

Bioactive Compounds from Red Microalgae with Therapeutic and Nutritional Value

Abstract
1. General Overview of Red Microalgae
2. Red Microalgae with Potential to Be Used in Food and Health Industries
3. Valuable Biochemicals with Therapeutic and Nutritional Potential Produced by Red Microalgae
3.1. Polysaccharides
3.2. Pigments
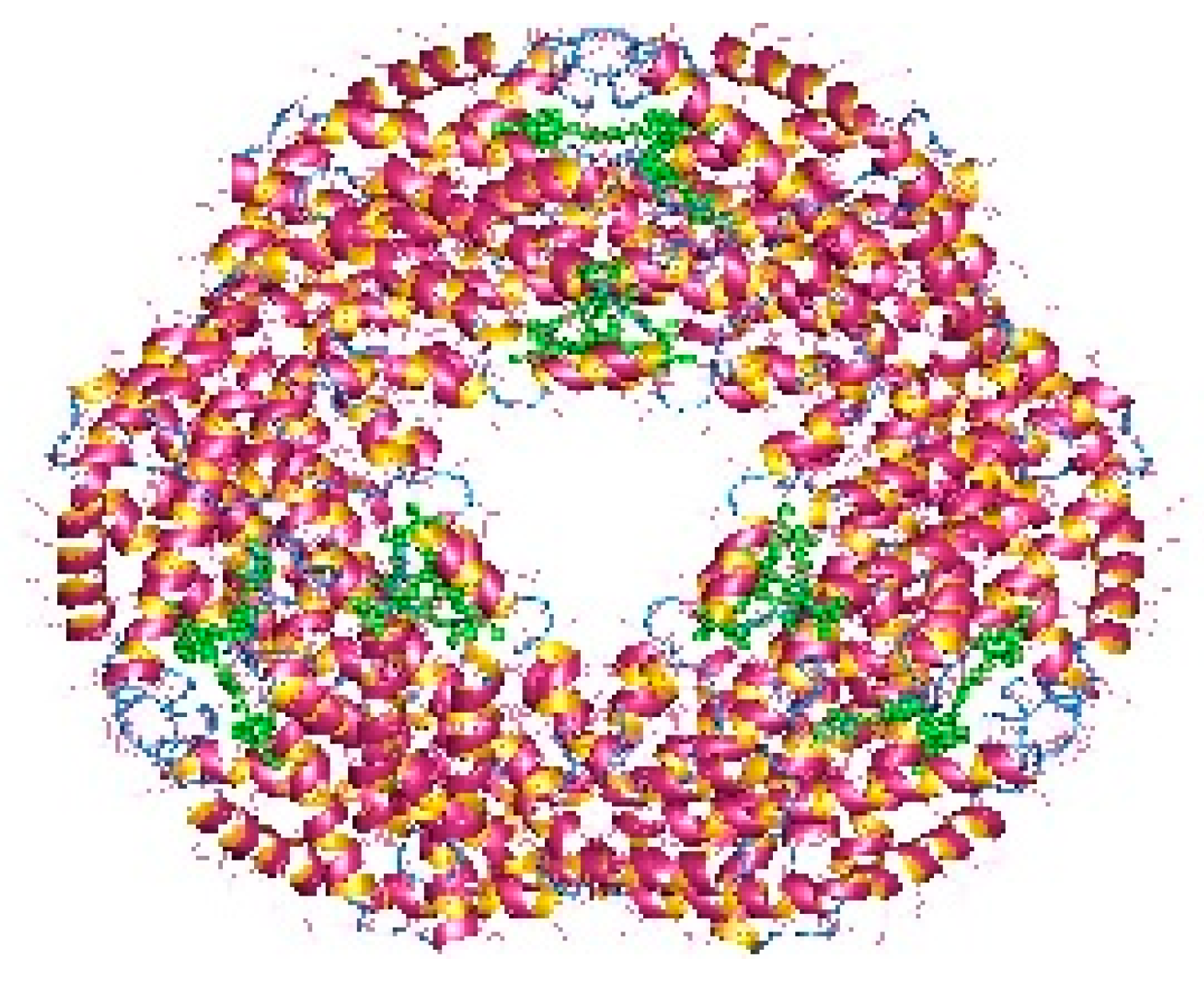
3.2.1. Phycobiliproteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Beneficial Substance | Application | Company/Product | Ref. |
|---|---|---|---|---|
| Porphyridium sp. | Pigments | Medical diagnostics, molecular biology | Greensea (Mèze, France) | [5] |
| Sulfated polysaccharides | Oxidative cell protection, immune photo-protection, anti-inflammatory, anti-irritation | Frutarom (Haifa, Israel) | [5] | |
| Living phytoplankton | Aquaculture | Greensea (Mèze, France) | [5] | |
| Oligosaccharides derived from EPS | Vascular maintenance, heavy leg syndrome relief, rosacea and redness inhibition | Silidine® by Greentech, St Beauzire, France | [39] | |
| sPS (sPS from marine bacteria and ulvan, associated with C-glycoside) | Improving the barrier function of skin, hydration | L’Oréal (Clichy, France) | [67] | |
| P. cruentum | Sulfated Polysaccharides | Cellular regeneration, moisturizing agent (cosmetics) | AlgoSource (Saint-Nazaire, France) | [5] |
| Antioxidant, anti-inflammatory, antimicrobial agent | Micoperi Blue Growth (Ortona, Italy) | [5] | ||
| Sun-shielding, anti-inflammatory, antiaging | Asta Technologies (Haryana, India) | [5] | ||
| Skin care | Solazyme (South San Francisco, CA, USA) | [5] | ||
| Oligosaccharide | Vasoconstriction of blood vessels | Greensea (Mèze, France) | [5] | |
| B-phycoerythrin | Medical diagnostics, molecular biology, fluorescence techniques | Phyco-Biotech (Montpellier, France) | [5] | |
| Phycobiliproteins | Medical diagnostics | Phyco-Biotech (Montpellier, France) | [5] | |
| Biomass | Aquaculture | Isua® Biotechnologie & Compagnie (Saint Just, France) | [5] | |
| P. purpureum P. cruentum | Exopolysaccharides | Antioxidant activity | Alguard® (Haifa, Israel) | [5] |
| P. purpureum | Phycoerythrin and EPS | Antiaging and sun care | Renouvellance® (Microphyt, France) | [39] |
| EPS | Melanin synthesis increase, enhances skin moisture and softness | Epsiline® (St Beauzire, France) | [39] | |
| EPS | Skin hydration | Hydrintense® (Vernier, Switzerland) | [39] | |
| Nondefined | Skin repair and hydration | algoVita (Tunis, Tunisia) | [39] | |
| Rhodella | Concentrated biomass | Improves skin condition | Detoxondria, CODIF technologie naturelle (Saint-Malo, France) | [39] |
| Rhodella violacea | Complete extract | Skin hydration | Rosacea, CODIF technologie naturelle (Saint-Malo, France) | [39] |
