
Effects of Circadian Rhythm and Feeding Modes on Rumen Fermentation and Microorganisms in Hu Sheep



Abstract
1. Introduction
2. Materials and Methods
2.1. Animals Trial Design and Sample Collection
2.2. Sample Collection and Measurements
2.2.1. Rumen Fermentation Parameters
2.2.2. DNA Extraction and 16S rRNA Sequencing
2.2.3. Sequence Analysis
2.3. Microbial Data Analysis
3. Results
3.1. Rumen Fermentation Parameters
3.2. Rumen Digestive Enzyme Activity
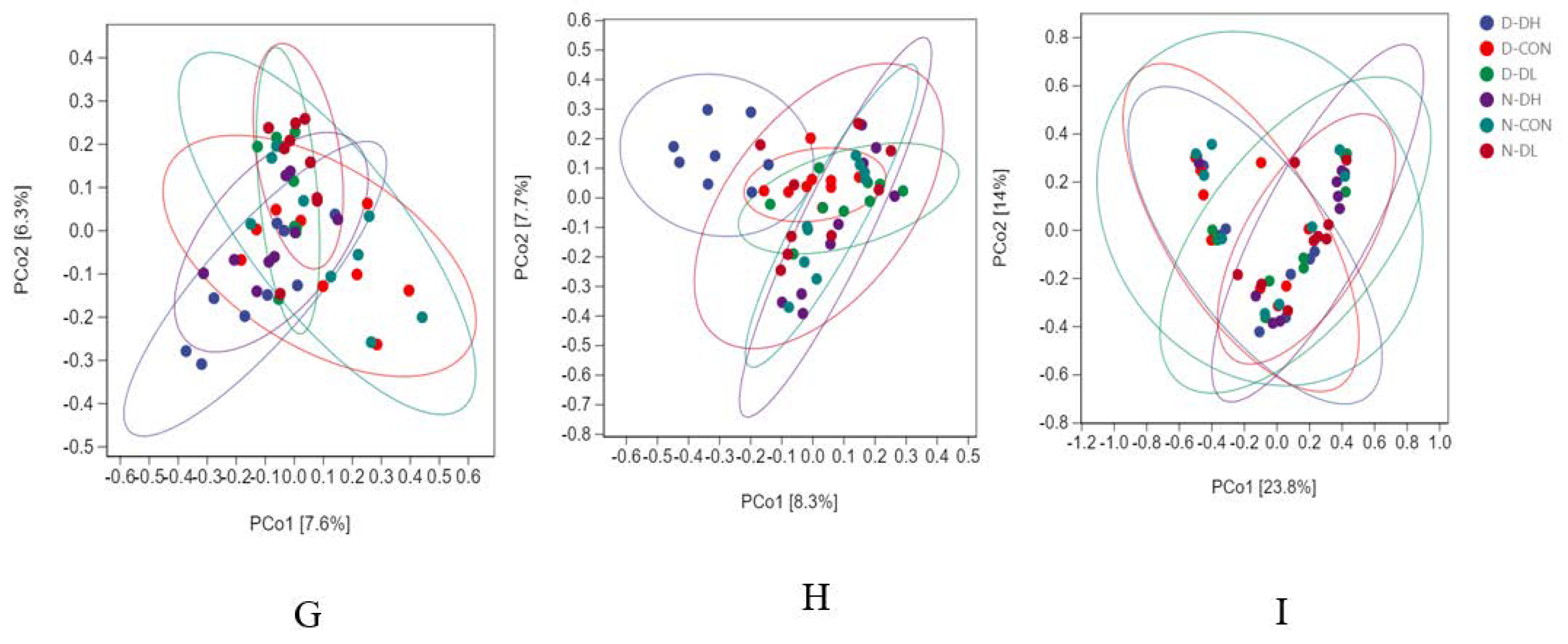
3.3. Rumen Microflora
4. Discussion
4.1. Circadian Rhythm Presented Effects on Rumen Environment, Enzyme Activity, and Rumen Microflora
4.1.1. Circadian Rhythm Presented Effects on Rumen Environment
4.1.2. Circadian Rhythm Presented Effects on Enzyme Activity
4.1.3. Effects of Circadian Rhythm on Rumen Microflora
4.2. Effects of Feeding Mode (Experimental Treatment) on Rumen Internal Environment, Enzyme Activity, and Rumen Microflora Based on Circadian Rhythm
4.2.1. Effects of Feeding Mode on Rumen Environment Based on Circadian Rhythm
4.2.2. Effects of Feeding Mode on Enzyme Activity Based on Circadian Rhythm
4.2.3. Effects of Feeding Mode on Rumen Microflora Based on Circadian Rhythm
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, G.; Wang, L.; Li, J.; Hou, R.; Wang, M.; Wang, Z.; Qu, Q.; Zhou, W.; Nie, Y.; Hu, Y.; et al. Seasonal shift of the gut microbiome synchronizes host peripheral circadian rhythm for physiological adaptation to a low-fat diet in the giant panda. Cell Rep. 2022, 38, 110203. [Google Scholar] [CrossRef]
- Honma, S. The mammalian circadian system: A hierarchical multi-oscillator structure for generating circadian rhythm. J. Physiol. Sci. 2018, 68, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Schapira, G.; Suez, J.; Tengeler, A.C.; Abramson, L.; Katz, M.N.; Korem, T.; Zmora, N.; et al. Transkingdom control of microbiota diurnal oscillations promotes metabolic homeostasis. Cell 2014, 159, 514–529. [Google Scholar] [CrossRef] [PubMed]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Verdouw, H.; Van Echteld, C.J.A.; Dekkers, E.M.J. Ammonia determination based on indophenol formation with sodium salicylate. Water Res. 1978, 12, 399–402. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef]
- Ramette, A. Multivariate analyses in microbial ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting Multivariate Models to Community Data: A Comment on Distance-Based Redundancy Analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Warton, D.I.; Wright, S.T.; Wang, Y. Distance-based multivariate analyses confound location and dispersion effects. Methods Ecol. Evol. 2012, 3, 89–101. [Google Scholar] [CrossRef]
