

Hymenolepis diminuta Reduce Lactic Acid Bacterial Load and Induce Dysbiosis in the Early Infection of the Probiotic Colonization of Swiss Albino Rat
 ,
,
Abstract
1. Introduction
2. Materials and Method
2.1. Maintenance of the Parasite
2.2. Sample Collection and Determination of Bacterial Load
2.3. Metagenomic Sequencing of the Gut Sample
2.4. Isolation and Purification of LAB
2.5. Confirmatory Tests of the Isolated LAB Colony
2.5.1. Potassium Hydroxide (KOH) Test
2.5.2. Catalase Test
2.5.3. Spore Staining
2.5.4. Characterization of Probiotic Properties through the In-Vitro Process
- Growth at Different pH Level: The determination of the pH tolerance of isolated bacteria was determined following the adopted protocol from Liong and Shah, 2005 [32].
- Bile Salt Tolerance Test: This was determined following the method adopted from Liong and Shah, 2005 [32]. In brief, 2% Bile salt (Oxgall, Hi-Media, Mumbai, India) was mixed with MRS broth in test tubes. Each test tube was inoculated with 100 µL fresh culture of isolated LAB and incubated at 37 °C for 48 h, and the growth was observed.
- Curd Production: For the determination of the Curd production test of isolated bacteria, the standard 4% of bacteria culture was added to 5 ml of boiled and cooled milk, incubated for 24 h, and then observed.
2.5.5. Morphology of LAB Bacteria
2.5.6. Growth at Different NaCl Concentration
2.5.7. Antimicrobial Activity, Hemolytic Activity and Cell Adhesion Property
2.5.8. Antibiotic Susceptibility Test
2.6. Phylogenetic Analysis of Probiotic LAB Isolates Using 16S rRNA Sequencing
3. Result
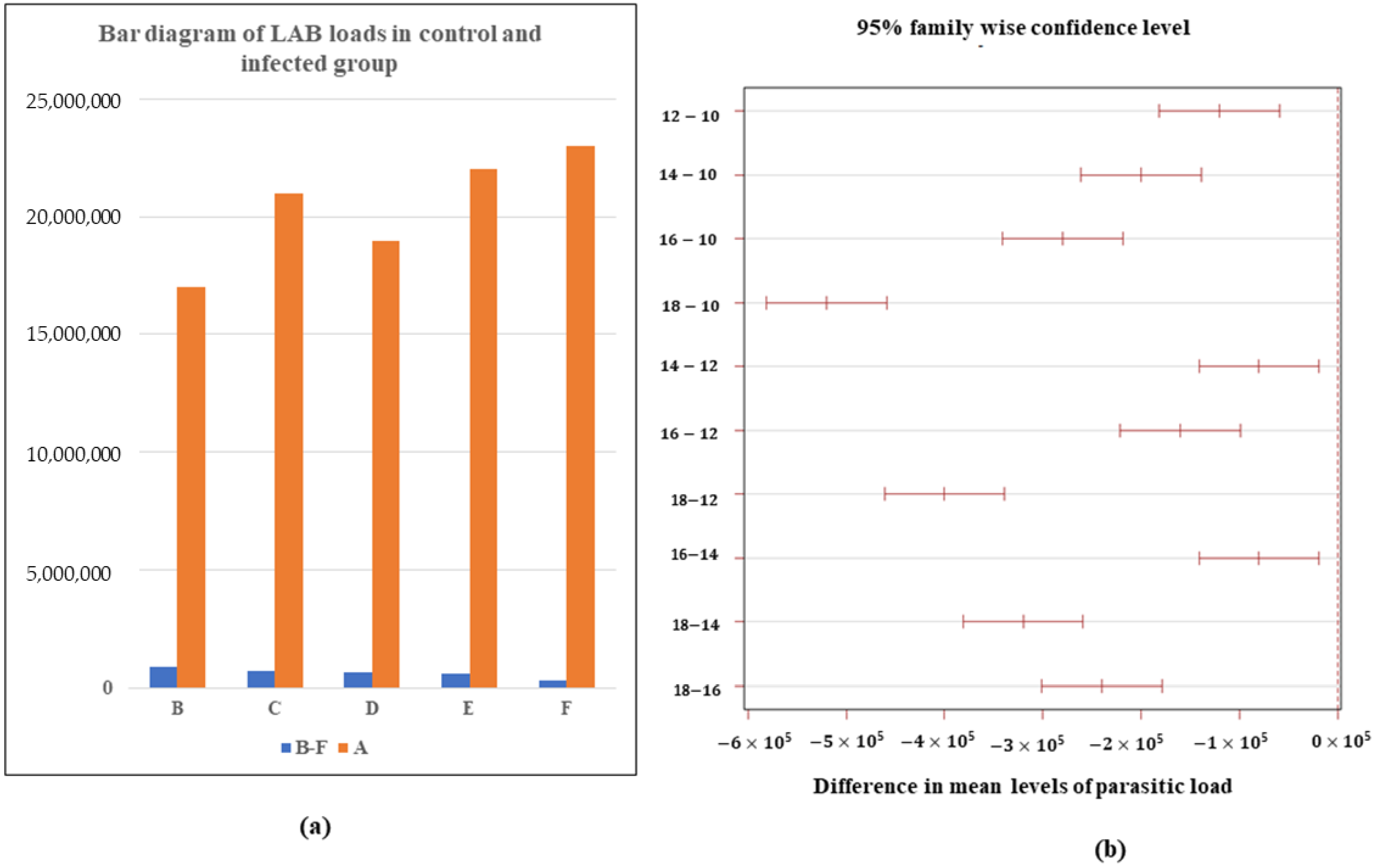
3.1. Determination of LAB Load
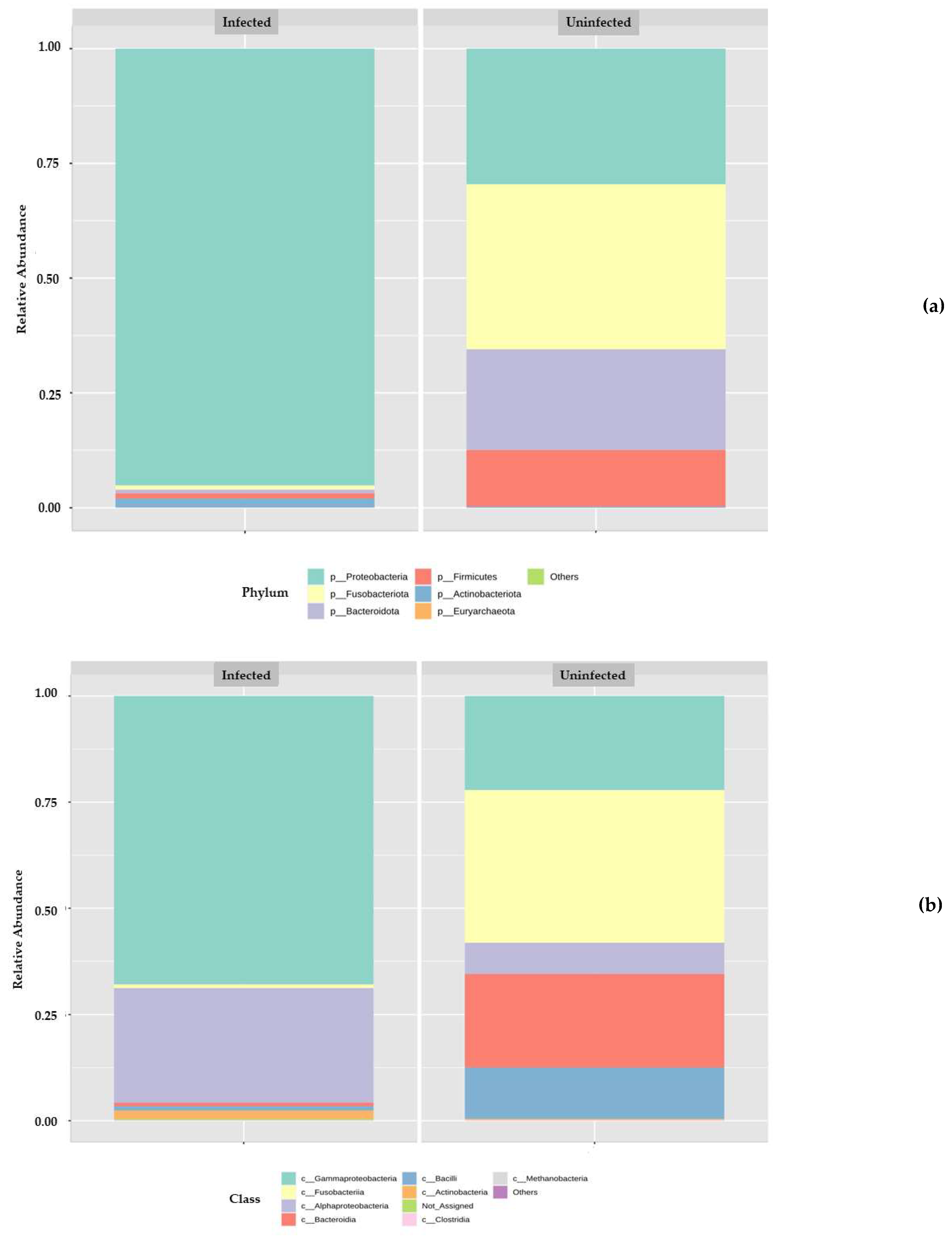
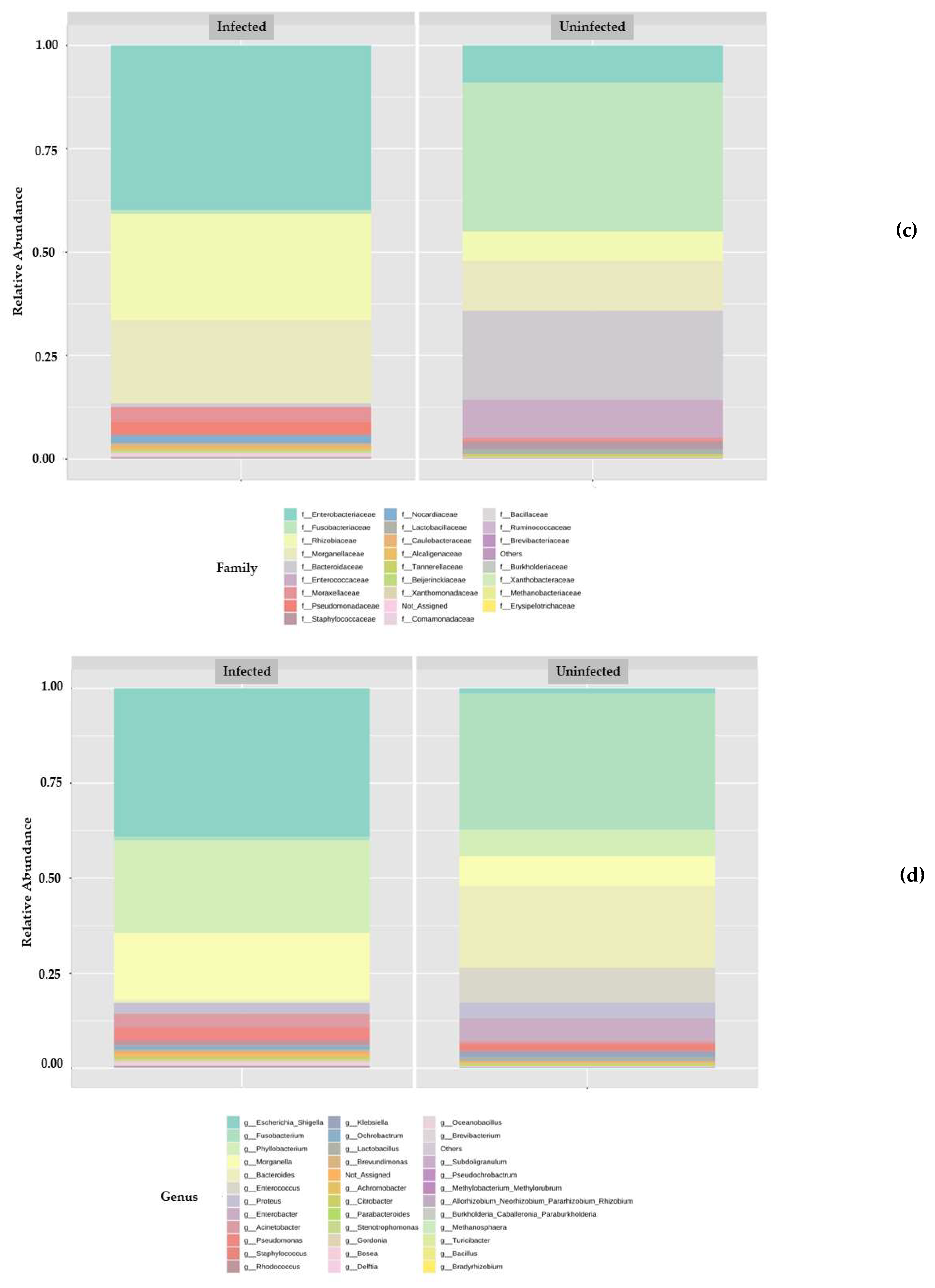
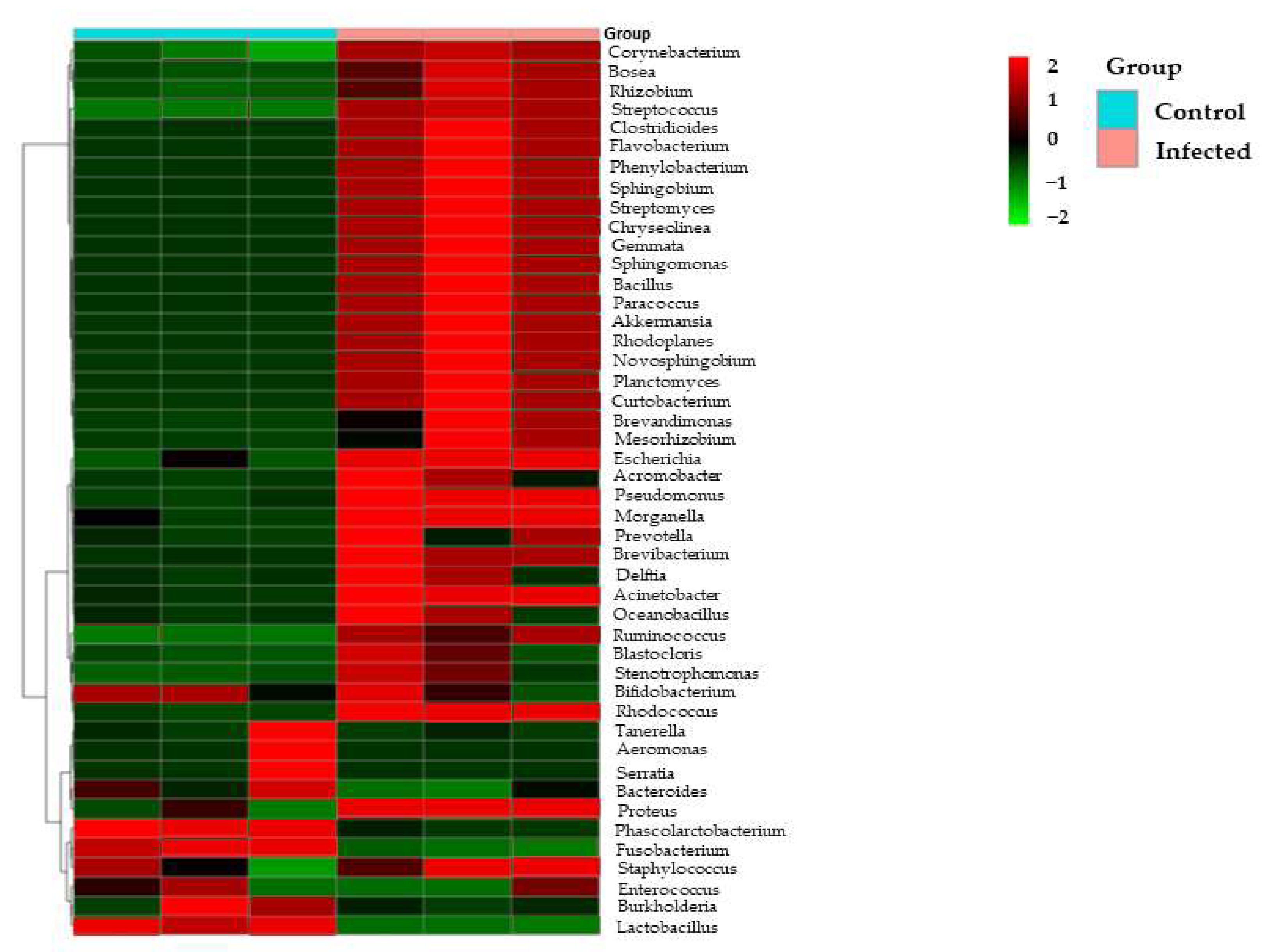
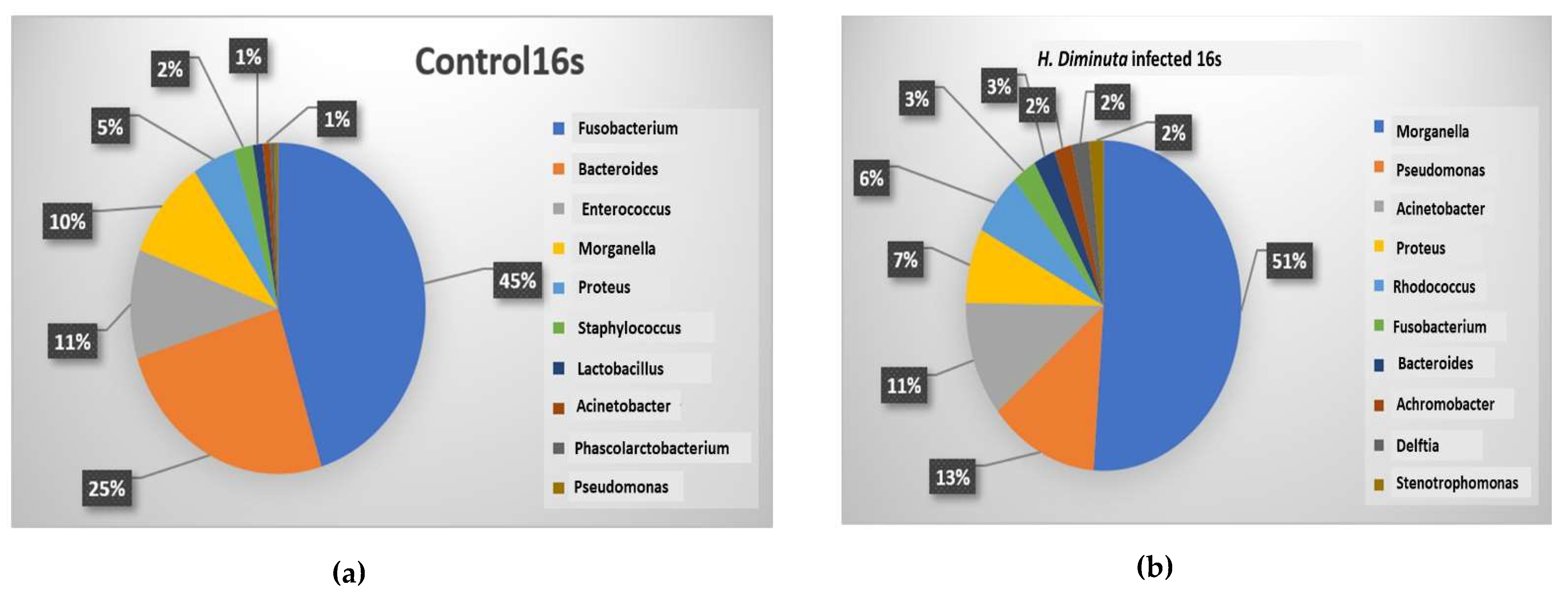
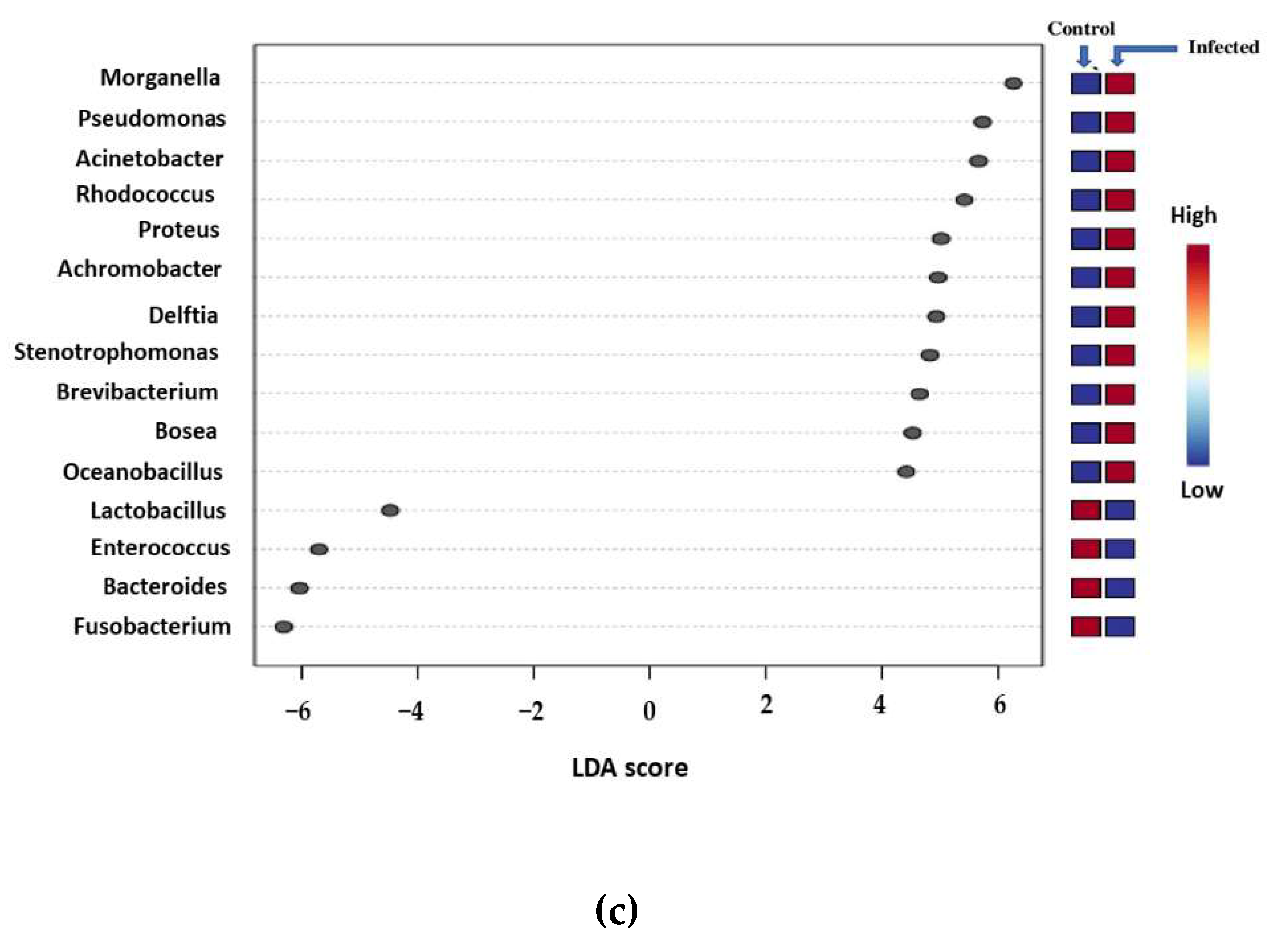
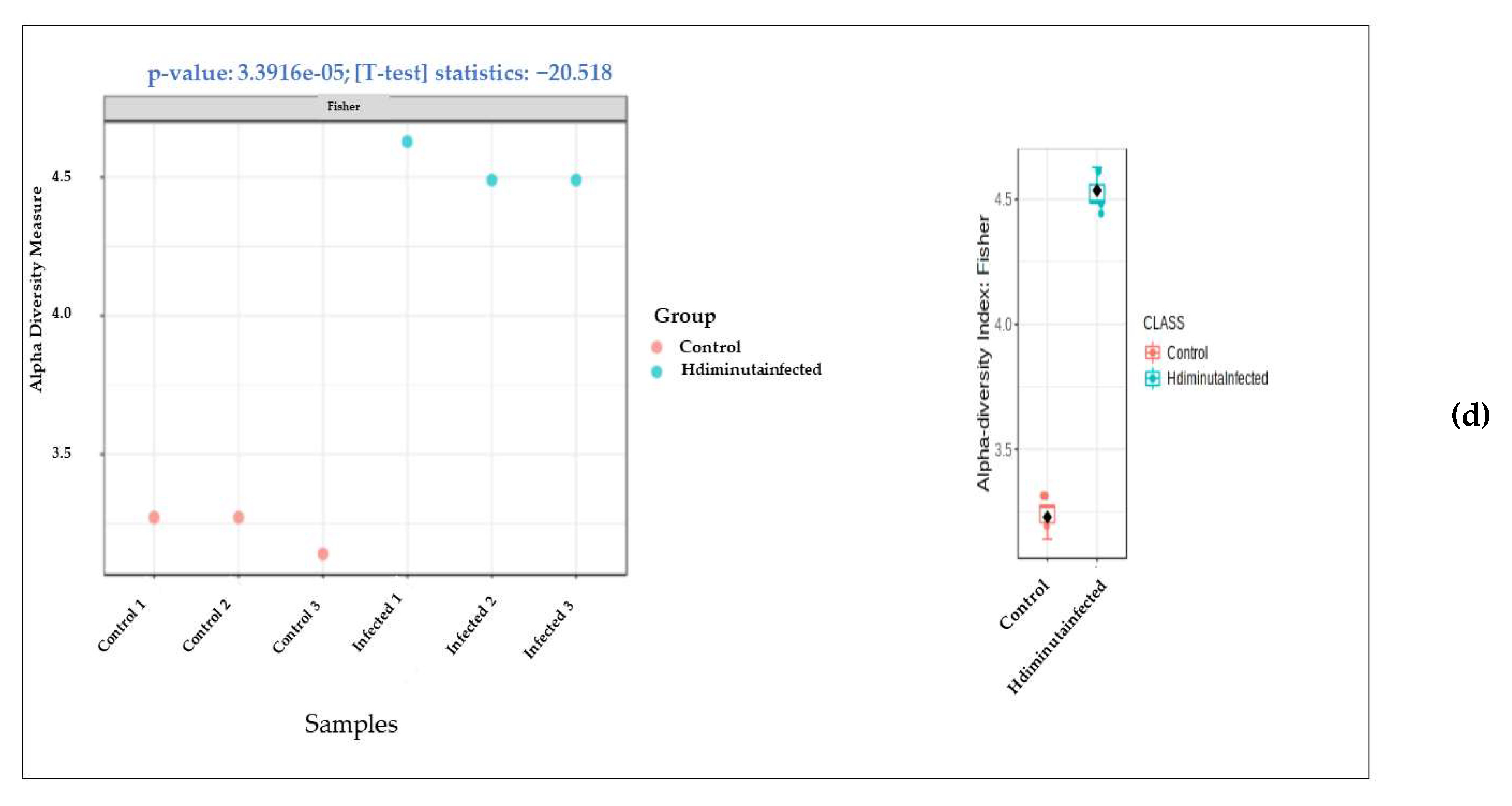
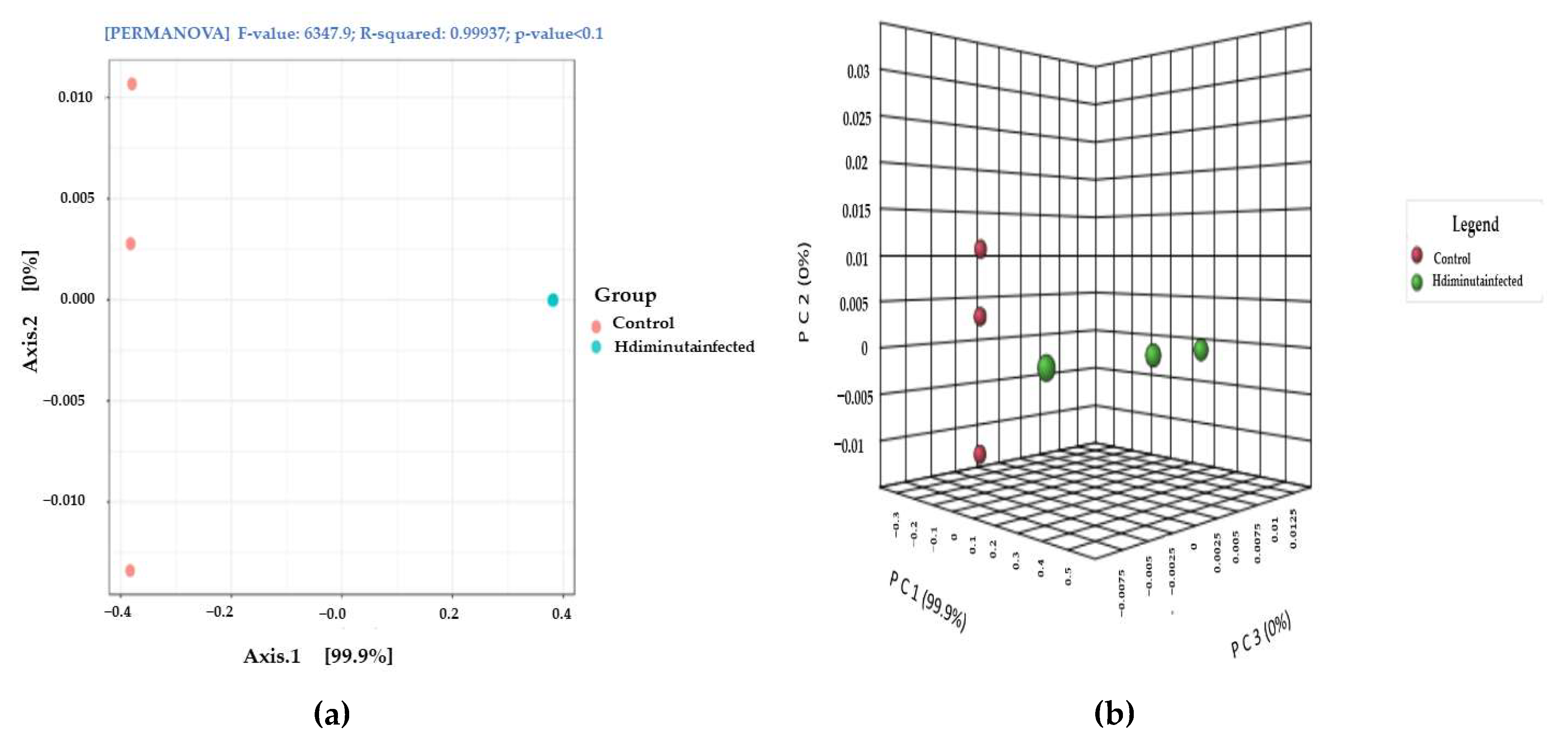
3.2. Metagenomic Analysis
3.3. Isolation and Purification of LAB
3.4. Probiotic Confirmation Tests of LAB Isolates
3.5. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tiwari, S.; Karuna, T.; Rautaraya, B. Hymenolepis diminuta infection in a child from a rural area: A rare case report. J. Lab. Physicians 2014, 6, 058–059. [Google Scholar] [CrossRef]
- Thompson, R.C.A. Neglected zoonotic helminths: Hymenolepis nana, Echinococcus canadensis and Ancylostoma ceylanicum. Clin. Microbiol. Infect. 2015, 21, 426–432. [Google Scholar] [CrossRef]
- Kandi, V.; Koka, S.S.; Bhoomigari, M.R. Hymenolepiasis in a pregnant woman: A case report of Hymenolepis nana infection. Cureus 2019, 11, e3810. [Google Scholar] [CrossRef]
