
Whole Genome Sequencing Reveals Presence of High-Risk Global Clones of Klebsiella pneumoniae Harboring Multiple Antibiotic Resistance Genes in Multiple Plasmids in Mwanza, Tanzania

Abstract
1. Introduction
2. Materials and Methods
2.1. Laboratory Procedures
2.2. Whole Genome Sequencing and In-Silico Analysis
2.3. Systematic Review
3. Results
3.1. General Overview of Genome-Sequenced Klebsiella pneumoniae
3.2. Types and Distributions of Resistance Genes in Sequenced-Klebsiella pneumoniae
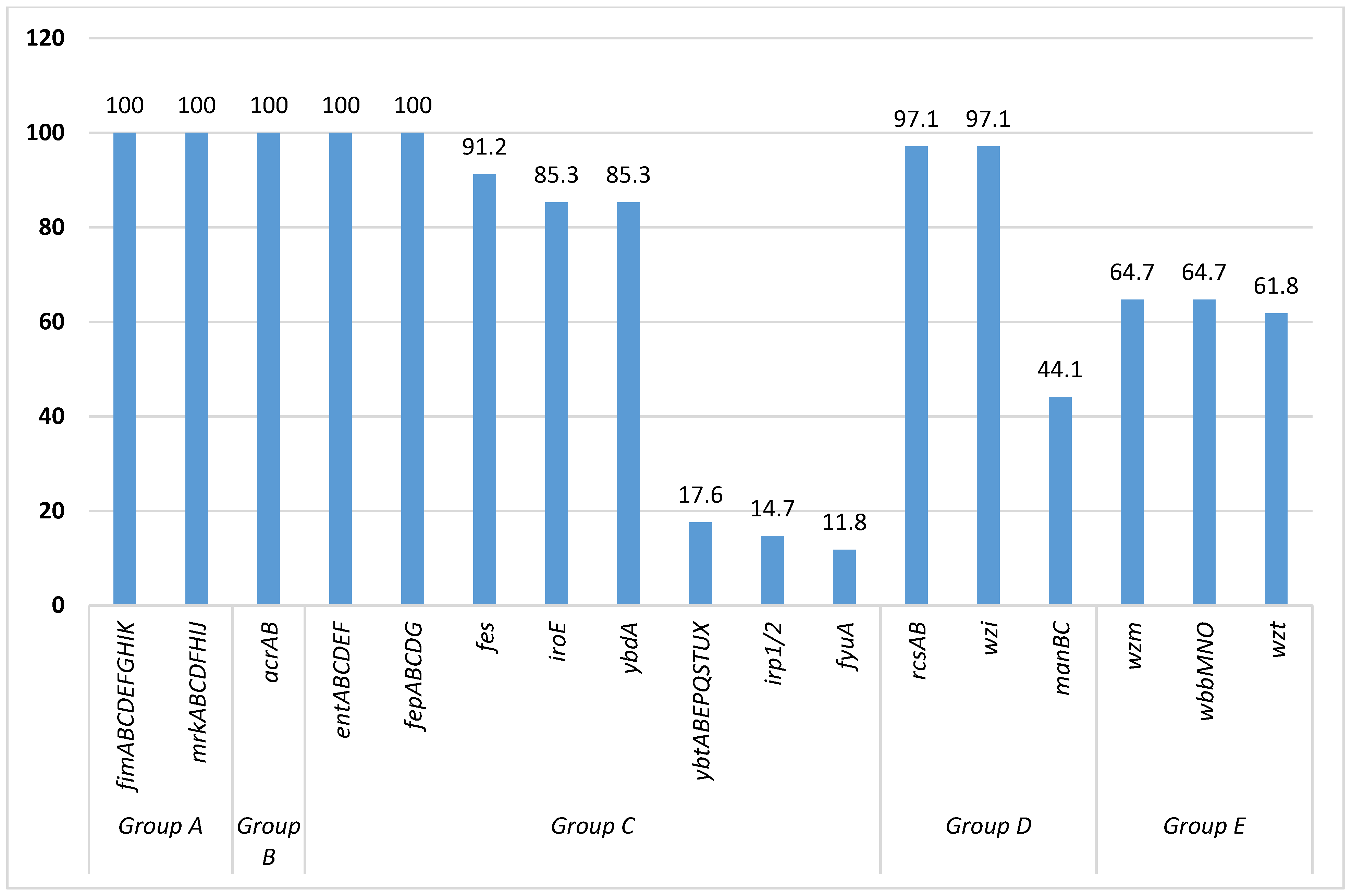
3.3. Descriptions of Acquired Virulence Genes among Sequenced K. pneumoniae
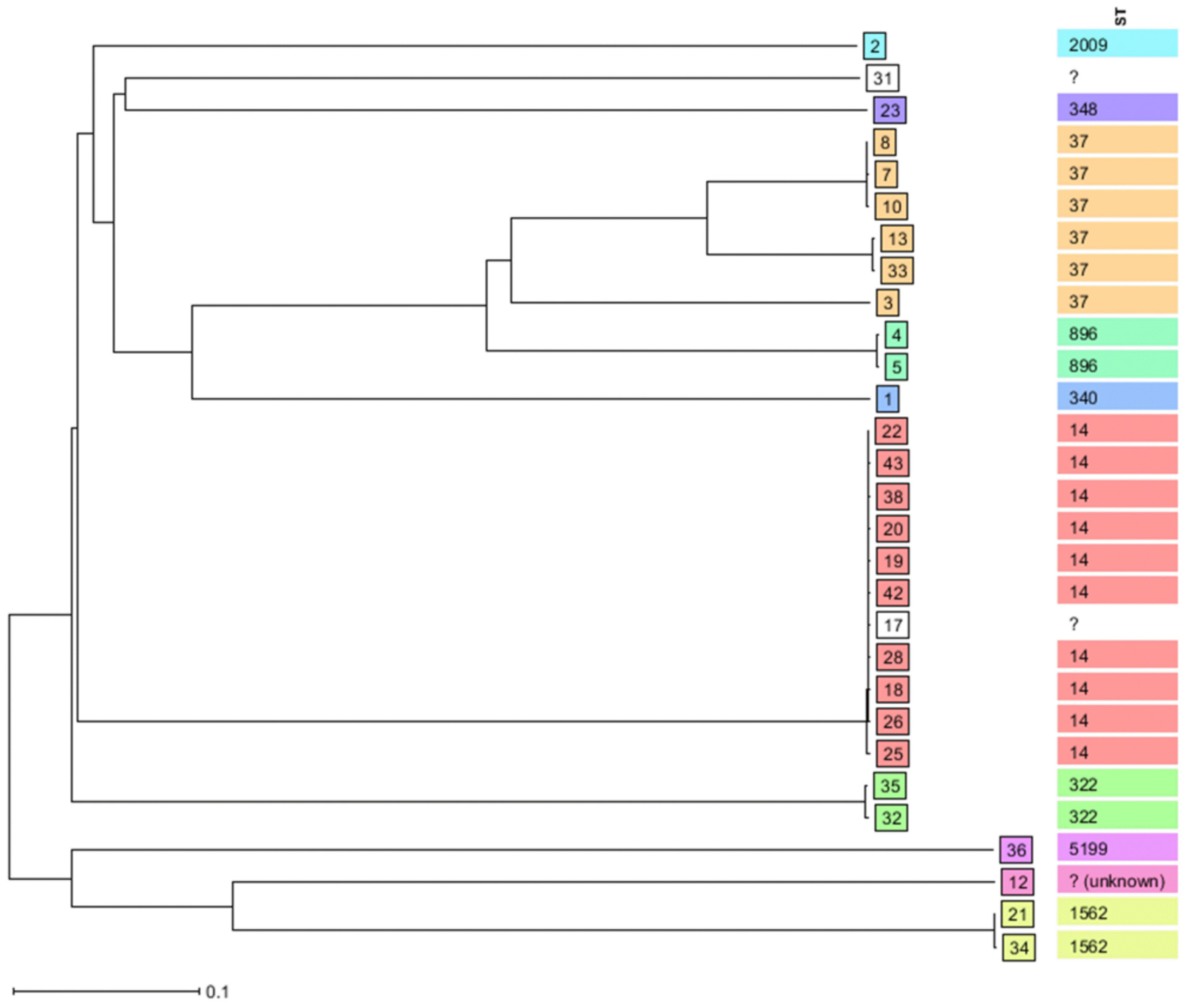
3.4. Clonal Distributions of Sequenced K. pneumoniae
3.5. Sequence Types (STs), Antibiotic Resistance Genes (ARGs), Virulence Genes (VRGs) and Plasmid Replicons Circulating between 2011 and 2022 in Mwanza among Enterobacterales
3.6. Plasmids Harboring Genes Encoding for Antibiotics Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballén, V.; Gabasa, Y.; Ratia, C.; Ortega, R.; Tejero, M.; Soto, S. Antibiotic resistance and virulence profiles of Klebsiella pneumoniae strains isolated from different clinical sources. Front. Cell. Infect. Microbiol. 2021, 11, 738223. [Google Scholar] [CrossRef] [PubMed]
- Silago, V.; Kovacs, D.; Msanga, D.R.; Seni, J.; Matthews, L.; Oravcová, K.; Zadoks, R.N.; Lupindu, A.M.; Hoza, A.S.; Mshana, S.E. Bacteremia in critical care units at Bugando Medical Centre, Mwanza, Tanzania: The role of colonization and contaminated cots and mothers’ hands in cross-transmission of multidrug resistant Gram-negative bacteria. Antimicrob. Resist. Infect. Control 2020, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Kishimbo, P.; Sogone, N.M.; Kalokola, F.; Mshana, S.E. Prevalence of gram negative bacteria causing community acquired pneumonia among adults in Mwanza City, Tanzania. Pneumonia 2020, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- Edward, E.A.; Mohamed, N.M.; Zakaria, A.S. Whole Genome Characterization of the High-Risk Clone ST383 Klebsiella pneumoniae with a Simultaneous Carriage of blaCTX-M-14 on IncL/M Plasmid and blaCTX-M-15 on Convergent IncHI1B/IncFIB Plasmid from Egypt. Microorganisms 2022, 10, 1097. [Google Scholar] [CrossRef]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef]
- Dadgostar, P. Antimicrobial resistance: Implications and costs. Infect. Drug Resist. 2019, 12, 3903. [Google Scholar] [CrossRef]