| Cyanidium caldarium | Nondefined | Reduces the signs of skin aging | TEGO® Stemlastin, (Essen, Germany) | [39] |
| Rhodosorus marinus | Nondefined | Soothes the nervous system | Mariliance, Givaudan (Vernier, Switzerland) | [39] |
3.2.2. Chlorophyll and Carotenoids
3.3. Polyunsaturated Fatty Acids
3.4. Micronutrients
4. Cultivation of Red Microalgae
| Strain | Nutrient Media, Conditions | Type of Photobioreactor | Product/Yield/ Result Achieved | Ref. |
|---|---|---|---|---|
| Porphyridium sp. UTEX 637 | ASW *; irradiance of 150 μmol photon m−2 s−1; aeration with air with 1–3% CO2; 24 ± 3 °C; cultivation time: stationary phase of growth | 1 L column Reactor | Enhanced antioxidant activity of PS | [51] |
| Porphyridium cruentum | ASW; blue light; PFD of 70 µE m−2 s−1; 25 °C; cultivation time: stationary phase of growth | Stirred airlift reactor | PS production yield of 0.95 g·L−1 | [108] |
| Porphyridium cruentum 2727 (strain UTEX 161) | Hemerick culture medium; 25 °C | 10 L tubular air-lift reactor | 6.0 mg/L EPS | [43] |
| Porphyridium cruentum (strain P.C-03) | Optimized OM Ι medium; PFD of 80 µE m−2 s−1; light–dark cycle of 18:6; 23–25 °C; cultivation time: 15 days | 15 L flat plate photobioreactor | Max growth rate of 0.32 day−1; max cell density of 137.9 × 108 L−1; PS production: 0.95 g·L−1 | [120] |
| Porphyridium cruentum 2727 Naegeli | Modified f/2 medium, final nitrate concentration of 8.82 × 10−3 M; final phosphate concentration of 3.62 × 10−4 M; PFD of 100 µmol m−2 s−1; 25 ± 1 °C | Attached cultivation column reactor | PS production of 42% dry weight | [121] |
| Porphyridium marinum 2727 CCAP 1380/10 | Pm medium; PFD of 360 μmol photons.m−2 s−1; 28 °C; cultivation time: 7 days after entering the stationary phase of growth | 5 L cylindrical, radially illuminated photobioreactor | EPS concentration of 2.5 g·L−1; EPS productivity of 0.149 g·L−1·day−1 | [122] |
| Porphyridium cruentum | Enriched ASW (2.8 gL−1 NaCl); 10.4 × 10−3 M KNO3 5.5 × 10−4 M KH2PO4 light intensity of 96 μmol m−2 s−1); 18 °C | 10 L tubular bioreactor | 415.88 ± 17.95 μg g−1 of β-carotene; 1513.12 ± 61.78 μg g−1 of chlorophyll a; specific growth rate of 0.70 d−1 | [123] |
5. Eco-Friendly Techniques for Extraction of Valuable Substances from Red Microalgae
6. Perspectives
- -
- Describing new species and strains with a better tolerance to environmental stress and an increased expression of valuable substances.
- -
- Applying new effective, selective, and low-cost methods for the separation of the desired compounds.
- -
- Developing new analytical and genetic tools for a better understanding of the chemical composition, biochemical pathways, and physiological processes in red microalgae.
- -
- Closing the gap between laboratory and large-scale production, keeping a steady ratio between different classes of the produced chemicals.