- Mahande, A.M.; Mahande, M.J. Prevalence of parasitic infections and associations with pregnancy complications and outcomes in northern Tanzania: A registry-based cross-sectional study. BMC Infect. Dis. 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Damtie, D.; Liyih, M. Prevalence and associated risk factors of intestinal parasitic infections among pregnant women attending antenatal care in Yifag Health Center, Northwest Ethiopia. Can. J. Infect. Dis. Med. Microbiol. 2021, 2021, 7291199. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.; Ferreira, F.S.; Atouguia, J.; Fortes, F.; Guerra, A.; Centeno-Lima, S. Infection by intestinal parasites, stunting and anemia in school-aged children from southern Angola. PLoS ONE 2015, 10, e0137327. [Google Scholar] [CrossRef] [PubMed]
- Derso, A.; Nibret, E.; Munshea, A. Prevalence of intestinal parastitic infections and associated risk factors among pregnant women attending antenatal care center at Felege Hiwot Referral Hospital, NorthWest Ethiopia. BMC Infect. Dis. 2016, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Capuano, D.M.; Rocha, G.D.M. Parasites with zoonotic potential in dog feces collected from public areas of the city of Ribeirão Preto, SP, Brazil. Rev. Bras. Epidemiol. 2006, 9, 81–86. [Google Scholar] [CrossRef]
- Fujiwara, R.T.; Zhan, B.; Mendez, S.; Loukas, A.; Bueno, L.L.; Wang, Y.; Plieskatt, J.; Oksov, Y.; Lustigman, S.; Bottazzi, M.E.; et al. Reduction of worm fecundity and canine host blood loss mediates protection against hookworm infection elicited by vaccination with recombinant Ac-16. Clin. Vaccine Immunol. 2007, 14, 281–287. [Google Scholar] [CrossRef]
- Kopp, S.R.; Coleman, G.T.; McCarthy, J.S.; Kotze, A.C. Phenotypic characterization of two Ancylostoma caninum isolates with different susceptibilities to the anthelmintic pyrantel. Antimicrob. Agents Chemother. 2008, 52, 3980–3986. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Brestoff, J.R.; Artis, D. Commensal bacteria at the interface of host metabolism and the immune system. Nat. Immunol. 2013, 14, 676–684. [Google Scholar] [CrossRef] [PubMed]
- McLean, M.H.; Dieguez, D., Jr.; Miller, L.M.; Young, H.A. Does the microbiota play a role in the pathogenesis of autoimmune disease? Gut 2015, 64, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.G.V.; Franz, C.M.; Schillinger, U.; Holzapfel, W.H. Lactobacillus spp. with in vitro probiotic properties from human faeces and traditional fermented products. Int. J. Food Microbiol. 2006, 109, 205–214. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira-Sequeira, T.C.G.; Ribeiro, C.M.; Gomes, M.I.F.V. Probiotics as potential biotherapeutic agents targeting intestinal parasites. Ciênc. Rural 2008, 38, 2670–2679. [Google Scholar] [CrossRef]
- Vine, N.G.; Leukes, W.D.; Kaiser, H.; Daya, S.; Baxter, J.; Hecht, T. Competition for attachment of aquaculture candidate probiotic and pathogenic bacteria on fish intestinal mucus. J. Fish Dis. 2004, 27, 319–326. [Google Scholar] [CrossRef]
- Ray, A.K.; Ghosh, K.; Ringø, E.J.A.N. Enzyme-producing bacteria isolated from fish gut: A review. Aquac. Nutr. 2012, 18, 465–492. [Google Scholar] [CrossRef]