- Asokan, G.V.; Vanitha, A. WHO global priority pathogens list on antibiotic resistance: An urgent need for action to integrate One Health data. Perspect. Public Health 2018, 138, 87–88. [Google Scholar]
- Enany, S.; Zakeer, S.; Diab, A.A.; Bakry, U.; Sayed, A.A. Whole genome sequencing of Klebsiella pneumoniae clinical isolates sequence type 627 isolated from Egyptian patients. PLoS ONE 2022, 17, e0265884. [Google Scholar] [CrossRef]
- Mshana, S.E.; Hain, T.; Domann, E.; Lyamuya, E.F.; Chakraborty, T.; Imirzalioglu, C. Predominance of Klebsiella pneumoniae ST14 carrying CTX-M-15 causing neonatal sepsis in Tanzania. BMC Infect. Dis. 2013, 13, 466. [Google Scholar] [CrossRef]
- Marando, R.; Seni, J.; Mirambo, M.M.; Falgenhauer, L.; Moremi, N.; Mushi, M.F.; Kayange, N.; Manyama, F.; Imirzalioglu, C.; Chakraborty, T. Predictors of the extended-spectrum-beta lactamases producing Enterobacteriaceae neonatal sepsis at a tertiary hospital, Tanzania. Int. J. Med. Microbiol. 2018, 308, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Giske, C.G.; Fröding, I.; Hasan, C.M.; Turlej-Rogacka, A.; Toleman, M.; Livermore, D.; Woodford, N.; Walsh, T.R. Diverse sequence types of Klebsiella pneumoniae contribute to the dissemination of bla NDM-1 in India, Sweden, and the United Kingdom. Antimicrob. Agents Chemother. 2012, 56, 2735–2738. [Google Scholar] [CrossRef] [PubMed]
- Spadar, A.; Phelan, J.; Elias, R.; Modesto, A.; Caneiras, C.; Marques, C.; Lito, L.; Pinto, M.; Cavaco-Silva, P.; Ferreira, H. Genomic epidemiological analysis of Klebsiella pneumoniae from Portuguese hospitals reveals insights into circulating antimicrobial resistance. Sci. Rep. 2022, 12, 13791. [Google Scholar] [CrossRef] [PubMed]
- Ragupathi, N.K.D.; Bakthavatchalam, Y.D.; Mathur, P.; Pragasam, A.K.; Walia, K.; Ohri, V.; Veeraraghavan, B. Plasmid profiles among some ESKAPE pathogens in a tertiary care centre in south India. Indian J. Med. Res. 2019, 149, 222. [Google Scholar] [PubMed]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [PubMed]
- Mirzaie, A.; Ranjbar, R. Antibiotic resistance, virulence-associated genes analysis and molecular typing of Klebsiella pneumoniae strains recovered from clinical samples. AMB Express 2021, 11, 122. [Google Scholar] [CrossRef]
- Ranjbar, R.; Kelishadrokhi, A.F.; Chehelgerdi, M. Molecular characterization, serotypes and phenotypic and genotypic evaluation of antibiotic resistance of the Klebsiella pneumoniae strains isolated from different types of hospital-acquired infections. Infect. Drug Resist. 2019, 12, 603. [Google Scholar] [CrossRef]
