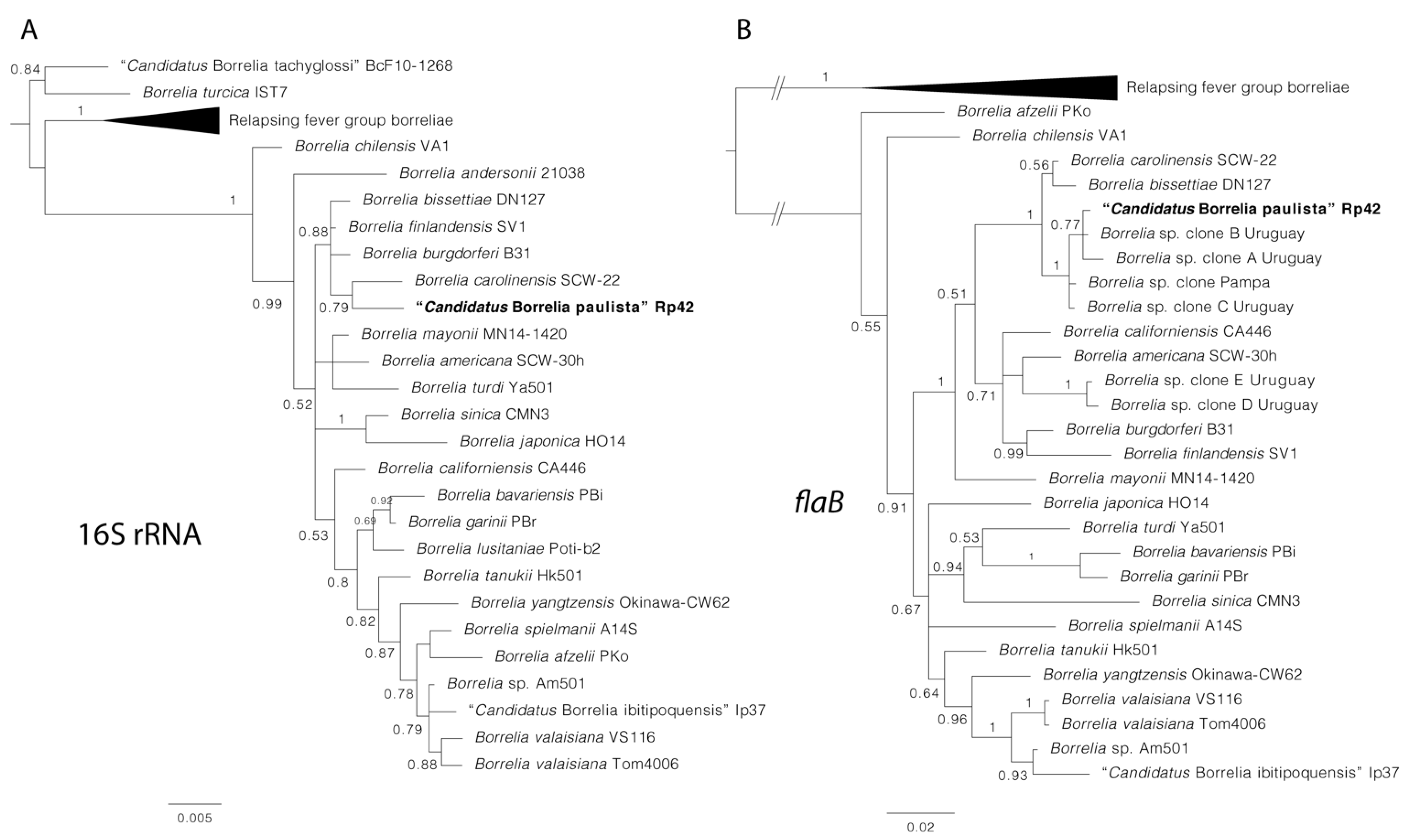
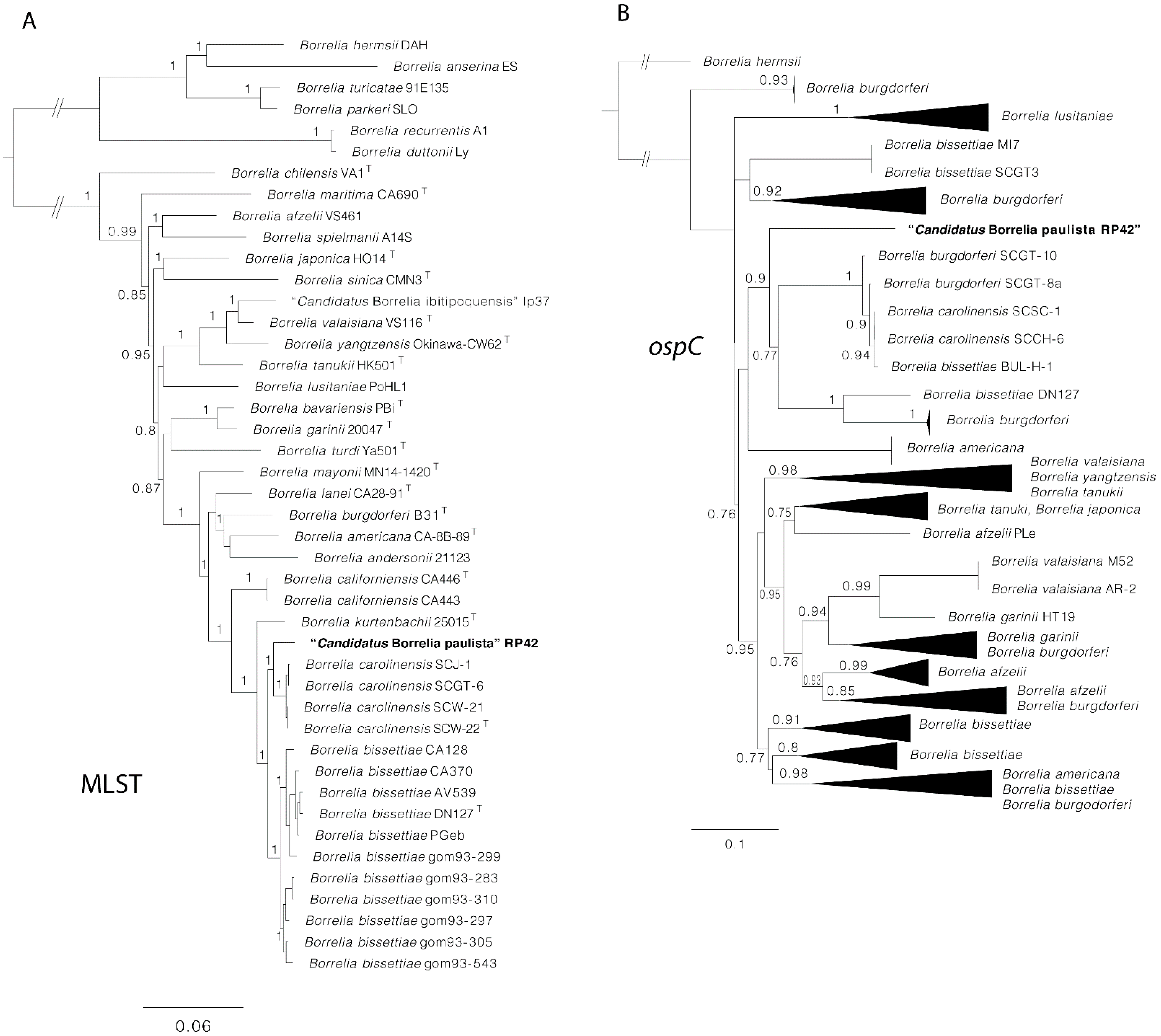
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H. Borrelia burgdorferi sensu lato complex with respect to public health. Ticks Tick Borne Dis. 2011, 2, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Margos, G.; Chu, C.-Y.; Takano, A.; Jiang, B.-G.; Liu, W.; Kurtenbach, K.; Masuzawa, T.; Fingerle, V.; Cao, W.C.; Kawabata, H. Borrelia yangtzensis sp. nov., a rodent-associated species in Asia, is related to Borrelia valaisiana. Int. J. Syst. Evol. 2015, 65, 3836–3840. [Google Scholar] [CrossRef]
- Muñoz-Leal, S.; Ramirez, D.G.; Luz, H.R.; Faccini, J.L.H.; Labruna, M.B. “Candidatus Borrelia ibitipoquensis,” a Borrelia valaisiana–related genospecies characterized from Ixodes paranaensis in Brazil. Microb. Ecol. 2020, 80, 682–689. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Strle, F.; Wormser, G.P.; Hu, L.T.; Branda, J.A.; Hovius, J.W.R.; Li, X.; Mead, P.S. Lyme borreliosis. Nat. Rev. Dis. Primers 2016, 2, 16090. [Google Scholar] [CrossRef]
- Pritt, B.S.; Respicio-Kingry, L.B.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; Bjork, J.; Liu, G.; Kingry, L.C.; Mead, P.S.; Neitzel, D.F.; et al. Borrelia mayonii sp. nov., a member of the Borrelia burgdorferi sensu lato complex, detected in patients and ticks in the upper midwestern United States. Int. J. Syst. Evol. 2016, 66, 4878–4880. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Gofton, A.; Wibberg, D.; Dangel, A.; Marosevic, D.; Loh, S.-M.; Oskam, C.; Fingerle, V. The genus Borrelia reloaded. PLoS ONE 2018, 13, e0208432. [Google Scholar] [CrossRef]