- Naveed, A.; Abdullah, S. Impact of parasitic infection on human gut ecology and immune regulations. Transl. Med. Commun. 2021, 6, 1–9. [Google Scholar] [CrossRef]
- Ramírez-Carrillo, E.; Gaona, O.; Nieto, J.; Sánchez-Quinto, A.; Cerqueda-García, D.; Falcón, L.I.; Rojas-Ramos, O.A.; González-Santoyo, I. Disturbance in human gut microbiota networks by parasites and its implications in the incidence of depression. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Bancroft, A.J.; Hayes, K.S.; Grencis, R.K. Life on the edge: The balance between macrofauna, microflora and host immunity. Trends Parasitol. 2012, 28, 93–98. [Google Scholar] [CrossRef]
- Chalupowicz, L.; Manulis-Sasson, S.; Itkin, M.; Sacher, A.; Sessa, G.; Barash, I. Quorum-sensing system affects gall development incited by Pantoea agglomerans pv. gypsophilae. Mol. Plant-Microbe Interact. 2008, 21, 1094–1105. [Google Scholar] [CrossRef] [PubMed]
- Dutkiewicz, J.; Mackiewicz, B.; Lemieszek, M.K.; Golec, M.; Milanowski, J. Pantoea agglomerans: A mysterious bacterium of evil and good. Part I. Deleterious effects: Dust-borne endotoxins and allergens-focus on cotton dust. Ann. Agric. Environ. Med. 2015, 22, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Dutkiewicz, J.; Mackiewicz, B.; Lemieszek, M.K.; Golec, M.; Milanowski, J. Pantoea agglomerans: A mysterious bacterium of evil and good. Part III. Deleterious effects: Infections of humans, animals and plants. Ann. Agric. Environ. Med. 2016, 23, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Dutkiewicz, J.; Mackiewicz, B.; Lemieszek, M.K.; Golec, M.; Milanowski, J. Pantoea agglomerans: A mysterious bacterium of evil and good. Part II. Deleterious effects: Dust-borne endotoxins and allergens—Focus on grain dust, other agricultural dusts and wood dust. Ann. Agric. Environ. Med. 2016, 23, 110–133. [Google Scholar] [CrossRef]
- Kohchi, C.; Inagawa, H.; Nishizawa, T.; Yamaguchi, T.; Nagai, S.; Soma, G.I. Applications of lipopolysaccharide derived from Pantoea agglomerans (IP-PA1) for health care based on macrophage network theory. J. Biosci. Bioeng. 2016, 102, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef]
- Kundu, S.; Roy, S.; Lyndem, L.M. Cassia alata L: Potential role as anthelmintic agent against Hymenolepis diminuta. Parasitol. Res. 2012, 111, 1187–1192. [Google Scholar] [CrossRef]
- Awan, J.A.; Rahman, S.U. Microbiology Manual; Unitech Communications: Faisalabad, Pakistan, 2005; pp. 49–51. [Google Scholar]
- Mathialagan, M.; Edward, Y.S.J.T.; David, P.; Senthilkumar, M.; Srinivasan, M.; Mohankumar, S. Isolation, characterization and identification of probiotic lactic acid bacteria (LAB) from honey bees. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 849–906. [Google Scholar] [CrossRef]