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rajvanshi, S.; Sharma, M.P. Microalgae: A potential source of biodiesel. J. Sustain. Bioenergy Syst. 2012, 2, 49–59. [Google Scholar] [CrossRef]
- Tesson, S.V.M.; Skjøth, C.A.; Šanti-Temkiv, T.; Löndahl, J. Airborne microalgae: Insights, opportunities, and challenges. Appl. Environ. Microbiol. 2016, 82, 1978–1991. [Google Scholar] [CrossRef] [PubMed]
- Pushkareva, E.; Johansen, J.R.; Elster, J. A review of the ecology, ecophysiology and biodiversity of microalgae in Arctic soil crusts. Polar Biol. 2016, 39, 2227–2240. [Google Scholar] [CrossRef]
- Arad, S.M.; Yaron, A. Natural pigments from red microalgae for use in foods and cosmetics. Trends Food Sci. Technol. 1992, 3, 92–97. [Google Scholar] [CrossRef]
- Gaignard, C.; Gargouch, N.; Dubessay, P.; Delattre, C.; Pierre, G.; Laroche, C.; Fendry, I.; Abdelkafi, S.; Micgaud, P. New horizons in culture and valorization of red microalgae. Biotechnol. Adv. 2019, 37, 193–222. [Google Scholar] [CrossRef] [PubMed]
- Matos, J.; Cardoso, C.; Bandarra, N.M.; Afonso, C. Microalgae as healthy ingredients for functional food: A review. Food Funct. 2017, 8, 2672–2685. [Google Scholar] [CrossRef]
- Hu, J.; Nagarajan, D.; Zhang, Q.; Chang, J.-S.; Lee, D.-J. Heterotrophic cultivation of microalgae for pigment production: A review. Biotechnol. Adv. 2018, 36, 54–67. [Google Scholar] [CrossRef]
- Yang, H.; Jin, X.; Lam, C.W.K.; Yan, S.-K. Oxidative stress and diabetes mellitus. Clin. Chem. Lab. Med. 2011, 49, 1773–1782. [Google Scholar] [CrossRef]
- Arad, S.M.; Levy-Ontman, O. Red microalgal cell-wall polysaccharides: Biotechnological aspects. Curr. Opin. Biotech. 2010, 21, 358–364. [Google Scholar] [CrossRef]
- Geresh, S.; Arad, S. The extracellular polysaccharides of the red microalgae: Chemistry and rheology. Bioresour. Technol. 1991, 38, 195–201. [Google Scholar] [CrossRef]
- Arad, S. Production of sulfated polysaccharides from red unicellular algae. In Algal Biotechnology; Stadler, T., Mollion, J., Verduset, M.C., Eds.; Elsevier Applied Science: London, UK, 1988; pp. 65–87. [Google Scholar]
- Castenholz, R.W.; McDermott, T.R. The Cyanidiales: Ecology, biodiversity, and biogeography. In Red Algae Genomic Age; Springer: Dordrecht, The Netherlands, 2010; pp. 357–371. ISBN 978-90-481-3794-7. [Google Scholar]
- Gloaguen, V.; Ruiz, G.; Morvan, H.; Mouradi-Givernaud, A.; Maes, E.; Krausz, P.; Strecker, G. The extracellular polysaccharide of Porphyridium sp.: An NMR study of lithium-resistant oligosaccharidic fragments. Carbohydr. Res. 2004, 339, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Aoki, M.; Ju, X.; Ueda, T.; Nakamura, Y.; Fujiwara, S.; Umemura, T.; Tsuzuki, M.; Minoda, A. Profiling of lipid and glycogen accumulations under different growth conditions in the sulfothermophilic red alga Galdieria sulphuraria. Bioresour. Technol. 2016, 200, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Varshney, P.; Mikulic, P.; Vonshak, A.; Beardall, J.; Wangikar, P.P. Extremophilic micro-algae and their potential contribution in biotechnology. Bioresour. Technol. 2015, 184, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Mobin, S.M.A.; Firoz, A. A review of microalgal biofuels, challenges and future directions. In Application of Thermo-Fluid Processes in Energy Systems: Key Issues and Recent Developments for a Sustainable Future, Singapore; Khan, M., Chowdhury, A., Hassan, N., Eds.; Springer Nature: Singapore, 2018; pp. 83–108. ISBN 978-981-10-0695-1. [Google Scholar]
- Chugh, M.; Kumar, L.; Shah, M.; Bharadvaja, N. Algal Bioremediation of heavy metals: An insight into removal mechanisms, recovery of by-products, challenges, and future opportunities. Energy Nexus 2022, 7, 100129. [Google Scholar] [CrossRef]
- Navarra, T. The Encyclopedia of Vitamins, Minerals and Supplements, 2nd ed.; Facts on File, Inc.: New York, NY, USA, 2004; ISBN 978-081604998. [Google Scholar]
- Metsoviti, M.N.; Katsoulas, N.; Karapanagiotidis, I.T.; Papapolymerou, G. Effect of nitrogen concentration, two-stage and prolonged cultivation on growth rate, lipid and protein content of Chlorella vulgaris. J. Chem. Technol. Biotechnol. 2019, 94, 1466–1473. [Google Scholar] [CrossRef]
- Mobin, S.M.A.; Firoz, A. Some promising microalgal species for commercial applications: A review. Energy Procedia 2017, 110, 510–517. [Google Scholar] [CrossRef]
- Rasala, B.A.; Mayfield, S.P. Photosynthetic biomanufacturing in green algae; production of recombinant proteins for industrial, nutritional, and medical uses. Photosynth. Res. 2015, 123, 227–239. [Google Scholar] [CrossRef]
- Wang, Y.; Tibbetts, S.M.; McGinn, P.J. Microalgae as sources of high-quality protein for human food and protein supplements. Foods 2021, 10, 3002. [Google Scholar] [CrossRef]
- Pignolet, O.; Jubeau, S.; Vaca-Garcia, C.; Michaud, P. Highly valuable microalgae: Biochemical and topological aspects. J. Ind. Microbiol. Biotechnol. 2013, 40, 781–796. [Google Scholar] [CrossRef]