- Miziara, C.S.M.G.; Serrano, V.A.G.; Yoshinari, N. Passage of Borrelia burgdorferi through diverse Ixodid hard ticks causes distinct diseases: Lyme borreliosis and Baggio-Yoshinari syndrome. Clinics 2018, 73, 1–4. [Google Scholar] [CrossRef]
- Oliveira, S.V.; Faccini-martínez, Á.A.; Cerutti, C.J. Lack of serological evidence for Lyme-like borreliosis in Brazil. Travel Med. Infect. Dis. 2018, 26, 62–63. [Google Scholar] [CrossRef]
- Durden, L.A. Taxonomy, hosts associations, life cycles and vectorial importance of ticks parasitizing small mammals. In Micromammals and Macroparasites: From Evolutionary Ecology to Management; Morand, S., Krasnov, B.R., Poulin, R., Eds.; Springer: Tokyo, Japan, 2006; pp. 1–647. [Google Scholar] [CrossRef]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H. Borrelia carolinensis sp. nov., a novel species of the Borrelia burgdorferi sensu lato complex isolated from rodents and a tick from the south-eastern USA. Int. J. Syst. Evol. 2011, 61, 381–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margos, G.; Lane, R.S.; Fedorova, N.; Koloczek, J.; Piesman, J.; Hojgaard, A.; Sing, A.; Fingerle, V. Borrelia bissettiae sp. nov. and Borrelia californiensis sp. nov. prevail in diverse enzootic transmission cycles. Int. J. Syst. Evol. 2016, 66, 1447–1452. [Google Scholar] [CrossRef]
- Tilly, K.; Krum, J.G.; Bestor, A.; Jewett, M.W.; Grimm, D.; Bueschel, D.; Byram, R.; Dorward, D.; VanRaden, M.J.; Stewart, P.; et al. Borrelia burgdorferi OspC protein required exclusively in a crucial early stage of mammalian infection. Infect. Immun. 2006, 74, 3554–3564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbour, A.G.; Travinsky, B. Evolution and distribution of the ospC gene, a transferable serotype determinant of Borrelia burgdorferi. mBio 2010, 1, e00153-10. [Google Scholar] [CrossRef] [Green Version]