- Powers, E.M. Efficacy of the Ryu nonstaining KOH technique for rapidly determining gram reactions of food-borne and waterborne bacteria and yeasts. Appl. Environ. Microbiol. 1995, 61, 3756–3758. [Google Scholar] [CrossRef]
- Prescott, L.M.; Harley, J.P. Laboratory Exercise in Microbiology, 5th ed.; McGraw-Hill Companies: New York, NY, USA, 2002; pp. 56–59. [Google Scholar]
- Liong, M.T.; Shah, N.P. Acid and bile tolerance and cholesterol removal ability of Lactobacilli strains. J. Dairy Sci. 2005, 88, 55–66. [Google Scholar] [CrossRef]
- Bergey, D.H.; Whitman, W.B.; De Vos, P.; Garrity, G.M.; Jones, D. Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Springer: London, UK; New York, NY, USA, 2009; Volume 3, pp. 477–510. [Google Scholar] [CrossRef]
- Chowdhury, A.; Hossain, M.N.; Mostazir, N.J.; Fakruddin, M.; Billah, M.; Ahmed, M. Screening of Lactobacillus spp. from buffalo yoghurt for probiotic and antibacterial activity. J. Bacteriol. Parasitol. 2012, 3, 156. [Google Scholar] [CrossRef]
- Jung, J.Y.; Han, S.S.; Kim, Z.H.; Kim, M.H.; Kang, H.K.; Jin, H.M.; Lee, M.H. In-Vitro Characterization of Growth Inhibition against the Gut Pathogen of Potentially Probiotic Lactic Acid Bacteria Strains Isolated from Fermented Products. Microorganisms 2021, 9, 2141. [Google Scholar] [CrossRef] [PubMed]
- Halder, D.; Mandal, M.; Chatterjee, S.S.; Pal, N.K.; Mandal, S. Indigenous probiotic Lactobacillus isolates presenting antibiotic like activity against human pathogenic bacteria. Biomedicines 2017, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Vlkova, E.; Rada, V.; Popelarova, P.; Trojanová, I.; Killer, J. Antimicrobial susceptibility of bifidobacterial isolated from gastrointestinal tract of calves. Livest. Sci. 2006, 105, 253–259. [Google Scholar] [CrossRef]
- Liasi, S.A.; Azmi, T.I.; Hassan, M.D.; Shuhaimi, M.; Rosfarizan, M. Antimicrobial activity and antibiotic sensitivity of three isolates of lactic acid bacteria from fermented fish product, Budu. Malays. J. Microbiol. 2009, 5, 33–37. [Google Scholar]
- Anderson, D.G.; Mckay, L.L. Simple and rapid method for isolating large plasmid DNA from lactic streptococci. Appl. Environ. Microbiol. 1983, 46, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Glendinning, L.; Nausch, N.; Free, A.; Taylor, D.W.; Mutapi, F. The microbiota and helminths: Sharing the same niche in the human host. Parasitology 2014, 141, 1255–1271. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Zhang, R.Q.; Zhai, Q.Y.; Liu, J.C.; Li, N.; Liu, W.X.; Li, L.; Shen, W. Metagenomic analysis of gut microbiota alteration in a mouse model exposed to mycotoxin deoxynivalenol. Toxicol. Appl. Pharmacol. 2019, 372, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Zaiss, M.M.; Harris, N.L. Interactions between the intestinal microbiome and helminth parasites. Parasite Immunol. 2016, 38, 5–11. [Google Scholar] [CrossRef]