- Ardiles, P.; Cerezal-Mezquita, P.; Salinas-Fuentes, F.; Órdenes, D.; Renato, G.; Ruiz-Domínguez, M. Biochemical composition and phycoerythrin extraction from red microalgae: A comparative study using green extraction technologies. Processes 2020, 8, 1628. [Google Scholar] [CrossRef]
- Huang, J. Analysis of the nutritional composition in Porphyridium cruentum. Food. Ferment. Ind. 2005, 31, 105–106. [Google Scholar]
- Borowitzka, M.A. Microalgae for aquaculture: Opportunities and constraints. J. Appl. Phycol. 1997, 9, 393–401. [Google Scholar] [CrossRef]
- Medina-Cabrera, E.V.; Rühmann, B.; Schmid, J.; Sieber, V. Characterization and comparison of Porphyridium sordidum and Porphyridium purpureum concerning growth characteristics and polysaccharide production. Algal Res. 2020, 49, 101931. [Google Scholar] [CrossRef]
- Schmidt, R.A.; Wiebe, M.G.; Eriksen, N.T. Heterotrophic high cell-density fed-batch cultures of the phycocyanin-producing red alga Galdieria sulphuraria. Biotechnol. Bioengin. 2005, 90, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Vítová, M.; Goecke, F.; Sigler, K.; Řezanka, T. Lipidomic analysis of the extremophilic red alga Galdieria sulphuraria in response to changes in pH. Algal Res. 2016, 13, 218–226. [Google Scholar] [CrossRef]
- Graziani, G.; Schiavo, S.; Nicolai, M.A.; Buono, S.; Fogliano, V.; Pinto, G.; Pollio, A. Microalgae as human food: Chemical and nutritional characteristics of the thermo-acidophilic microalga Galdieria sulphuraria. Food Funct. 2013, 4, 144–152. [Google Scholar] [CrossRef]
- Graverholt, O.V.; Eriksen, N.T. Heterotrophic high-cell density fed-batch and continuous-flow cultures of Galdieria sulphuraria and production of phycocyanin. Appl. Microbiol. Biotechnol. 2007, 77, 69–75. [Google Scholar] [CrossRef]
- Gavalás-Olea, A.; Siol, A.; Sakka, Y.; Köser, J.; Nentwig, N.; Hauser, T.; Filser, J.; Thöming, J.; Lang, I. Potential of the red alga Dixoniella grisea for the production of additives for lubricants. Plants 2021, 10, 1836. [Google Scholar] [CrossRef]
- Eggert, A.; Raimund, S.; Van Den Daele, K.; Karsten, U. Biochemical characterization of mannitol metabolism in the unicellular red alga Dixoniella grisea (Rhodellophyceae). Eur. J. Phycol. 2006, 41, 405–413. [Google Scholar] [CrossRef]
- Martínez-Miranda, J.G.; Chairez, I.; Durán-Páramo, E. Mannitol production by heterofermentative lactic acid bacteria: A review. Appl. Biochem. Biotechnol. 2022, 194, 2762–2795. [Google Scholar] [CrossRef]
- Becker, E. Microalgae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Schubert, N.; Garcia-Mendoza, E.; Pacheco-Ruiz, I. Carotenoid composition of marine red algae. J. Phycol. 2006, 42, 1208–1216. [Google Scholar] [CrossRef]
- Durmaz, Y.; Monteiro, M.; Bandarra, N.; Gökpinar, S.; Işik, O. The effect of low temperature on fatty acid composition and tocopherols of the red microalga, Porphyridium cruentum. J. Appl. Phycol. 2007, 19, 223–227. [Google Scholar] [CrossRef]
- Li, T.; Xu, J.; Wang, W.; Chen, Z.; Li, C.; Wu, H.; Wu, H.; Xiang, W. A novel three-step extraction strategy for high-value products from red algae Porphyridium purpureum. Foods 2021, 10, 2164. [Google Scholar] [CrossRef]
- Hachicha, R.; Elleuch, F.; Ben Hlima, H.; Dubessay, P.; de Baynast, H.; Delattre, C.; Pierre, G.; Hachicha, R.; Abdelkafi, S.; Michaud, P.; et al. Biomolecules from microalgae and cyanobacteria: Applications and market survey. Appl. Sci. 2022, 12, 1924. [Google Scholar] [CrossRef]
- Pierre, G.; Delattre, C.; Dubessay, P.; Jubeau, S.; Vialleix, C.; Cadoret, J.-P.; Probert, I.; Michaud, P. What is in store for EPS microalgae in the next decade? Molecules 2019, 24, 4296. [Google Scholar] [CrossRef]
- Banskota, A.H.; Sperker, S.; Stefanova, R.; McGinn, P.J.; O’Leary, S.J.B. Antioxidant properties and lipid composition of selected microalgae. J. Appl. Phycol. 2018, 31, 309–318. [Google Scholar] [CrossRef]
- Patel, A.K.; Laroche, C.; Marcati, A.; Ursu, A.V.; Jubeau, S.; Marchal, L.; Petit, E.; Djelveh, G.; Michaud, P. Separation and fractionation of exopolysaccharides from Porphyridium cruentum. Bioresour. Technol. 2013, 145, 345–350. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J. Appl. Phycol. 2015, 27, 1109–1119. [Google Scholar] [CrossRef]
- Barkallah, M.; Ben Atitallah, A.; Hentati, F.; Dammak, M.; Hadrich, B.; Fendri, I.; Ayadi, M.-A.; Michaud, P.; Abdelkafi, S. Effect of Spirulina platensis biomass with high polysaccharides content on quality attributes of common carp (Cyprinus carpio) and common barbel (Barbus barbus) fish burgers. Appl. Sci. 2019, 9, 2197. [Google Scholar] [CrossRef]
- Painter, T.J. Carbohydrate polymers in desert reclamation: The potential of microalgal biofertilizers. Carbohydr. Polym. 1993, 20, 77–86. [Google Scholar] [CrossRef]