- Sebastian, P.S.; Bottero, M.N.S.; Carvalho, L.; Mackenstedt, U.; Lareschi, M.; Venzal, J.M.; Nava, S. Borrelia burgdorferi sensu lato in Ixodes cf. neuquenensis and Ixodes sigelos ticks from the Patagonian region of Argentina. Acta Trop. 2016, 162, 218–221. [Google Scholar] [CrossRef]
- Ivanova, L.B.; Tomova, A.; González-Acuña, D.; Murúa, R.; Moreno, C.X.; Hernández, C.; Cabello, J.; Cabello, C.; Daniels, T.J.; Godfrey, H.P.; et al. Borrelia chilensis, a new member of the Borrelia burgdorferi sensu lato complex that extends the range of this genospecies in the Southern Hemisphere. Environ. Microbiol. 2014, 16, 1069–1080. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Leal, S.; Lopes, M.G.; Marcili, A.; Martins, T.F.; González-Acuña, D.; Labruna, M.B. Anaplasmataceae, Borrelia and Hepatozoon agents in ticks (Acari: Argasidae, Ixodidae) from Chile. Acta Trop. 2019, 192, 91–103. [Google Scholar] [CrossRef]
- Thomas, R.S.; Santodomingo, A.M.S.; Muñoz-Leal, S.; Silva-de la Fuente, M.C.; Llanos-Soto, S.; Salas, L.M.; González-Acuña, D. Rodents as potential reservoirs for Borrelia spp. in northern Chile. Revista Brasileira de Parasitologia Veterinária 2020, 29, 1–10. [Google Scholar] [CrossRef]
- Dall’Agnol, B.; Michel, T.; Weck, B.; Souza, U.A.; Webster, A.; Leal, B.F.; Klafke, G.M.; Martins, J.R.; Ott, R.; Venzal, J.M.; et al. Borrelia burgdorferi sensu lato in Ixodes longiscutatus ticks from Brazilian Pampa. Ticks Tick Borne Dis. 2017, 8, 928–932. [Google Scholar] [CrossRef]