- Huson, D.H.; Mitra, S.; Ruscheweyh, H.J.; Weber, N.; Schuster, S.C. Integrative analysis of environmental sequences using MEGAN4. Genome Res. 2011, 21, 1552–1560. [Google Scholar] [CrossRef] [PubMed]
- Asnicar, F.; Weingart, G.; Tickle, T.L.; Huttenhower, C.; Segata, N. Compact graphical representation of phylogenetic data and metadata with GraPhlAn. PeerJ 2015, 3, e1029. [Google Scholar] [CrossRef]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical methods for detecting differentially abundant features in clinical metagenomic samples. PLoS Comput. Biol. 2009, 5, e1000352. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Ehrlinger, J. ggRandomForests: Exploring Random Forest Survival. arXiv 2016, arXiv:1612.08974. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Sato, S.; Ikeda, A.; Tsuchiya, Y.; Ikuta, K.; Murayama, I.; Kanehira, M.; Okada, K.; Mizuguchi, H. A radio transmission pH measurement system for continuous evaluation of fluid pH in the rumen of cows. Vet. Res. Commun. 2012, 36, 85–89. [Google Scholar] [CrossRef]
- Gustafsson, A.H.; Palmquist, D.L. Diurnal Variation of Rumen Ammonia, Serum Urea, and Milk Urea in Dairy Cows at High and Low Yields. J. Dairy Sci. 1993, 76, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Z.; Hao, X.Y.; Xiang, B.W.; Zhang, W.J.; Zhang, H.X.; Zhang, J.X. Effects of Corn Husk and Soybean Husk Replacing Corn and Corn Straw on Growth Performance and Rumen Metabolism of Fattening Sheep. Chin. J. Anim. Nutr. 2020, 32, 765–774. [Google Scholar]
- Committee on Animal Nutrition, Board on Agriculture, National Research Council. Ruminant Nitrogen Usage; Animal Feed Science and Technology; National Academies Press: Washington, DC, USA, 1985. [Google Scholar]
- Yang, B.; Wang, J.; Yan, Y.N.; Aygul, A.; Zhang, J.S. Diurnal Dynamics of Rumen pH, Digestive Enzyme Activities and Protozoa Population in Dairy Cows. Chin. J. Anim. Nutr. 2021, 33, 1534–1544. [Google Scholar]
- Gharechahi, J.; Salekdeh, G.H. A metagenomic analysis of the camel rumen’s microbiome identifies the major microbes responsible for lignocellulose degradation and fermentation. Biotechnol. Biofuels 2018, 11, 216. [Google Scholar] [CrossRef]
- Goo, R.H.; Moore, J.G.; Greenberg, E.; Alazraki, N.P. Circadian variation in gastric emptying of meals in humans. Gastroenterology 1987, 93, 515–518. [Google Scholar] [CrossRef]
- Lindberg, G.; Iwarzon, M.; Hammarlund, B. 24-Hour Ambulatory Electrogastrography in Healthy Volunteers. Scand. J. Gastroenterol. 2009, 31, 658–664. [Google Scholar] [CrossRef]