- Olusoga, O.D.; Majeed, A.A.; Sunday, O.A.; Adegboyega, O.A.; Alexander, W.M. Emergence of Diarrhoeagenic Klebsiella pneumoniae Carrying astA and senB genes in Nigeria. Afr. J. Microbiol. Res. 2022, 16, 264–267. [Google Scholar]
- Hudzicki, J. Kirby-Bauer disk diffusion susceptibility test protocol. Am. Soc. Microbiol. 2009, 15, 55–63. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- Jünemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; Von Haeseler, A.; Stoye, J. Updating benchtop sequencing performance comparison. Nat. Biotechnol. 2013, 31, 294–296. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Schwengers, O.; Barth, P.; Falgenhauer, L.; Hain, T.; Chakraborty, T.; Goesmann, A. Platon: Identification and characterization of bacterial plasmid contigs in short-read draft assemblies exploiting protein sequence-based replicon distribution scores. Microb. Genom. 2020, 6, mgen000398. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Moremi, N.; Manda, E.V.; Falgenhauer, L.; Ghosh, H.; Imirzalioglu, C.; Matee, M.; Chakraborty, T.; Mshana, S.E. Predominance of CTX-M-15 among ESBL producers from environment and fish gut from the shores of Lake Victoria in Mwanza, Tanzania. Front. Microbiol. 2016, 7, 1862. [Google Scholar] [CrossRef] [PubMed]
- Xanthopoulou, K.; Imirzalioglu, C.; Walker, S.V.; Behnke, M.; Dinkelacker, A.G.; Eisenbeis, S.; Gastmeier, P.; Gölz, H.; Käding, N.; Kern, W.V. Surveillance and Genomic Analysis of Third-Generation Cephalosporin-Resistant and Carbapenem-Resistant Klebsiella pneumoniae Complex in Germany. Antibiotics 2022, 11, 1286. [Google Scholar] [CrossRef]
- Esteban-Cantos, A.; Aracil, B.; Bautista, V.; Ortega, A.; Lara, N.; Saez, D.; Fernández-Romero, S.; Pérez-Vázquez, M.; Navarro, F.; Grundmann, H. The carbapenemase-producing Klebsiella pneumoniae population is distinct and more clonal than the carbapenem-susceptible population. Antimicrob. Agents Chemother. 2017, 61, e02520-16. [Google Scholar] [CrossRef]
- Moubareck, C.A.; Mouftah, S.F.; Pál, T.; Ghazawi, A.; Halat, D.H.; Nabi, A.; AlSharhan, M.A.; AlDeesi, Z.O.; Peters, C.C.; Celiloglu, H. Clonal emergence of Klebsiella pneumoniae ST14 co-producing OXA-48-type and NDM carbapenemases with high rate of colistin resistance in Dubai, United Arab Emirates. Int. J. Antimicrob. Agents 2018, 52, 90–95. [Google Scholar] [CrossRef]