- Serpa, M.C.A.; Luz, H.R.; Costa, F.B.; Weck, B.C.; Benatti, H.R.; Martins, T.F.; Correa, L.S.; Ramirez, D.G.; Rocha, V.; Dias, T.C.; et al. Small mammals, ticks and rickettsiae in natural and human-modified landscapes: Diversity and occurrence of Brazilian spotted fever in Brazil. Ticks Tick Borne Dis. 2021, 12, 101805. [Google Scholar] [CrossRef] [PubMed]
- Reis, N.R.; Peracchi, A.L.; Pedro, W.A.; Lima, I.P. Mamíferos do Brasil; SEMA: Londrina, Brazil, 2006; pp. 1–437. [Google Scholar]
- Bonvicino, C.R. Guia dos Roedores do Brasil, com Chaves para Gêneros Baseados em Características Externas; Centro Pan-Americano de Febre Aftosa—OPAS/OMS: Rio de Janeiro, Brazil, 2008; pp. 1–120. [Google Scholar]
- Steuber, S.; Abdel-Rady, A.; Clausen, P.H. PCR-RFLP analysis: A promising technique for host species identification of blood meals from tsetse flies (Diptera: Glossinidae). Parasitol. Res. 2005, 97, 247–254. [Google Scholar] [CrossRef]
- Parola, P.; Diatta, G.; Socolovschi, C.; Mediannikov, O.; Tall, A.; Bassene, H.; Trape, J.F.; Raoult, D. Tick-Borne Relapsing Fever Borreliosis, Rural Senegal. Emerg Infect. Dis. 2011, 17, 883–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwan, T.G.; Raffel, S.J.; Schrumpf, M.E.; Policastro, P.F.; Rawlings, J.A.; Lane, R.S.; Breitschwerdt, E.B.; Porcella, S.F. Phylogenetic Analysis of the Spirochetes Borrelia parkeri and Borrelia turicatae and the Potential for Tick-Borne Relapsing Fever in Florida. J. Clin. Microbiol. 2005, 43, 3851–3859. [Google Scholar] [CrossRef] [Green Version]
- Stromdahl, E.Y.; Williamson, P.C.; Kollars, T.M.J.; Evans, S.R.; Barry, R.K.; Vince, M.A.; Dobbs, N.A. DNA evidence of Borrelia lonestari in Amblyomma americanum (Acari: Ixodidae) removed from humans. J. Clin. Microbiol. 2003, 41, 5557–5562. [Google Scholar] [CrossRef] [Green Version]