- Liang, X.; Bushman, F.D.; Fitzgerald, G.A. Rhythmicity of the intestinal microbiota is regulated by gender and the host circadian clock. Proc. Natl. Acad. Sci. USA 2015, 112, 10479–10484. [Google Scholar] [CrossRef]
- Shaani, Y.; Zehavi, T.; Eyal, S.; Miron, J.; Mizrahi, I. Microbiome niche modification drives diurnal rumen community assembly, overpowering individual variability and diet effects. ISME J. 2018, 12, 2446–2457. [Google Scholar] [CrossRef]
- Da Silva, L.D.; Pereira, O.G.; Da Silva, T.C.; Valadares Filho, S.C.; Ribeiro, K.G. Effects of silage crop and dietary crude protein levels on digestibility, ruminal fermentation, nitrogen use efficiency, and performance of finishing beef cattle. Anim. Feed Sci. Technol. 2016, 220, 22–33. [Google Scholar] [CrossRef]
- Khafipour, E.; Krause, D.O.; Plaizier, J.C. A grain-based subacute ruminal acidosis challenge causes translocation of lipopolysaccharide and triggers inflammation. J. Dairy Sci. 2009, 92, 1060–1070. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Tunisa, M.; Wang, S.; Yang, H.; Yang, K. Effects of gossypol acetate on rumen microbial population, digestion and metabolism in sheep. Chin. J. Anim. Nutr. 2019, 31, 3421–3430. [Google Scholar]
- Wang, X.J.; Li, F.D.; Wang, S.Q.; Nian, F.; Li, C.; Wang, W.M.; Liu, T. Effects of neutral detergent Fiber level on tissue morphology and Digestive Enzyme Activities in digestive tract of lambs. Chin. J. Anim. Nutr. 2020, 32, 1704–1712. [Google Scholar]
- Sri, C.R.; Suhasini, K. Effect of Holistic Adoption of Dairy Farming Technologies on Households Farm Income of Small and Marginal Farmers: A Study of Telangana. Asian J. Agric. Ext. Econ. Sociol. 2020, 38, 114–122. [Google Scholar] [CrossRef]
- Wang, Z.; Elekwachi, C.; Jiao, J.; Wang, M.; Tang, S.; Zhou, C.; Tan, Z.; Forster, R.J. Changes in Metabolically Active Bacterial Community during Rumen Development, and Their Alteration by Rhubarb Root Powder Revealed by 16S rRNA Amplicon Sequencing. Front. Microbiol. 2017, 8, 159. [Google Scholar] [CrossRef]
- Bi, Y.; Zeng, S.; Zhang, R.; Diao, Q.; Tu, Y. Effects of dietary energy levels on rumen bacterial community composition in Holstein heifers under the same forage to concentrate ratio condition. BMC Microbiol. 2018, 18, 69. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef]
- Adamberg, K.; Kolk, K.; Jaagura, M.; Vilu, R.; Adamberg, S. The composition and metabolism of faecal microbiota is specifically modulated by different dietary polysaccharides and mucin: An isothermal microcalorimetry study. Benef. Microbes 2018, 9, 21–34. [Google Scholar] [CrossRef]