- Arad, S.; Richmond, A. Industrial production of microalgal cell-mass and secondary products-species of high potential: Porphyridium sp. In Handbook of Microalgal Culture: Biotechnology & Applied Phycology; Richmond, A., Ed.; Blackwell Science: Carlton, Australia, 2004; pp. 289–297. [Google Scholar]
- Liberman, G.N.; Ochbaum, G.; Arad, S.M.; Bitton, R. The sulfated polysaccharide from a marine red microalga as a platform for the incorporation of zinc ions. Carbohydr. Polym. 2016, 152, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Liberman, G.N.; Ochbaum, G.; Mejubovsky-Mikhelis, M.; Ronit Bitton, R.; Arad, S.M. Physico-chemical characteristics of the sulfated polysaccharides of the red microalgae Dixoniella grisea and Porphyridium aerugineum. Int. J. Biol. Macromol. 2020, 145, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-N.; Li, Y.; Zhang, Y.; Xiang, W.; Li, A.F.; Li, T. Comparison on characterization and antioxidant activity of exopolysaccharides from two Porphyridium strains. J. Appl. Phycol. 2021, 33, 2983–2994. [Google Scholar] [CrossRef]
- Dvir, I.; Stark, A.H.; Chayoth, R.; Madar, Z.; Arad, S.M. Hypocholesterolemic effects of nutraceuticals produced from the red microalga Porphyridium sp. in rats. Nutrients 2009, 1, 156–167. [Google Scholar] [CrossRef]
- Tannin-Spitz, T.; Bergman, M.; van-Moppes, D.; Grossman, S.; Arad, S. Antioxidant activity of the polysaccharide of the red microalga Porphyridium sp. J. Appl. Phycol. 2005, 17, 215–222. [Google Scholar] [CrossRef]
- Setyaningsih, I.; Prasetyo, H.; Agungpriyono, D.R.; Tarman, K. Antihyperglycemic activity of Porphyridium cruentum biomass and extra-cellular polysaccharide in streptozotocin-induced diabetic rats. Int. J. Biol. Macromol. 2020, 156, 1381–1386. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, R.M.; de Morais, A.M.M.B. Health applications of bioactive compounds from marine microalgae. Life Sci. 2013, 93, 479–486. [Google Scholar] [CrossRef]
- Usov, A.I. Polysaccharides of the red algae. Adv. Carbohydr. Chem. Biochem. 2011, 65, 115–217. [Google Scholar] [CrossRef]
- Shimonaga, T.; Fujiwara, S.; Kaneko, M.; Izumo, A.; Nihei, S.; Francisco, P.B.; Satoh, A.; Fujita, N.; Nakamura, Y.; Tsuzuki, M. Variation in storage a-polyglucans of red algae: Amylose and semi-amylopectin types in Porphyridium and glycogen type in Cyanidium. Mar. Biotechnol. 2007, 9, 192–202. [Google Scholar] [CrossRef]
- Viola, R.; Nyvall, P.; Pedersen, M. The unique features of starch metabolism in red algae. Proc. R Soc. Lond. B 2001, 268, 1417–1422. [Google Scholar] [CrossRef] [PubMed]
- Gargouch, N.; Elleuch, F.; Karkouch, I.; Tabbene, O.; Pichon, C.; Gardarin, C.; Rihouey, C.; Picton, L.; Abdelkafi, S.; Fendri, I.; et al. Potential of exopolysaccharide from Porphyridium marinum to contend with bacterial proliferation, biofilm formation, and breast cancer. Mar. Drugs 2021, 19, 66. [Google Scholar] [CrossRef] [PubMed]
- Nagle, V.; Gaikwad, M.; Pawar, Y.; Dasgupta, S. Marine red alga Porphyridium sp. as a source of sulfated polysaccharides (SPs) for combating against COVID-19. biology, 2020; Preprints. [Google Scholar]
- Huang, J.; Chen, B.; You, W. Studies on separation of extracellular polysaccharide from Porphyridium cruentum and its anti-HBV activity in vitro. Chin. J. Drugs 2001, 6. [Google Scholar]
- De Jesus Raposo, M.F.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Influence of sulphate on the composition and antibacterial and antiviral properties of the exopolysaccharide from Porphyridium cruentum. Life Sci. 2014, 101, 56–63. [Google Scholar] [CrossRef]
- Huleihel, M.; Ishamu, V.; Tal, J.; Arad, S.M. Antiviral effect of red microalgal polysaccharides on Herpes simplex and Varicella zoster viruses. J. Appl. Phycol. 2001, 13, 127–134. [Google Scholar] [CrossRef]
- Pereira, L. Seaweeds as source of bioactive substances and skin care therapy—Cosmeceuticals, algotheraphy, and thalassotherapy. Cosmetics 2018, 5, 68. [Google Scholar] [CrossRef]
- Grassauer, A.; Prieschl-Grassauer, E. Antiviral Composition Comprising a Sulfated Polysaccharide. U.S. Patent No. 10,342,820, 9 July 2019. [Google Scholar]
- Dillon, H.F.; Somanchi, A.; Rao, K. Methods and Compositions for Cholesterol Reduction in Mammals. U.S. Patent No. US 20070167398A1, 19 January 2006. [Google Scholar]
- Yanhui, C. Cosmetic Composition with Functions of Repairing and Reinforcing Skin Barrier and Application Thereof. CN Patent No. CN 107412042, 17 June 2015. [Google Scholar]
- Bayona, K.C.D.; Gallon, S.M.N.; Estrada, A.L.; Rios, J.C.; Garces, L.A.; Martinez, A.M. Activity of sulfated polysaccharides from microalga Porphyridium cruentum over degenerative mechanisms of the skin. Int. J. Sci. Adv. Technol. 2012, 2, 85–92. [Google Scholar]
- Potter, A.; Ghibaudo, M.; Baltenneck, C. Association of Sulfated Polysaccharides and C-Glycoside and the Uses Thereof. WO Patent No. WO2014174188A1, 26 April 2013. [Google Scholar]
- Potter, A.; Thibaut, S.; Ribaut, C. Use of Sulphated Polysaccharides as Antidandruff Agent. WO Patent No. WO2013093307A1, 20 December 2011. [Google Scholar]
