
Tilorone-Dihydrochloride Protects against Rift Valley Fever Virus Infection and Disease in the Mouse Model

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Compounds
2.3. In Vitro Virus Yield Reduction Assay
2.4. In Vitro Time of Addition Experiments
2.5. Mouse Efficacy Studies
2.6. Animal Ethics Statement
2.7. qRT-PCR
2.8. Histopathological Analysis
2.9. Statistical Analysis
3. Results
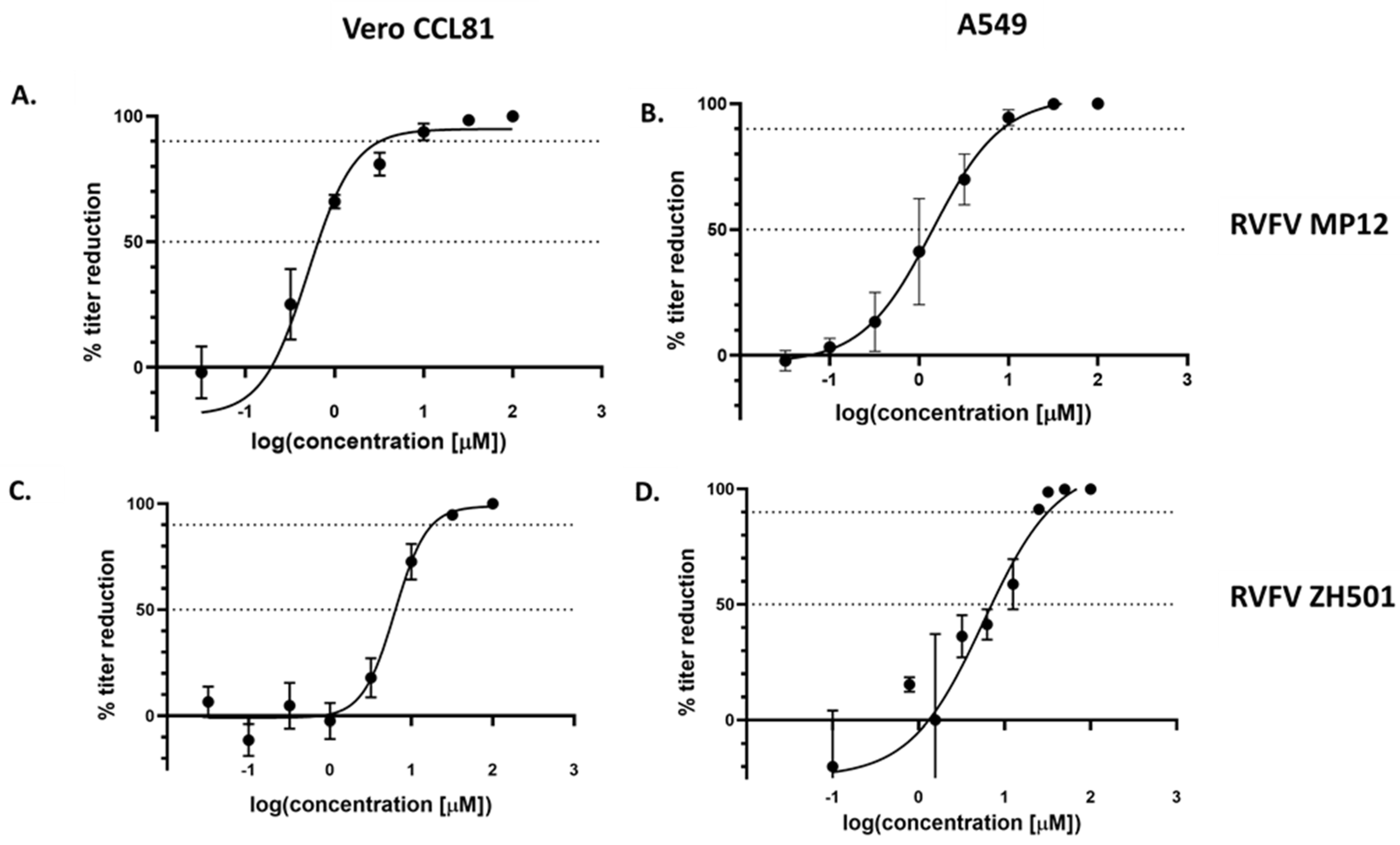
3.1. Tilorone Inhibits Rift Valley Fever Virus Replication In Vitro
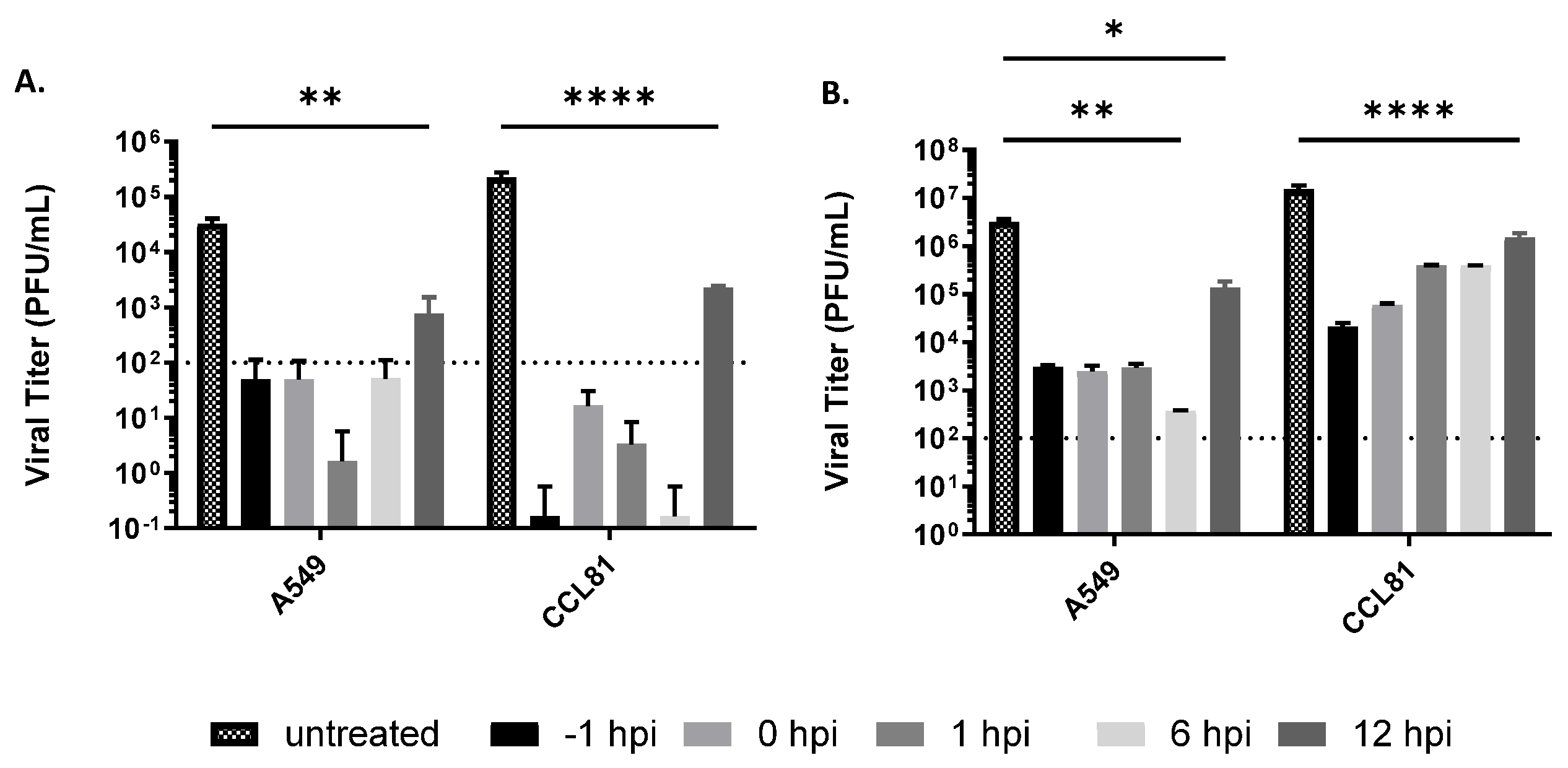
3.2. Delayed Treatment Efficacy of Tilorone on Rift Valley Fever Virus Infection In Vitro