- Zhang, D.; Hu, Q.; Liu, X.; Liu, X.; Gao, F.; Liang, Y.; Zou, K.; Su, Z.; Zhi, W.; Zhou, Z. A longitudinal study reveals the alterations of the Microtus fortis colonic microbiota during the natural resistance to Schistosoma japonicum infection. Exp. Parasitol. 2020, 219, 108030. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef]
- Portet, A.; Toulza, E.; Lokmer, A.; Huot, C.; Duval, D.; Galinier, R.; Gourbal, B. Experimental Infection of the Biomphalaria glabrata Vector Snail by Schistosoma mansoni Parasites Drives Snail Microbiota Dysbiosis. Microorganisms 2021, 9, 1084. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- McKenney, E.A.; Williamson, L.; Yoder, A.D.; Rawls, J.F.; Bilbo, S.D.; Parker, W. Alteration of the rat cecal microbiome during colonization with the helminth Hymenolepis diminuta. Gut Microbes 2015, 6, 182–193. [Google Scholar] [CrossRef]
- Wegener Parfrey, L.; Jirků, M.; Šíma, R.; Jalovecka, M.; Sak, B.; Grigore, K.; Jirků Pomajbíková, K. A benign helminth alters the host immune system and the gut microbiota in a rat model system. PLoS ONE 2017, 12, e0182205. [Google Scholar] [CrossRef]
- Boucard, A.S.; Thomas, M.; Lebon, W.; Polack, B.; Florent, I.; Langella, P.; Bermúdez-Humarán, L.G. Age and Giardia intestinalis Infection Impact Canine Gut Microbiota. Microorganisms 2021, 9, 1862. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, E.M.; Yi, M.H.; Lee, J.; Lee, S.; Hwang, Y.; Yong, D.; Sohn, W.M.; Yong, T.S. Chinese liver fluke Clonorchis sinensis infection changes the gut microbiome and increases probiotic Lactobacillus in mice. Parasitol. Res. 2019, 118, 693–699. [Google Scholar] [CrossRef]
- Rausch, S.; Held, J.; Fischer, A.; Heimesaat, M.M.; Kühl, A.A.; Bereswill, S.; Hartmann, S. Small intestinal nematode infection of mice is associated with increased enterobacterial loads alongside the intestinal tract. PLoS ONE 2013, 8, e74026. [Google Scholar] [CrossRef]
- Byerley, L.O.; Samuelson, D.; Blanchard IV, E.; Luo, M.; Lorenzen, B.N.; Banks, S.; Ponder, M.A.; Welsh, D.A.; Taylor, C.M. Changes in the gut microbial communities following addition of walnuts to the diet. J. Nutr. Biochem. 2017, 48, 94–102. [Google Scholar] [CrossRef]
- Lai, H.H.; Chiu, C.H.; Kong, M.S.; Chang, C.J.; Chen, C.C. Probiotic Lactobacillus casei: Effective for managing childhood diarrhea by altering gut microbiota and attenuating fecal inflammatory markers. Nutrients 2019, 11, 1150. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Vendrell, D.; de Blas, I.; Ruiz-Zarzuela, I.; Muzquiz, J.L.; Girones, O. Characterization of probiotic properties of lactic acid bacteria isolated from intestinal microbiota of fish. Aquaculture 2008, 278, 188–191. [Google Scholar] [CrossRef]
- Lähteinen, T.; Malinen, E.; Koort, J.M.; Mertaniemi-Hannus, U.; Hankimo, T.; Karikoski, N.; Pakkanen, S.; Laine, H.; Sillanpää, H.; Söderholm, H.; et al. Probiotic properties of Lactobacillus isolates originating from porcine intestine and feces. Anaerobe 2010, 16, 293–300. [Google Scholar] [CrossRef]