- Arad, S. Compositions Comprising Red Microalgae Polysaccharides and Metals. U.S. Patent No. US20110070159A1, 11 February 2014. [Google Scholar]
- Nimrod, A.; Greenman, B. Heavy Metal Salts of Hyaluronic Acid and Their Use as Antimicrobial Agents. U.S. Patent No. US4746504A, 24 May 1988. [Google Scholar]
- Arad, S.; Huliheil, M.; Tal, J. Antiviral Agents. WO Patent No. WO1997000689A1, 9 January 1997. [Google Scholar]
- Coragliotti, A.; Franklin, S.; Day, A.G.; Decker, S.M. Microalgal Polysaccharide. U.S. Patent No. US8927522B2, 6 January 2015. [Google Scholar]
- Pannard, A.; Pédrono, J.; Bormans, M.; Briand, E.; Claquin, P.; Lagadeue, Y. Production of exopolymers (EPS) by cyanobacteria: Impact on the carbon-to-nutrient ratio of the particulate organic matter. Aquat. Ecol. 2016, 50, 29–44. [Google Scholar] [CrossRef]
- Bermejo, R.R.; Alvarez-Pez, J.M.; Acien Fernandez, F.G.; Molina Grima, E. Recovery of pure B-phycoerythrin from the microalga Porphyridium cruentum. J. Biotechnol. 2002, 93, 73–85. [Google Scholar] [CrossRef]
- Wright, S.W.; Jeffrey, S.W. Pigment markers for phytoplankton production. In Marine Organic Matter: Biomarkers, Isotopes and DNA; The Handbook of Environmental Chemistry; Volkman, J.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 2N, ISBN 978-3-540-28401-7. [Google Scholar]
- Gantt, E.; Lipschultz, C.A. Phycobilisomes of Porphyridium cruentum. Pigment analysis. Biochemistry 1974, 13, 2960–2966. [Google Scholar] [CrossRef]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a commodity: Trends in applied research, patents and commercialization. J. Appl. Phycol. 2008, 20, 113–136. [Google Scholar] [CrossRef]
- Dvir, I.; Moppes, D.; Arad, S. Foodomics: To discover the health potential of microalgae. In Comprehensive Foodomics; Cifuentes, A., Ed.; Elsevier: Oxford, UK, 2021; pp. 658–671. [Google Scholar]
- Hsieh-Lo, M.; Castillo, G.; Ochoa-Becerra, M.A.; Mojica, L. Phycocyanin and phycoerythrin: Strategies to improve production yield and chemical stability. Algal Res. 2019, 42, 101600. [Google Scholar] [CrossRef]
- Richa, V.K.K.; Minu, K.I.G.; Rajeshwar, P.S. Biotechnological potentials of phycobiliproteins. Int. J. Pharma Bio Sci. 2011, 2, 1–9. [Google Scholar]
- Minkova, K.; Toshkova, R.; Gardeva, E.; Tchorbadjieva, M.; Ivanova, N.J.; Yossifova, L.; Gigova, L. Antitumor activity of B-phycoerythrin from Porphyridium cruentum. J. Pharm. Res. 2011, 4, 1480–1482. [Google Scholar]
- Pan, Q.; Chen, M.; Li, J.; Wu, Y.; Zhen, C.; Liang, B. Antitumor function and mechanism of phycoerythrin from Porphyra haitanensis. Biol. Res. 2013, 46, 87–95. [Google Scholar] [CrossRef]
- Glazer, A.N. Phycobiliproteins—A family of valuable, widely used fluorophores. J. Appl. Phycol. 1994, 6, 105–112. [Google Scholar] [CrossRef]
- Thoisen, C.; Hansen, B.W.; Nielsen, S.L. A simple and fast method for extraction and quantification of cryptophyte phycoerythrin. MethodsX 2017, 4, 209–213. [Google Scholar] [CrossRef]
- Ibanez-Gonzáleza, J.M.; Mazzuca-Sobczuka, T.; Redondo-Mirandaa, R.M.; Molina-Grimaa, E.; Cooney, C.L. A novel vortex flow reactor for the purification of B-phycoerythrin from Porphyridium cruentum. Chem. Eng. Res. Des. 2016, 111, 24–33. [Google Scholar] [CrossRef]
- Gross, W.; Schnarrenberger, C. Heterotrophic growth of two strains of the acido-thermophilic red alga Galdieria sulphuraria. Plant Cell Physiol. 1995, 36, 633–638. [Google Scholar] [CrossRef]
- Global Forecast to 2025, and Geography MRFB-104360; Meticulous Market Research Pvt. Ltd.: Maharashtra, India, 2020; p. 150.
- Kim, S.M.; Kang, S.W.; Kwon, O.N.; Chung, D.; Pan, C.H. Fucoxanthin as a major carotenoid in Isochrysis aff. galbana: Characterization of extraction for commercial application. J. Korean Soc. App. Biol. Chem. 2012, 55, 477–483. [Google Scholar] [CrossRef]
- Juin, C.; de Oliveira Junior, R.G.; Fleury, A.; Oudinet, C.; Pytowski, L.; Bérard, J.B.; Nicolau, E.; Thiéry, V.; Lanneluc, I.; Beaugeard, L.; et al. Zeaxanthin from Porphyridium purpureum induces apoptosis in human melanoma cells expressing the oncogenic BRAF V600E mutation and sensitizes them to the BRAF inhibitor vemurafenib. Braz. J. Pharmacogn. 2018, 28, 457–467. [Google Scholar] [CrossRef]
- Toker, O.S. Porphyridium cruentum as a natural colorant in chewing gum. Food Sci. Technol. 2019, 39, 195–201. [Google Scholar] [CrossRef]
- Gallego, R.; Martínez, M.; Cifuentes, A.; Ibáñez, E.; Herrero, M. Development of a green downstream process for the valorization of Porphyridium cruentum biomass. Molecules 2019, 24, 1564. [Google Scholar] [CrossRef] [PubMed]
- Morais, M.G.; Vaz, B.S.; Morais, E.G.; Costa, J.A.V. Biologically active metabolites synthesized by microalgae. BioMed Res. Int. 2015, 2015, 835761. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.; Mendiola, J.A.; Plaza, M.; Ibañez, E. Screening for bioactive compounds from algae. In Advanced Biofuels and Bioproducts; Lee, J., Ed.; Springer: New York, NY, USA, 2013; pp. 833–872. [Google Scholar]