- Mouftah, S.F.; Pál, T.; Higgins, P.G.; Ghazawi, A.; Idaghdour, Y.; Alqahtani, M.; Omrani, A.S.; Rizvi, T.A.; Sonnevend, Á. Diversity of carbapenem-resistant Klebsiella pneumoniae ST14 and emergence of a subgroup with KL64 capsular locus in the Arabian Peninsula. Eur. J. Clin. Microbiol. Infect. Dis. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, W.; Xu, X.; Zhou, X.; Shi, C. Transmissible ST3-IncHI2 plasmids are predominant carriers of diverse complex IS 26-Class 1 integron arrangements in multidrug-resistant Salmonella. Front. Microbiol. 2018, 9, 2492. [Google Scholar] [CrossRef] [PubMed]
- Minja, C.A.; Shirima, G.; Mshana, S.E. Conjugative plasmids disseminating ctx-m-15 among human, animals and the environment in Mwanza Tanzania: A need to intensify one health approach. Antibiotics 2021, 10, 836. [Google Scholar] [CrossRef] [PubMed]
- Bi, D.; Zheng, J.; Li, J.-J.; Sheng, Z.-K.; Zhu, X.; Ou, H.-Y.; Li, Q.; Wei, Q. In silico typing and comparative genomic analysis of IncFIIK plasmids and insights into the evolution of replicons, plasmid backbones, and resistance determinant profiles. Antimicrob. Agents Chemother. 2018, 62, e00764-18. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Fang, T.; Zhou, X.; Zhang, D.; Shi, X.; Shi, C. IncHI2 plasmids are predominant in antibiotic-resistant Salmonella isolates. Front. Microbiol. 2016, 7, 1566. [Google Scholar] [CrossRef] [PubMed]
- Shankar, C.; Sethuvel, D.P.M.; Neeravi, A.R.; Venkatesan, M.; Ragupathi, N.K.D.; Anandan, S.; Veeraraghavan, B. Identification of plasmids by PCR based replicon typing in bacteremic Klebsiella pneumoniae. Microb. Pathog. 2020, 148, 104429. [Google Scholar] [CrossRef]
- Qu, D.; Shen, Y.; Hu, L.; Jiang, X.; Yin, Z.; Gao, B.; Zhao, Y.; Yang, W.; Yang, H.; Han, J. Comparative analysis of KPC-2-encoding chimera plasmids with multi-replicon IncR: IncpA1763-KPC: IncN1 or IncFIIpHN7A8: IncpA1763-KPC: IncN1. Infect. Drug Resist. 2019, 12, 285. [Google Scholar] [CrossRef]
- Fuga, B.; Cerdeira, L.; Moura, Q.; Fontana, H.; Fuentes-Castillo, D.; Carvalho, A.C.; Lincopan, N. Genomic data reveals the emergence of an IncQ1 small plasmid carrying blaKPC-2 in Escherichia coli of the pandemic sequence type 648. J. Glob. Antimicrob. Resist. 2021, 25, 8–13. [Google Scholar] [CrossRef]