- Ripamonti, B.; Agazzi, A.; Bersani, C.; De Dea, P.; Pecorini, C.; Pirani, S.; Rebucci, R.; Savoini, G.; Stella, S.; Stenico, A.; et al. Screening of species-specific lactic acid bacteria for veal calves multi-strain probiotic adjuncts. Anaerobe 2011, 17, 97–105. [Google Scholar] [CrossRef]
- Succi, M.; Tremonte, P.; Reale, A.; Sorrentino, E.; Grazia, L.; Pacifico, S.; Coppola, R. Bile salt and acid tolerance of Lactobacillus rhamnosus strains isolated from Parmigiano Reggiano cheese. FEMS Microbiol. Lett. 2005, 244, 129–137. [Google Scholar] [CrossRef]
- Coulibaly, I.; Robin, D.D.; Destain, J.; Philippe, T. Characterization of Lactic Acid Bacteria Isolated from Poultry Farms in Senegal. Afr. J. Biotechnol. 2008, 7, 2006–2012. [Google Scholar] [CrossRef]
- Adnan, A.F.M.; Tan, I.K. Isolation of lactic acid bacteria from Malaysian foods and assessment of the isolates for industrial potential. Bioresour. Technol. 2007, 98, 1380–1385. [Google Scholar] [CrossRef]
- Bujnakova, D.; Strakova, E.; Kmet, V. In vitro evaluation of the safety and probiotic properties of Lactobacilli isolated from chicken and calves. Anaerobe 2014, 29, 118–127. [Google Scholar] [CrossRef]
- Pavli, F.; Argyri, A.; Papadopoulou, O.; Nychas, G.; Chorianopoulos, N.; Tassou, C. Probiotic potential of lactic acid bacteria from traditional fermented dairy and meat products: Assessment by in vitro tests and molecular characterization. J. Prob. Health 2016, 4, 157. [Google Scholar] [CrossRef]
- Mrázek, J.; Štrosová, L.; Fliegerova, K.; Kott, T.; Kopečný, J. Diversity of insect intestinal microflora. Folia Microbiol. 2008, 53, 229–233. [Google Scholar] [CrossRef]
- Moreira, J.L.S.; Mota, R.M.; Horta, M.F.; Teixeira, S.M.; Neumann, E.; Nicoli, J.R.; Nunes, Á.C. Identification to the species level of Lactobacillus isolated in probiotic prospecting studies of human, animal or food origin by 16S-23S rRNA restriction profiling. BMC Microbiol. 2005, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Su, X.; Zeng, B.; Yan, H.; Huang, Y.; Wang, E.; Yun, H.; Zhang, Y.; Liu, F.; Li, W.; et al. The gut epithelial receptor LRRC19 promotes the recruitment of immune cells and gut inflammation. Cell Rep. 2016, 14, 695–707. [Google Scholar] [CrossRef]
- Su, X.; Yan, H.; Huang, Y.; Yun, H.; Zeng, B.; Wang, E.; Liu, Y.; Zhang, Y.; Liu, F.; Che, Y.; et al. Expression of FABP4, adipsin and adiponectin in Paneth cells is modulated by gut Lactobacillus. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mulaw, G.; Sisay Tessema, T.; Muleta, D.; Tesfaye, A. In vitro evaluation of probiotic properties of lactic acid bacteria isolated from some traditionally fermented Ethiopian food products. Int. J. Microbiol. 2019, 2019, 7179514. [Google Scholar] [CrossRef]