- Fu, W.; David, R.N.; Zhiqian, Y.; Maonian, X.; Khraiwesh, B.; Jijakli, K.; Chaiboonchoe, A.; Alzahmi, A.; Al-Khairy, D.; Brynjolfsson, S.; et al. Bioactive compounds from microalgae: Current development and prospects. In Studies in Natural Products Chemistry; Elsevier B.V.: Cambridge, MA, USA, 2017; pp. 199–225. [Google Scholar]
- Raposo, M.F.; de Morais, A.M.; de Morais, R.M. Carotenoids from marine microalgae, a valuable natural source for the prevention of chronic diseases. Mar. Drugs 2015, 14, 5128–5155. [Google Scholar] [CrossRef] [PubMed]
- Gill, I.; Valivety, R. Polyunsaturated fatty acids, part 1: Occurrence, biological activities and applications. Trends Biotechnol. 1997, 15, 401–409. [Google Scholar] [CrossRef]
- Su, G.M.; Jiao, K.L.; Chang, J.Y.; Li, Z.; Guo, X.Y.; Sun, Y.; Zeng, X.H.; Lu, Y.H.; Lin, L. Enhancing total fatty acids and arachidonic acid production by the red microalgae Porphyridium purpureum. Bioresour. Bioproc. 2016, 3, 33. [Google Scholar] [CrossRef]
- Jiao, K.L.; Xiao, W.P.; Xu, Y.C.; Zeng, X.H.; Ho, S.-H.; Laws, E.A.; Lu, Y.; Ling, X.P.; Shi, T.; Sun, Y. Using a trait-based approach to optimize mixotrophic growth of the red microalga Porphyridium purpureum towards fatty acid production. Biotechnol. Biofuel. 2018, 11, 273. [Google Scholar] [CrossRef]
- Shanab, S.M.M.; Hafez, R.M.; Fouad, A.S. A review on algae and plants as potential source of arachidonic acid. J. Adv. Res. 2018, 11, 3–13. [Google Scholar] [CrossRef]
- Harwood, J.L.; Guschina, I.A. The versatility of algae and their lipid metabolism. Biochimie 2009, 91, 679–684. [Google Scholar] [CrossRef]
- Sato, N.; Moriyama, T.; Mori, N.; Toyoshima, M. Lipid metabolism and potentials of biofuel and high added-value oil production in red algae. World J. Microbiol. Biotechnol. 2017, 33, 74. [Google Scholar] [CrossRef] [PubMed]
- Vismara, R.; Vestir, S.; Kusmic, C.; Barsanti, L.; Gualtieri, P. Natural vitamin E enrichment of Artemia salina red freshwater and marine microalgae. J. Appl. Phycol. 2003, 15, 75–80. [Google Scholar] [CrossRef]
- Jialal, I.; Traber, M.; Devaraj, S. Is there a vitamin E paradox? Curr. Opin. Lipidol. 2001, 12, 49–53. [Google Scholar] [CrossRef]
- Ugwu, C.U.; Aoyagi, H.; Uchiyama, H. Photobioreactors for mass cultivation of algae. Bioresour. Technol. 2008, 99, 4021–4028. [Google Scholar] [CrossRef] [PubMed]
- Cezare-Gomes, E.A.; Mejia-da-Silva, L.D.C.; Pérez-Mora, L.S.; Matsudo, M.C.; Ferreira-Camargo, L.S.; Singh, K.A.; de Carvalho, J.C.M. Potential of microalgae carotenoids for industrial application. Appl. Biochem. Biotechnol. 2019, 188, 602–634. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.C.M.; Matsudo, M.C.; Bezerra, R.P.; Ferreira-Camargo, L.S.; Sato, S. Microalgae bioreactors. In Algal Biorefineries; Bajpai, R., Prokop, A., Zappi, M., Eds.; Springer: Amsterdam, The Netherlands, 2014; pp. 83–126. [Google Scholar]
- Jones, R.F.; Speer, H.L.; Kury, W. Studies on the growth of the red alga Porphyridium cruentum. Physiol. Plant. 2010, 16, 636–643. [Google Scholar] [CrossRef]
- You, T.; Barnett, S.M. Effect of light quality on production of extracellular polysaccharides and growth rate of Porphyridium cruentum. Biochem. Eng. J. 2004, 19, 251–258. [Google Scholar] [CrossRef]
- Ras, M.; Steyer, J.P.; Bernard, O. Temperature effect on microalgae: A crucial factor for outdoor production. Rev. Environ. Sci. Biotechnol. 2013, 12, 153–164. [Google Scholar] [CrossRef]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution. Annu. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Price, D.C.; Chan, C.X.; Qiu, H.; Rose, N.; Ball, S.; Weber, A.P.; Arias, M.C.; Henrissat, B.; Coutinho, P.M.; et al. Genome of the red alga Porphyridium purpureum. Nat. Commun. 2013, 4, 1941. [Google Scholar] [CrossRef]
- Masojidek, J.; Torzillo, G.; Kobližek, M. Photosynthesis in microalgae. In Handbook of Microalgal Culture, 2nd ed.; Richmond, A., Hu, Q., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Reeb, V.; Bhattacharya, D. The thermo-acidophilic cyanidiophyceae (cyanidiales). In Red Algae in the Genomic Age; Chapman, D., Seckbach, J., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 409–426. [Google Scholar] [CrossRef]
- Miller, S.R.; Castenholtz, R.W. Ecological physiology of Synechococcus sp. strain SH-94-5, a naturally occurring cyanobacterium deficient in nitrate assimilation. Appl. Environ. Microbiol. 2001, 67, 3002–3009. [Google Scholar] [CrossRef]
- Naval, J.; Antia, B.; Berland, R.; Bonin, D.J.; Maestrini, S.Y. Effects of urea concentration in supporting growth of certain marine microplanktonic algae. Phycologia 1977, 16, 105–111. [Google Scholar] [CrossRef]