- Damiano, P.; Salema, E.J.; Silago, V. The susceptibility of multidrug resistant and biofilm forming Klebsiella pneumoniae and Escherichia coli to antiseptic agents used for preoperative skin preparations at zonal referral hospital in Mwanza, Tanzania. Malawi Med. J. 2021, 33, 59–64. [Google Scholar]
- Jesumirhewe, C.; Springer, B.; Allerberger, F.; Ruppitsch, W. Genetic Characterization of Antibiotic Resistant Enterobacteriaceae Isolates from Bovine Animals and the Environment in Nigeria. Front. Microbiol. 2022, 13, 793541. [Google Scholar] [CrossRef]
- Donner, L.; Staley, Z.R.; Petali, J.; Sangster, J.; Li, X.; Mathews, W.; Snow, D.; Howe, A.; Soupir, M.; Bartelt-Hunt, S. The Human Health Implications of Antibiotic Resistance in Environmental Isolates from Two Nebraska Watersheds. Microbiol. Spectr. 2022, 10, e02082-21. [Google Scholar] [CrossRef]
- Mahmoudvand, H.; Azadpour, M.; Nowroozi, J.; Goudarzi, G. Presence of qacEΔ1 and cepA genes and susceptibility to a hospital biocide in clinical isolates of Klebsiella pneumoniae in Iran. Trop. Biomed. 2015, 32, 109–115. [Google Scholar]
- Abuzaid, A.; Hamouda, A.; Amyes, S. Klebsiella pneumoniae susceptibility to biocides and its association with cepA, qacΔE and qacE efflux pump genes and antibiotic resistance. J. Hosp. Infect. 2012, 81, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Haeggman, S.; Löfdahl, S.; Paauw, A.; Verhoef, J.; Brisse, S. Diversity and evolution of the class A chromosomal beta-lactamase gene in Klebsiella pneumoniae. Antimicrob Agents Chemother. 2004, 48, 2400–2408. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L.A. Virulence factors and their mechanisms of action: The view from a damage–response framework. J. Water Health 2009, 7, S2–S18. [Google Scholar] [CrossRef]
- Peterson, J.W. Bacterial pathogenesis. In Medical Microbiology, 4th ed.; University of Texas Medical Branch: Galveston, TX, USA, 1996. [Google Scholar]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]
- Gorrie, C.L.; Mirčeta, M.; Wick, R.R.; Judd, L.M.; Lam, M.; Gomi, R.; Abbott, I.J.; Thomson, N.R.; Strugnell, R.A.; Pratt, N.F. Genomic dissection of Klebsiella pneumoniae infections in hospital patients reveals insights into an opportunistic pathogen. Nat. Commun. 2022, 13, 3017. [Google Scholar] [CrossRef]