- Bunikis, J.; Garpmo, U.F.; Tsao, J.; Berglund, J.; Fish, D.; Barbour, A. Sequence typing reveals extensive strain diversity of the Lyme borreliosis agents Borrelia burgdorferi in North America and Borrelia afzelii in Europe. Microbiology 2004, 150, 1741–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margos, G.; Gatewood, A.G.; Aanensen, D.M.; Hanincova, K.; Terekhova, D.; Vollmer, S.A.; Cornet, M.; Piesman, J.; Donaghy, M.; Bormane, A.; et al. MLST of housekeeping genes captures geographic population structure and suggests a European origin of Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA 2008, 105, 8730–8735. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Leal, S.; Faccini-Martínez, Á.A.; Costa, F.B.; Marcili, A.; Mesquita, E.T.K.C.; Marques, E.P.; Labruna, M.B. Isolation and molecular characterization of a relapsing fever Borrelia recovered from Ornithodoros rudis in Brazil. Ticks Tick Borne Dis. 2018, 9, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kima, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. Spec. Publ. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Patton, J.L.; Pardiñas, U.F.J.; D’Elía, G. Mammals of South America; University of Chicago Press: Chicago, IL, USA; London, UK, 2015; pp. 1–1384. [Google Scholar] [CrossRef]
- Foley, J.; Ott-Conn, C.; Wort, J.; Poulsen, A.; Clifford, D. An Ixodes minor and Borrelia carolinensis enzootic cycle involving a critically endangered Mojave Desert rodent. Ecol. Evol. 2014, 4, 576–581. [Google Scholar] [CrossRef]