- Li, X.; Li, W.; Zhai, J.; Wei, H. Effect of nitrogen limitation on biochemical composition and photosynthetic performance for fed-batch mixotrophic cultivation of microalga Spirulina platensis. Bioresour. Technol. 2018, 263, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Guihéneuf, F.; Stengel, D.B. Towards the biorefinery concept: Interaction of light, temperature and nitrogen for optimizing the co-production of high-value compounds in Porphyridium purpureum. Algal Res. 2015, 10, 152–163. [Google Scholar] [CrossRef]
- Hudson, J.J.; Taylor, W.D.; Schindler, D.W. Phosphate concentrations in lakes. Nature 2000, 406, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Z.; Zhuang, H.R.; Chen, B.L.; Shi, Q.Q.; Wu, S.G. Effect of organic substance on the growth of Porphyridium cruentum. Microbiology 2001, 28, 31–35. [Google Scholar]
- Liqin, S.; Wang, C.; Lei, S. Effects of light regime on extracellular polysaccharide production by Porphyridium cruentum cultured in flat plate photobioreactors. In Proceedings of the 2nd International Conference on Bioinformatics and Biomedical Engineering, Shanghai, China, 16–18 May 2008; pp. 1488–1491. [Google Scholar] [CrossRef]
- Lutzu, G.A.; Zhang, L.; Zhang, Z.; Liu, T. Feasibility of attached cultivation for polysaccharides production by Porphyridium cruentum. Bioprocess Biosyst. Eng. 2017, 40, 73–83. [Google Scholar] [CrossRef]
- Soanen, N.; Silva, E.D.; Gardarin, C.; Michaud, P.; Laroche, C. Improvement of exopolysaccharide production by Porphyridium marinum. Bioresour. Technol. 2016, 213, 231–238. [Google Scholar] [CrossRef]
- Durmaz, Y.; Tamtürk, F.; Konar, N.; Toker, O.; Palabiyik, I. Effect of pigment composition of Porphyridium cruentum as continuously culture method in industrial scale tubular photobioreactor. Int. J. Life Sci. Biotechnol. Pharma Res. 2017, 6, 18–21. [Google Scholar] [CrossRef]
- Lauceri, R.; Zittelli, G.C.; Torzillo, G.A. Simple method for rapid purification of phycobiliproteins from Arthrospira platensis and Porphyridium cruentum biomass. Algal Res. 2019, 44, 101685. [Google Scholar] [CrossRef]
- Munier, M.; Jubeau, S.; Wijaya, A.; Morancais, M.; Dumay, J.; Marchal, L.; Jaouen, P.; Fleurence, J. Physicochemical factors affecting the stability of two pigments: R-phycoerythrin of Grateloupia turuturu and B-phycoerythrin of Porphyridium cruentum. Food Chem. 2014, 150, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Schneedorferová, I.; Tomčala, A.; Valterová, I. Effect of heat treatment on the n-3/n-6 ratio and content of polyunsaturated fatty acids in fish tissues. Food Chem. 2015, 176, 205–211. [Google Scholar] [CrossRef]
- Barba, F.J.; Grimi, N.; Vorobiev, E. New approaches for the use of non-conventional cell disruption technologies to extract potential food additives and nutraceuticals from microalgae. Food Eng. Rev. 2015, 7, 45–62. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Application of novel extraction technologies for bioactives from marine algae. J. Agric. Food Chem. 2013, 61, 4667–4675. [Google Scholar] [CrossRef] [PubMed]
- Lang, Q.; Wai, C.M. Supercritical fluid extraction in herbal and natural product studies—A practical review. Talanta 2001, 53, 771–782. [Google Scholar] [CrossRef]
- Zougagh, M.; Valcárcel, M.; Rios, A. Supercritical fluid extraction: A critical review of its analytical usefulness TrAC. Trends Anal. Chem. 2004, 23, 399–405. [Google Scholar] [CrossRef]






| Species | Protein Content (% Dry Matter) | PS Content (% Dry Matter) | PUFA/Lipid Content | Carotenoids | Ref. |
|---|---|---|---|---|---|
| Porphyridium cruentum | 28–39 | >50 | 43.7% of total fatty acids | 19.11 ± 4.33 (mg g−1 extract) zeaxanthin; 43.15 ± 0.84 (mg g−1 extract) total carotenoids | [35,36,37] |
| Porphyridium purpureum | 15.08 | >50 | 40% of total fatty acids | 396.7 ± 0.3 µg g−1 dry weight β-carotene; 586.3 ± 0.3 µg g−1 dry weight zeaxanthin | [38,39,40] |
| Porphyridium aerugineum | 32 | No data | 5–9% of total fatty acids | 0.4 ± 0.01 mg g−1 dry weight zeaxanthin; 0.4 ± 0.07 mg g−1 dry weight β-carotene | [41,42,43] |
| Galdieria sulphuraria | 26–32 | 63–69 | Poor | 575 ± 123 mg kg−1 astaxanthin; 387 ± 112 mg kg−1 lutein | [29] |
| Dixoniella grisea | 10% cellular and 7% released | 56 | 4–5% of lipids | No phycobilisomes | [5,32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsvetanova, F.; Yankov, D. Bioactive Compounds from Red Microalgae with Therapeutic and Nutritional Value. Microorganisms 2022, 10, 2290. https://doi.org/10.3390/microorganisms10112290
Tsvetanova F, Yankov D. Bioactive Compounds from Red Microalgae with Therapeutic and Nutritional Value. Microorganisms. 2022; 10(11):2290. https://doi.org/10.3390/microorganisms10112290
Chicago/Turabian StyleTsvetanova, Flora, and Dragomir Yankov. 2022. "Bioactive Compounds from Red Microalgae with Therapeutic and Nutritional Value" Microorganisms 10, no. 11: 2290. https://doi.org/10.3390/microorganisms10112290
APA StyleTsvetanova, F., & Yankov, D. (2022). Bioactive Compounds from Red Microalgae with Therapeutic and Nutritional Value. Microorganisms, 10(11), 2290. https://doi.org/10.3390/microorganisms10112290