- Silago, V.; Kovacs, D.; Samson, H.; Seni, J.; Matthews, L.; Oravcová, K.; Lupindu, A.M.; Hoza, A.S.; Mshana, S.E. Existence of multiple ESBL genes among phenotypically confirmed ESBL producing Klebsiella pneumoniae and Escherichia coli concurrently isolated from clinical, colonization and contamination samples from neonatal units at Bugando Medical Center, Mwanza, Tanzania. Antibiotics 2021, 10, 476. [Google Scholar]



{kind=link}
{kind=link}
| Characteristics | Frequency (n) | Percentage (%) | |
|---|---|---|---|
| Sequence types (STs; N = 34) | Determined STs | 28 | 82.4 |
| Undetermined STs | 6 | 17.6 | |
| Types of STs identified (N = 28) | ST14 | 11 | 39.3 |
| ST37 | 6 | 21.4 | |
| ST1562 | 2 | 7.1 | |
| ST322 | 2 | 7.1 | |
| ST896 | 2 | 7.1 | |
| Other STs (ST2009, ST45, ST5199, ST384, ST340) | 5 | 17.9 | |
| Bacteria harboring typeable plasmid-replicons (N = 34) | Yes | 18 | 52.9 |
| No | 16 | 47.1 | |
| Amount of typeable plasmid-replicons in each bacteria (N = 18) | 1 | 14 | 77.8 |
| 2 | 2 | 11.1 | |
| 3 | 1 | 5.6 | |
| 4 | 1 | 5.6 | |
| Types of plasmid replicons (N = 25) | IncFIB(K) | 12 | 48.0 |
| IncFII(K) | 4 | 16.0 | |
| IncQ1 | 3 | 12.0 | |
| IncR | 3 | 12.0 | |
| Others [(IncHI2/2A, n=2) and (ColRNAI, n = 1)] | 3 | 12.0 | |
| Antibiotics/biocides-resistance genes | Yes | 32 | 94.1 |
| No | 2 | 5.9 | |
| Acquired virulence genes | Yes | 34 | 100 |
| No | 0 | 0 | |
| Types of Resistance Genes | Frequency (n) | Percentages (%) | |
|---|---|---|---|
| Β-lactam resistance | blaCTX-M-15 | 29 | 90.6 |
| blaTEM-1B | 27 | 84.4 | |
| blaSHV-100 | 10 | 31.3 | |
| blaSCO-1 | 10 | 31.3 | |
| blaOXA-1 | 4 | 12.5 | |
| blaLEN10 | 2 | 6.3 | |
| blaSHV-148 | 1 | 3.1 | |
| Aminoglycosides resistance | aph(3′)-Ib | 16 | 50 |
| aph(6)-Id | 16 | 50 | |
| aph(3′)-III | 1 | 3.1 | |
| aph(3′)-Ia | 2 | 6.3 | |
| aac(3)-IIa | 15 | 46.9 | |
| aac(3)-IId | 13 | 46.9 | |
| aac(6′)-Ib-cr | 4 | 12.4 | |
| strA | 16 | 50 | |
| strB | 16 | 50 | |
| aadA1 | 2 | 6.3 | |
| aadA2 | 3 | 9.4 | |
| Trimethoprim resistance | dfrA1 | 1 | 3.1 |
| dfrA7 | 1 | 3.1 | |
| dfrA12 | 2 | 6.3 | |
| dfrA14 | 4 | 12.5 | |
| dfrA30 | 13 | 46.9 | |
| dfrG | 2 | 6.3 | |
| Sulfamethoxazole resistance | sul2 | 27 | 84.4 |
| sul1 | 3 | 9.4 | |
| Quinolones resistance | OqxA | 27 | 84.4 |
| OqxB | 28 | 87.5 | |
| qnrB1 | 2 | 6.3 | |
| qnrB66 | 1 | 3.1 | |
| aac(6′)-Ib-cr | 2 | 6.3 | |
| Tetracycline resistance | tet(A) | 4 | 12.5 |
| tet(M) | 2 | 6.3 | |
| tet(L) | 1 | 3.1 | |
| Aminoglycosides/quinolones resistance | aac(3)-IIa | 14 | 43.8 |
| aac(6′)-Ib-cr | 4 | 12.5 | |
| Aminocyclitol/aminoglycosides resistance | aadA1 | 1 | 3.1 |