- Wallace, R.J.; McKain, N.; McEwan, N.R.; Miyagawa, E.; Chaudhary, L.C.; King, T.P.; Walker, N.D.; Apajalahti, J.H.; Newbold, C.J. Eubacterium pyruvativorans sp. nov., a novel non-saccharolytic anaerobe from the rumen that ferments pyruvate and amino acids, forms caproate and utilizes acetate and propionate. Int. J. Syst. Evol. Microbiol. 2003, 53, 965–970. [Google Scholar] [CrossRef]
- Varga, J.; Rigo, K.; Teren, J. Degradation of ochratoxin A by Aspergillus species. Int. J. Food Microbiol. 2000, 59, 1–7. [Google Scholar] [CrossRef]
- Sousa, D.O.; Oliveira, C.A.; Velasquez, A.V.; Souza, J.M.; Chevaux, E.; Mari, L.J.; Silva, L.F. Live yeast supplementation improves rumen fibre degradation in cattle grazing tropical pastures throughout the year. Anim. Feed Sci. Technol. 2018, 236, 149–158. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota Diurnal Rhythmicity Programs Host Transcriptome Oscillations. Cell 2016, 167, 1495–1510. [Google Scholar] [CrossRef] [PubMed]
- Bełżecki, G.; McEwan, N.R.; Kowalik, B.; Michałowski, T.; Miltko, R. Effect of Entodinium caudatum on starch intake and glycogen formation by Eudiplodinium maggii in the rumen and reticulum. Protistology 2017, 57, 38–49. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Ingredients | ||
|---|---|---|---|
| Concentrate (%) | Leymus Chinensis (%) | Concentrate + Leymus Chinensis (%) | |
| Dry matter, DM/(%) | 94.77 | 90.93 | 93.08 |
| Organics matter, OM/(%) | 88.93 | 93.26 | 90.84 |
| Metabolic energy, ME/(MJ/kg) | 11.52 | 5.93 | 9.06 |
| Crude protein, CP/(%) | 19.27 | 7.60 | 14.14 |
| Ether extract, EE/(%) | 3.43 | 2.45 | 3.00 |
| Neutral detergent fiber, NDF/(%) | 18.46 | 65.90 | 39.33 |
| Acid detergent fiber, ADF/(%) | 14.34 | 40.62 | 25.90 |
| Crude ash Ash/(%) | 11.07 | 6.74 | 9.16 |
| Calcium, Ca/(%) | 0.81 | 0.50 | 0.71 |
| Phosphorus, P/(%) | 0.49 | 0.08 | 0.31 |
| Groups | Diets | |
|---|---|---|
| Sunrise | Sunset | |
| CON | 50% total concentrate + 50% total roughage | 50% total concentrate + 50% total roughage |
| DH | 70% total concentrate + 30% total roughage | 30% total concentrate + 70% total roughage |
| DL | 30% total concentrate + 70% total roughage | 70% total concentrate + 30% total roughage |
| Items | Time Period | Groups | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| CON | DH | DL | Group | Time | Group ∗ Time | |||
| pH | Day | 6.84 a | 6.26 b | 7.02 a | 0.12 | <0.01 | <0.01 | 0.05 |
| Night | 6.30 a | 6.12 b | 6.38 a | |||||
| NH3-N/(mg·dL−1) | Day | 10.40 b | 17.91 a | 10.08 b | 0.75 | <0.01 | 0.01 | <0.01 |
| Night | 11.18 | 10.56 | 11.46 | |||||
| MCP/(mg·mL−1) | Day | 1.39 | 1.78 | 1.42 | 0.01 | 0.82 | 0.12 | 0.79 |