- Cohen, M. Evaluation of probiotic properties of lactic acid bacteria isolated from some traditionally fermented food products. J. Med. Microbiol. Immunol. Res. 2021, 4, 16. [Google Scholar]
- Ekwi Damian, N.; Fossi, B.T.; Ngoune, L.T.; Tabe, E.S. Isolation, Molecular Identification and Phylogenetic Analysis of Bacteriocinogenic Lactic Acid Bacteria from Traditionally Fermented Foods from four Major Cities in Cameroon against Salmonella enterica pathogens. Int. J. Innov. Sci. Res. Technol. 2021, 6, 819–829. [Google Scholar]
- Reuter, G. The Lactobacillus and Bifidobacterium microflora of the human intestine: Composition and succession. Curr. Issues Intest. Microbiol. 2001, 2, 43–53. [Google Scholar]
- Čitar, M.; Hacin, B.; Tompa, G.; Štempelj, M.; Rogelj, I.; Dolinšek, J.; Narat, M.; Matijašić, B.B. Human intestinal mucosa-associated Lactobacillus and Bifidobacterium strains with probiotic properties modulate IL-10, IL-6 and IL-12 gene expression in THP-1 cells. Benef. Microbes 2015, 6, 325–336. [Google Scholar] [CrossRef]
- Ahmad, A.K.; Abdel-Hafeez, E.H.; Kamal, A.M. Some studies on spontaneous Hymenolepis diminuta infection in laboratory rats. J. Egypt. Soc. Parasitol. 2015, 45, 115–124. [Google Scholar] [CrossRef]
- Kapczuk, P.; Kosik-Bogacka, D.; Łanocha-Arendarczyk, N.; Gutowska, I.; Kupnicka, P.; Chlubek, D.; Baranowska-Bosiacka, I. Selected molecular mechanisms involved in the parasite–host system Hymenolepis diminuta–Rattus norvegicus. Int. J. Mol. Sci. 2018, 19, 2435. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Strain Code | KOH Test | Catalase Test | Spore Production | Curd Production Test | Gram Staining | Salt Tolerance (NaCl) | pH Tolerance | Bile Salt Tolerance Test | Hemolytic Activity | Antimicrobial Activity | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2% | 3% | 4.5% | 6% | 2.5 | 3.5 | 5.5 | 6.5 | 7 | ||||||||||
| CONTROL | S1 | - | - | - | + | Cocci + | + | + | - | - | - | - | - | + | - | γ | - | |
| S2 | - | - | - | - | Rod + | + | + | - | - | + | + | + | + | + | + | γ | - | |
| S3 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S4 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S5 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S6 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S7 | + | - | - | - | Rod- | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S8 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | + | γ | - | |
| S9 | - | + | - | - | Rod + | + | + | - | - | + | + | + | + | + | - | γ | - | |
| S10 | - | - | - | + | Rod + | + | + | + | + | + | + | + | + | + | + | γ | + | |
| S11 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S12 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S13 | - | - | - | + | Rod + | + | + | + | + | + | + | + | + | + | + | γ | + | |
| S14 | - | - | - | + | Rod + | + | + | + | + | + | + | + | + | + | + | γ | + | |
| S15 | - | - | - | - | Rod + | + | + | + | - | - | - | - | + | + | - | γ | - | |