- Solís-Hernández, A.; Rodríguez-Vivas, R.I.; Esteve-Gassent, M.D.; Villegas-Pérez, S.L. Prevalencia de Borrelia burgdorferi sensu lato en roedores sinantrópicos de dos comunidades rurales de Yucatán, México. Biomedica 2016, 36, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Wolcott, K.A.; Margos, G.; Fingerle, V.; Becker, N. Host association of Borrelia burgdorferi sensu lato: A review. Ticks Tick Borne Dis. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Weksler, M.; Bonvicino, C.R. Taxonomy of pigmy rice rats genus Oligoryzomys Bangs, 1900 (Rodentia, Sigmodontinae) of the Brazilian Cerrado, with the description of two new species. Arquivos do Museu Nacional 2005, 63, 113–130. [Google Scholar]
- Weksler, M.; Lemos, E.; D’Andrea, P.; Bonvicino, C.R. The Taxonomic Status of Oligoryzomys mattogrossae (Allen 1916) (Rodentia: Cricetidae: Sigmodontinae), Reservoir of Anajatuba Hantavirus. Am. Mus. Novit. 2017, 3880, 1–32. [Google Scholar] [CrossRef]
- Barbieri, A.M.; Venzal, J.M.; Marcili, A.; Almeida, A.P.; González, E.M.; Labruna, M.B. Borrelia burgdorferi sensu lato Infecting Ticks of the Ixodes ricinus Complex in Uruguay: First Report for the Southern Hemisphere. Vector-Borne Zoonotic Dis. 2013, 13, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Labruna, M.B.; Onofrio, V.C.; Barros-Battesti, D.M.; Gianizella, S.L.; Venzal, J.M.; Guglielmone, A.A. Synonymy of Ixodes aragaoi with Ixodes fuscipes, and reinstatement of Ixodes spinosus (Acari: Ixodidae). Ticks Tick Borne Dis. 2020, 11, 101349. [Google Scholar] [CrossRef] [PubMed]
- Nava, S.; Venzal, J.M.; González-Acuña, D.; Martins, T.F.; Guglielmone, A.A. Ticks of the Southern Cone of America: Diagnosis, Distribution and Hosts with Taxonomy, Ecology and Sanitary Importance; Elsevier: Cambridge, MA, USA, 2017; 348p. [Google Scholar]