| Night | 1.09 | 1.08 | 1.11 | |||||
| Items | Time Period | Groups | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| CON | DH | DL | Group | Time | Group ∗ Time | |||
| Acetic acid/(%) | Day | 58.19 | 59.54 | 60.23 | 0.01 | 0.84 | <0.01 | <0.01 |
| Night | 61.17 | 57.69 | 56.29 | |||||
| Propionic acid/(%) | Day | 19.06 b | 26.76 a | 19.06 b | 0.23 | <0.01 | <0.01 | <0.01 |
| Night | 17.62 | 18.08 | 18.57 | |||||
| Isobutyric acid/(%) | Day | 12.50 ab | 12.45 a | 9.86 b | 0.13 | 0.05 | 0.63 | 0.46 |
| Night | 10.59 | 16.61 | 12.49 | |||||
| Butyric acid/(%) | Day | 8.76 a | 1.86 b | 9.55 a | 0.47 | <0.01 | 0.23 | 0.05 |
| Night | 8.57 a | 6.98 b | 9.23 a | |||||
| Isovaleric acid/(%) | Day | 0.83 | 0.81 | 0.78 | 0.03 | 0.06 | <0.01 | 0.11 |
| Night | 1.34 | 1.39 | 1.20 | |||||
| Valeric acid/(%) | Day | 0.66 b | 1.06 a | 0.52 b | 0.15 | 0.03 | 0.06 | <0.01 |
| Night | 0.74 | 063 | 0.63 | |||||
| TVFA/(mmol·L−1) | Day | 59.22 b | 81.31 a | 65.53 b | 0.34 | 0.47 | <0.01 | <0.01 |
| Night | 65.08 | 51.16 | 59.07 | |||||
| A/P | Day | 3.12 a | 2.52 b | 3.22 a | 0.03 | 0.005 | <0.01 | 0.55 |
| Night | 3.48 | 3.17 | 3.53 | |||||
| Items | Time | Groups | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| CON | DH | DL | Group | Time | Group ∗ Time | |||
| Amylase (U/g) | Day | 35.75 b | 44.63 a | 34.60 b | 0.32 | <0.01 | <0.01 | <0.01 |
| Night | 31.44 c | 33.47 b | 36.71 a | |||||
| Lipase (U/g) | Day | 61.42 b | 64.43 ab | 66.77 a | 0.30 | 0.06 | <0.01 | 0.05 |
| Night | 59.2 | 57.36 | 59.49 | |||||
| Cellulase (U/g) | Day | 4.43 | 4.44 | 4.85 | 0.15 | 0.06 | <0.01 | 0.42 |
| Night | 3.38 ab | 2.65 b | 3.66 a | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Ma, T.; Tu, Y.; Ma, S.; Diao, Q. Effects of Circadian Rhythm and Feeding Modes on Rumen Fermentation and Microorganisms in Hu Sheep. Microorganisms 2022, 10, 2308. https://doi.org/10.3390/microorganisms10122308
Zhang C, Ma T, Tu Y, Ma S, Diao Q. Effects of Circadian Rhythm and Feeding Modes on Rumen Fermentation and Microorganisms in Hu Sheep. Microorganisms. 2022; 10(12):2308. https://doi.org/10.3390/microorganisms10122308
Chicago/Turabian StyleZhang, Chuntao, Tao Ma, Yan Tu, Shulin Ma, and Qiyu Diao. 2022. "Effects of Circadian Rhythm and Feeding Modes on Rumen Fermentation and Microorganisms in Hu Sheep" Microorganisms 10, no. 12: 2308. https://doi.org/10.3390/microorganisms10122308
APA StyleZhang, C., Ma, T., Tu, Y., Ma, S., & Diao, Q. (2022). Effects of Circadian Rhythm and Feeding Modes on Rumen Fermentation and Microorganisms in Hu Sheep. Microorganisms, 10(12), 2308. https://doi.org/10.3390/microorganisms10122308