| S16 | - | - | - | - | Rod + | - | - | - | - | - | - | - | + | + | - | γ | - | |
| S17 | - | - | - | + | Rod + | + | + | + | + | + | + | + | + | + | + | γ | + | |
| S18 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S19 | - | + | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S20 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S24 | - | - | - | - | Rod + | + | - | - | - | - | - | - | + | + | - | γ | - | |
| S25 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S26 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S27 | - | - | - | + | Rod + | + | + | + | + | + | + | + | + | + | + | γ | + | |
| S28 | - | - | - | - | Rod + | + | + | - | - | - | - | - | + | + | - | γ | - | |
| S29 | - | - | - | + | Rod + | + | + | + | + | + | + | + | + | + | + | γ | + | |
| INFECTION | S21 | - | - | - | - | Rod + | + | + | + | - | - | - | + | + | + | - | γ | - |
| S22 | - | - | - | - | Rod + | + | + | + | - | - | - | + | + | + | - | γ | - | |
| S23 | - | - | - | - | Rod + | + | + | + | - | - | - | + | + | + | - | γ | - | |
| Strain Code | ZDI (mm) of Antibiotic | |||||||
| AMP | MET | TR | AMC | PB | P | E | RIF | |
| S10 | 18 I | 19 I | 18 I | 21 S | 18 I | 22 S | 26 S | 21 S |
| S13 | 24 S | 16 I | 22 S | 24 S | 19 I | 18 I | 17 I | 16 I |
| S14 | 18 I | 16 I | 16 I | 19 I | 22 S | 26 S | 24 S | 24 S |
| S17 | 19 I | 19 I | 26 S | 18 I | 19 I | 26 S | 23 S | 24 S |
| S27 | 18 I | 20 I | 18 I | 19 I | 20 I | 19 I | 23 S | 24 S |
| S29 | 19 I | 16 I | 16 I | 19 I | 20 I | 23 S | 22 S | 19 S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mandal, S.; Mondal, C.; Mukherjee, T.; Saha, S.; Kundu, A.; Ghosh, S.; Lyndem, L.M. Hymenolepis diminuta Reduce Lactic Acid Bacterial Load and Induce Dysbiosis in the Early Infection of the Probiotic Colonization of Swiss Albino Rat. Microorganisms 2022, 10, 2328. https://doi.org/10.3390/microorganisms10122328
Mandal S, Mondal C, Mukherjee T, Saha S, Kundu A, Ghosh S, Lyndem LM. Hymenolepis diminuta Reduce Lactic Acid Bacterial Load and Induce Dysbiosis in the Early Infection of the Probiotic Colonization of Swiss Albino Rat. Microorganisms. 2022; 10(12):2328. https://doi.org/10.3390/microorganisms10122328
Chicago/Turabian StyleMandal, Sudeshna, Chandrani Mondal, Tanmoy Mukherjee, Samiparna Saha, Anirban Kundu, Sinchan Ghosh, and Larisha M. Lyndem. 2022. "Hymenolepis diminuta Reduce Lactic Acid Bacterial Load and Induce Dysbiosis in the Early Infection of the Probiotic Colonization of Swiss Albino Rat" Microorganisms 10, no. 12: 2328. https://doi.org/10.3390/microorganisms10122328
APA StyleMandal, S., Mondal, C., Mukherjee, T., Saha, S., Kundu, A., Ghosh, S., & Lyndem, L. M. (2022). Hymenolepis diminuta Reduce Lactic Acid Bacterial Load and Induce Dysbiosis in the Early Infection of the Probiotic Colonization of Swiss Albino Rat. Microorganisms, 10(12), 2328. https://doi.org/10.3390/microorganisms10122328