- Guglielmone, A.; Robbins, R. Tick Species Found Feeding on Humans: A Global Overview. In Hard Ticks (Acari: Ixodida: Ixodidae) Parasitizing Humans; Springer: Cham, Switzerland, 2018; pp. 1–205. [Google Scholar] [CrossRef]
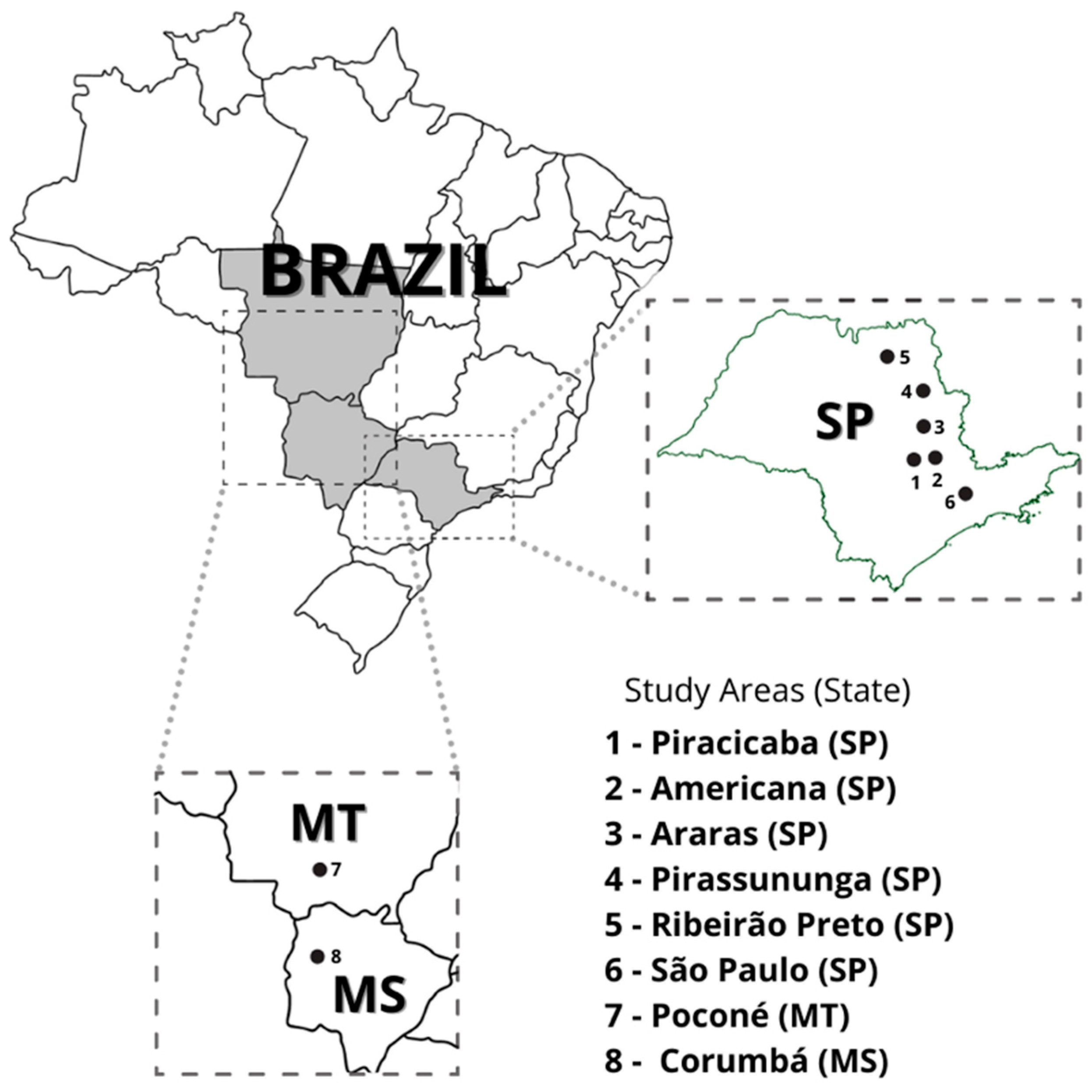
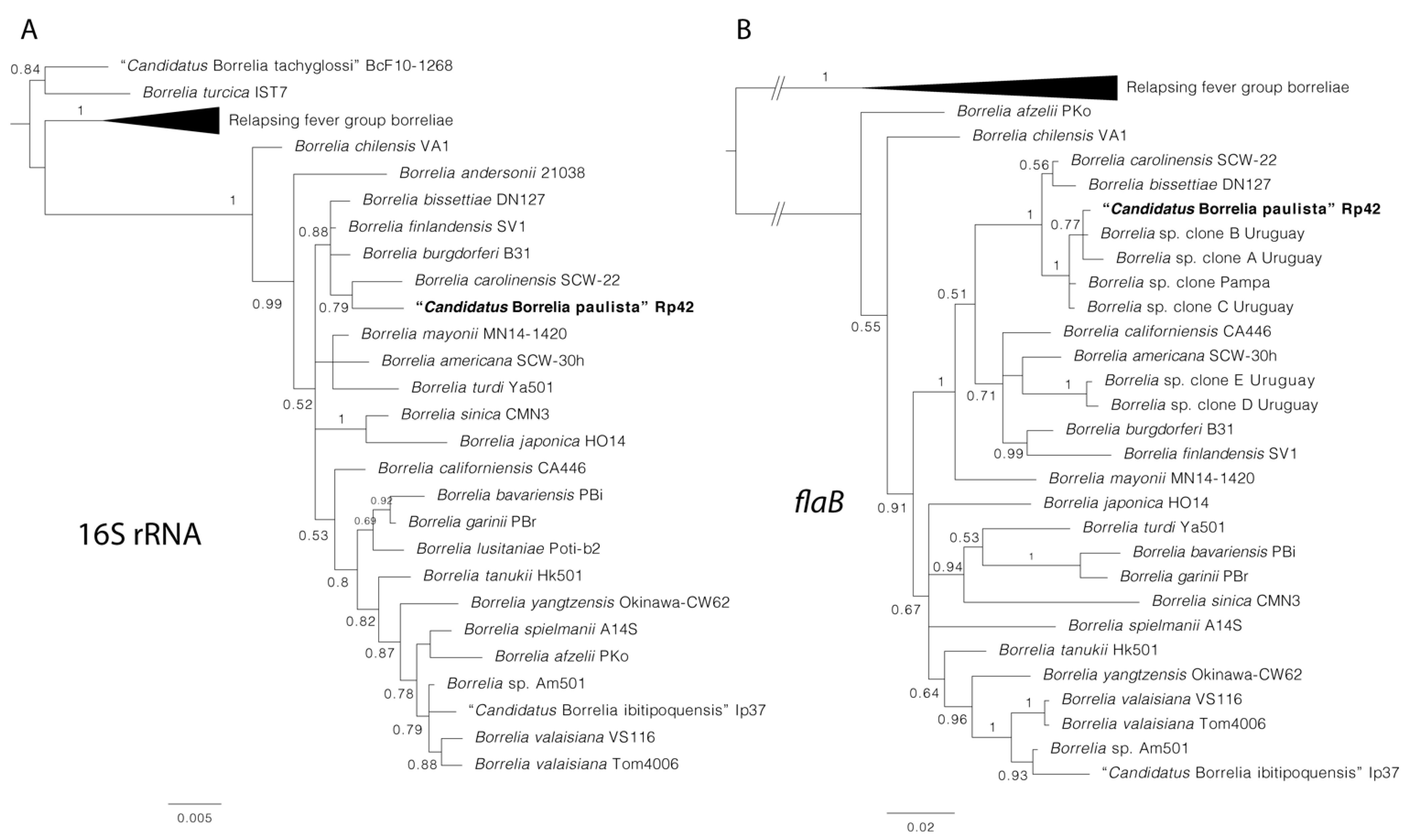
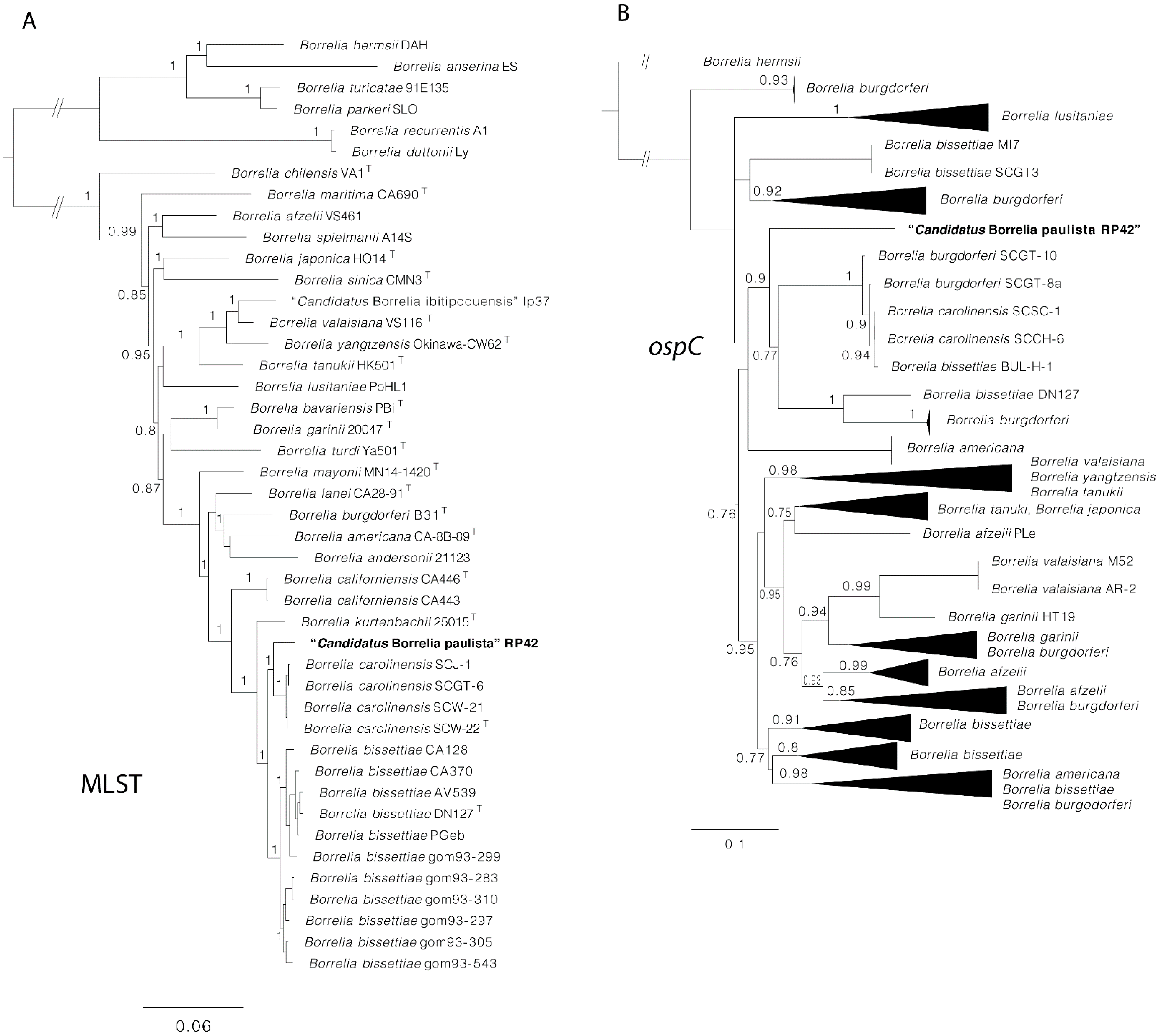

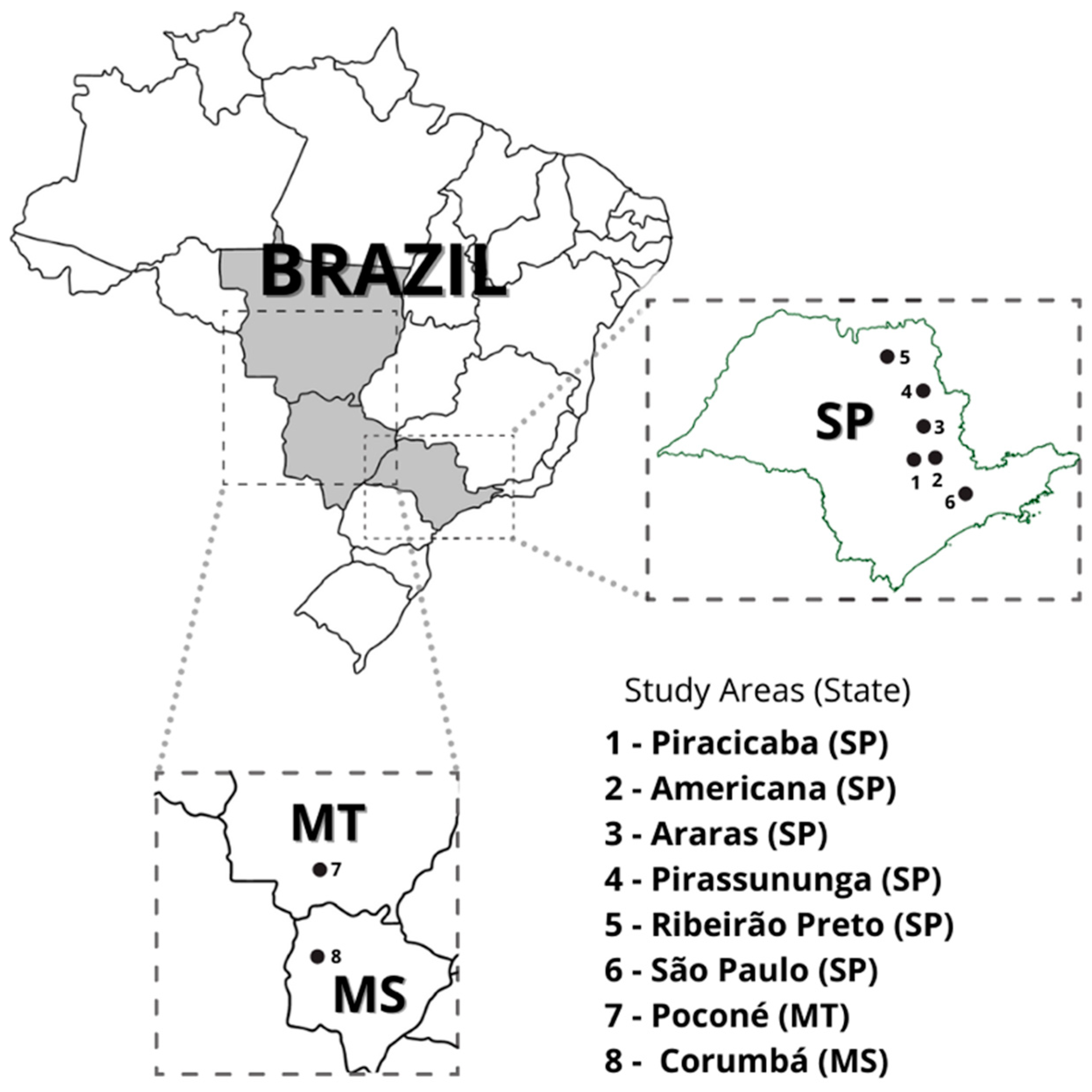{kind=link}
{kind=link}
{kind=link}
| Species | Areas | Total (Tissue Samples) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| A1 | A2 | A3 | A4 | A5 | A6 | A7 | A8 | ||
| Order Didelmorphia | |||||||||
| Didelphis albiventris | 9 | 14 | 2 | 6 | 7 | - | 1 | - | 39 |
| Didelphis aurita | 1 | - | - | - | - | 2 | - | - | 3 |
| Gracilinanus agilis | - | - | 5 | 17 | 7 | - | 28 | 2 | 59 |
| Gracilinanus microtarsus | - | 1 | 2 | - | - | - | - | 4 | 7 |
| Marmosa (Micoureus) constantinae | - | - | - | 2 | - | 2 | - | - | 4 |
| Monodelphis domestica | - | - | - | - | - | - | - | 11 | 11 |
| Phylander sp. | - | - | - | 1 | - | - | - | - | 1 |
| Order Rodentia | |||||||||
| Rattus rattus | 1 | 14 | - | - | 10 | - | - | - | 25 |
| Mus musculus | - | 1 | - | - | - | - | - | - | 1 |
| Oecomys aff. marmorae | - | - | - | 1 | - | - | 11 | 13 | 25 |
| Nectomys squamipes | - | - | - | 4 | 10 | - | - | - | 14 |
| Necromys lasiurus | - | - | 9 | 1 | - | - | 4 | - | 14 |
| Oligoryzomys spp. | 8 | 1 | 16 | 24 | 20 | - | - | - | 69 |
| Juliomys cf. ossitenuis | - | - | - | 2 | - | - | - | - | 2 |
| Akodon sp. | - | - | - | 5 | 2 | 6 | - | - | 13 |
| Hylaeamys megacephalus | - | - | - | 3 | 1 | - | 30 | - | 34 |
| Euryoryzomys russatus | - | - | 3 | - | - | 9 | - | - | 12 |
| Cavia sp. | - | - | 2 | 1 | - | - | - | - | 3 |
| Clyomis laticeps | - | - | - | - | - | - | - | 1 | 1 |