| aadA2 | 1 | 3.1 | |
| Biocides/antibiotics resistance | OqxA | 5 | 15.6 |
| OqxB | 4 | 12.5 | |
| Biocides resistance | qacE | 4 | 12.5 |
| Isolate No. | ST | Plasmids | Antibiotic Classes and Respective Genes Encoding for Their Resistance, Harbored in Plasmids | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Beta-Lactams | Aminoglycosides | Sulfamethoxazole | Trimethoprim | Tetracyclines | Quinolones | Biocides | |||
| ESBL 21 | 45 | IncFIB IncFII | blaCTX-M-15 blaTEM-1B blaOXA-1 | aac(6′)-Ib-cr aph(3″)-Ib aac(3)-IIa aph(6)-Id | sul2 | dfrA14 | tet(A) | aac(6′)-Ib-cr qnrB1 | |
| ESBL 88 | 1562 | IncR | blaCTX-M-15 blaTEM-1B | aac(3)-IIa | sul2 | ||||
| ESBL 219 | 322 | IncFIB IncFII IncR | blaCTX-M-15 blaTEM-1B | aac(3)-IId | |||||
| ESBL 428 | 1155 | IncFII IncQ1 IncR | blaCTX-M-15 blaTEM-1B | aph(3″)-Ib aac(3)-IId | sul1 | dfrA7 dfrA30 | qacE | ||
| ESBL 703 | 14 | IncFIB IncFII | blaCTX-M-15 blaTEM-1B blaSCO-1 | aac(3)-IIa aph(6)-Id aph(3″)-Ib aac(3)-IId | sul2 | ||||
| ESBL 1270 | 340 | IncF ColRNAI IncR | blaCTX-M-15 blaOXA-1 | aac(6″)-Ib-cr aph(3″)-Ia aadA2 | sul1 | dfrA12 | tet(A) | qacE | |
| ESBL 1358 | 37 | IncFII IncR | blaCTX-M-15 blaTEM-1B | sul2 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silago, V.; Mshana, S.E. Whole Genome Sequencing Reveals Presence of High-Risk Global Clones of Klebsiella pneumoniae Harboring Multiple Antibiotic Resistance Genes in Multiple Plasmids in Mwanza, Tanzania. Microorganisms 2022, 10, 2396. https://doi.org/10.3390/microorganisms10122396
Silago V, Mshana SE. Whole Genome Sequencing Reveals Presence of High-Risk Global Clones of Klebsiella pneumoniae Harboring Multiple Antibiotic Resistance Genes in Multiple Plasmids in Mwanza, Tanzania. Microorganisms. 2022; 10(12):2396. https://doi.org/10.3390/microorganisms10122396
Chicago/Turabian StyleSilago, Vitus, and Stephen E. Mshana. 2022. "Whole Genome Sequencing Reveals Presence of High-Risk Global Clones of Klebsiella pneumoniae Harboring Multiple Antibiotic Resistance Genes in Multiple Plasmids in Mwanza, Tanzania" Microorganisms 10, no. 12: 2396. https://doi.org/10.3390/microorganisms10122396
APA StyleSilago, V., & Mshana, S. E. (2022). Whole Genome Sequencing Reveals Presence of High-Risk Global Clones of Klebsiella pneumoniae Harboring Multiple Antibiotic Resistance Genes in Multiple Plasmids in Mwanza, Tanzania. Microorganisms, 10(12), 2396. https://doi.org/10.3390/microorganisms10122396