| Thrichomys pachyurus | - | - | - | - | - | - | - | 6 | 6 |
| Cerradomys sp. | - | - | - | 1 | - | - | - | - | 1 |
| Cerradomys subflavus | - | - | - | - | - | - | 1 | - | 1 |
| Dasyprocta azarae | - | - | - | - | - | - | 4 | - | 4 |
| Oecomys sp. 1 | - | - | - | - | - | - | 8 | - | 8 |
| Oecomys sp. 2 | - | - | - | - | - | - | 2 | - | 2 |
| Not identified | - | - | 3 | 1 | - | - | 16 | - | 20 |
| Order Cingulata | |||||||||
| Dasypus novemcinctus | 1 | - | - | - | - | - | - | - | 1 |
| Order Carnivora | |||||||||
| Nasua nasua | 1 | - | - | - | - | - | - | 1 | 2 |
| Total | 21 | 31 | 42 | 69 | 57 | 19 | 105 | 38 | 382 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weck, B.C.; Serpa, M.C.A.; Labruna, M.B.; Muñoz-Leal, S. A Novel Genospecies of Borrelia burgdorferi Sensu Lato Associated with Cricetid Rodents in Brazil. Microorganisms 2022, 10, 204. https://doi.org/10.3390/microorganisms10020204
Weck BC, Serpa MCA, Labruna MB, Muñoz-Leal S. A Novel Genospecies of Borrelia burgdorferi Sensu Lato Associated with Cricetid Rodents in Brazil. Microorganisms. 2022; 10(2):204. https://doi.org/10.3390/microorganisms10020204
Chicago/Turabian StyleWeck, Bárbara C., Maria Carolina A. Serpa, Marcelo B. Labruna, and Sebastián Muñoz-Leal. 2022. "A Novel Genospecies of Borrelia burgdorferi Sensu Lato Associated with Cricetid Rodents in Brazil" Microorganisms 10, no. 2: 204. https://doi.org/10.3390/microorganisms10020204
APA StyleWeck, B. C., Serpa, M. C. A., Labruna, M. B., & Muñoz-Leal, S. (2022). A Novel Genospecies of Borrelia burgdorferi Sensu Lato Associated with Cricetid Rodents in Brazil. Microorganisms, 10(2), 204. https://doi.org/10.3390/microorganisms10020204