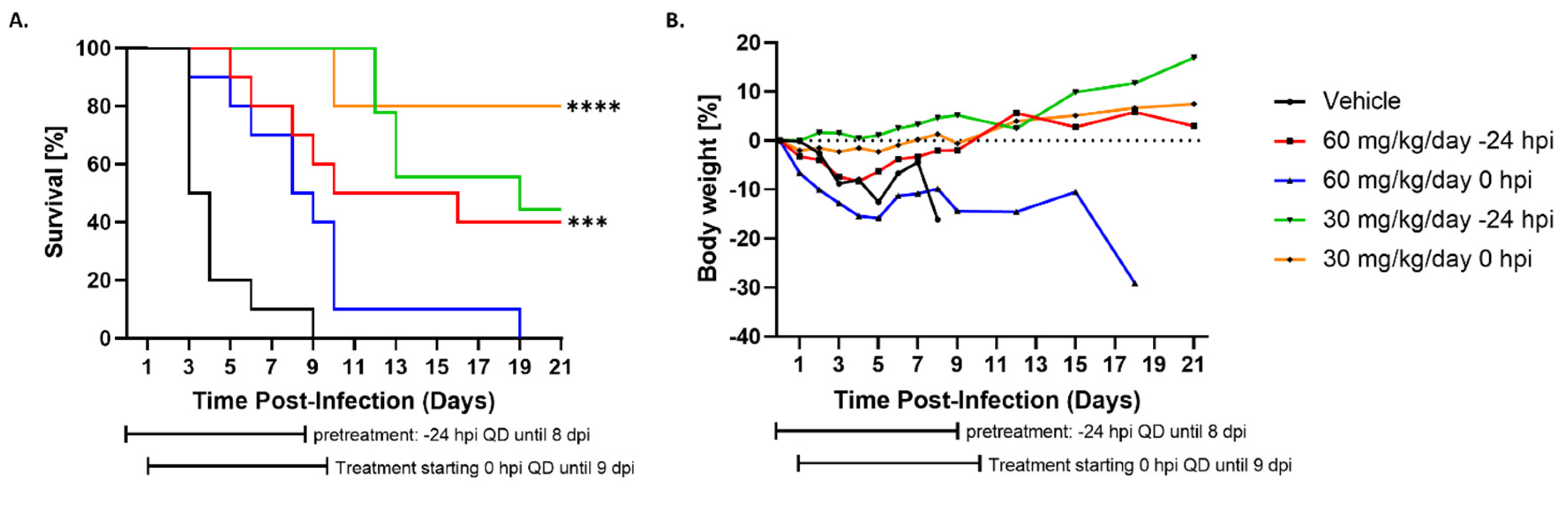
3.3. Administration of Tilorone Reduces RVFV-Induced Mortality in the BALB/c Mouse Model
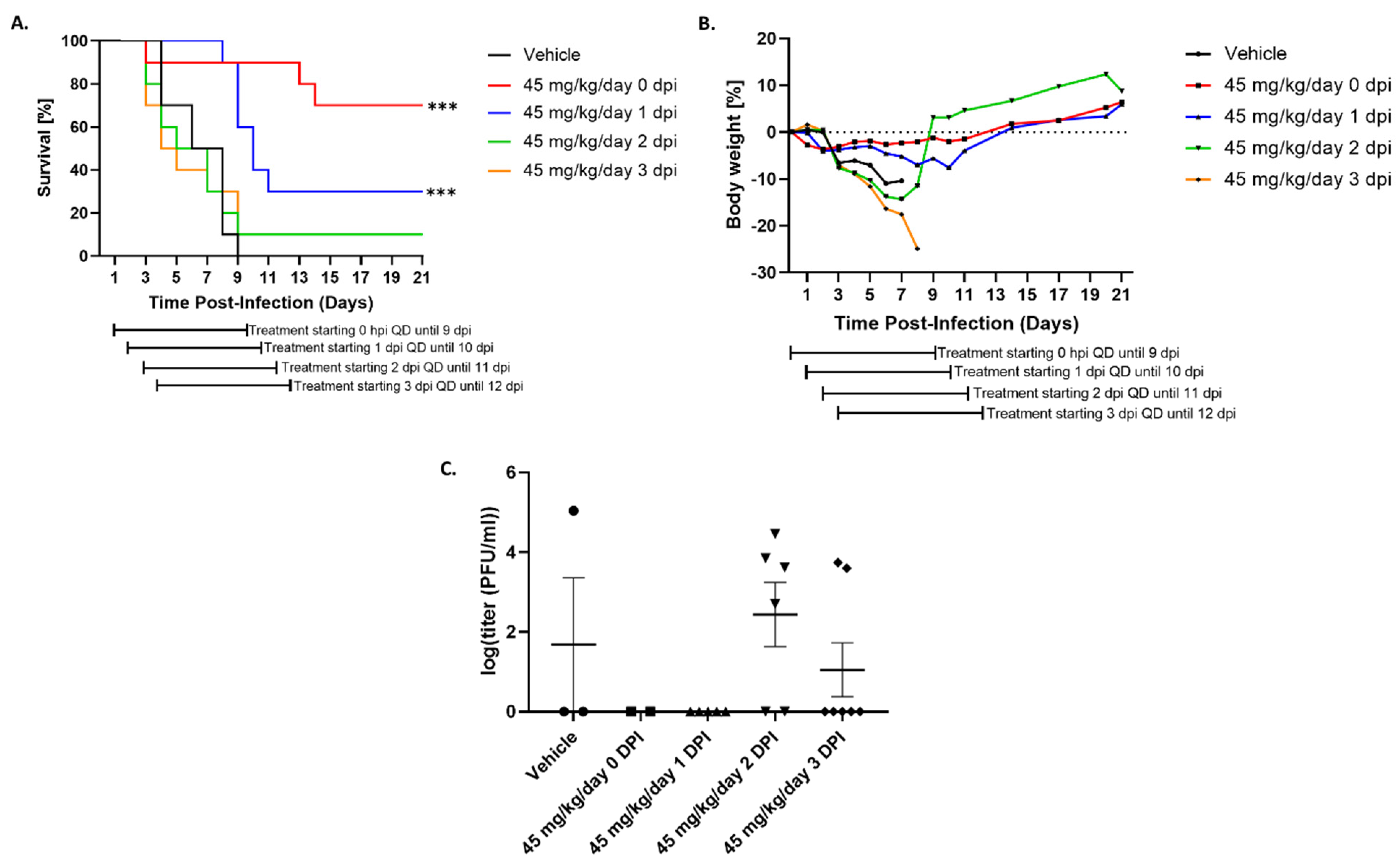
3.4. Tilorone Has Limited Window of Therapeutic Efficacy against Lethal RVFV Infection in BALB/c Mice
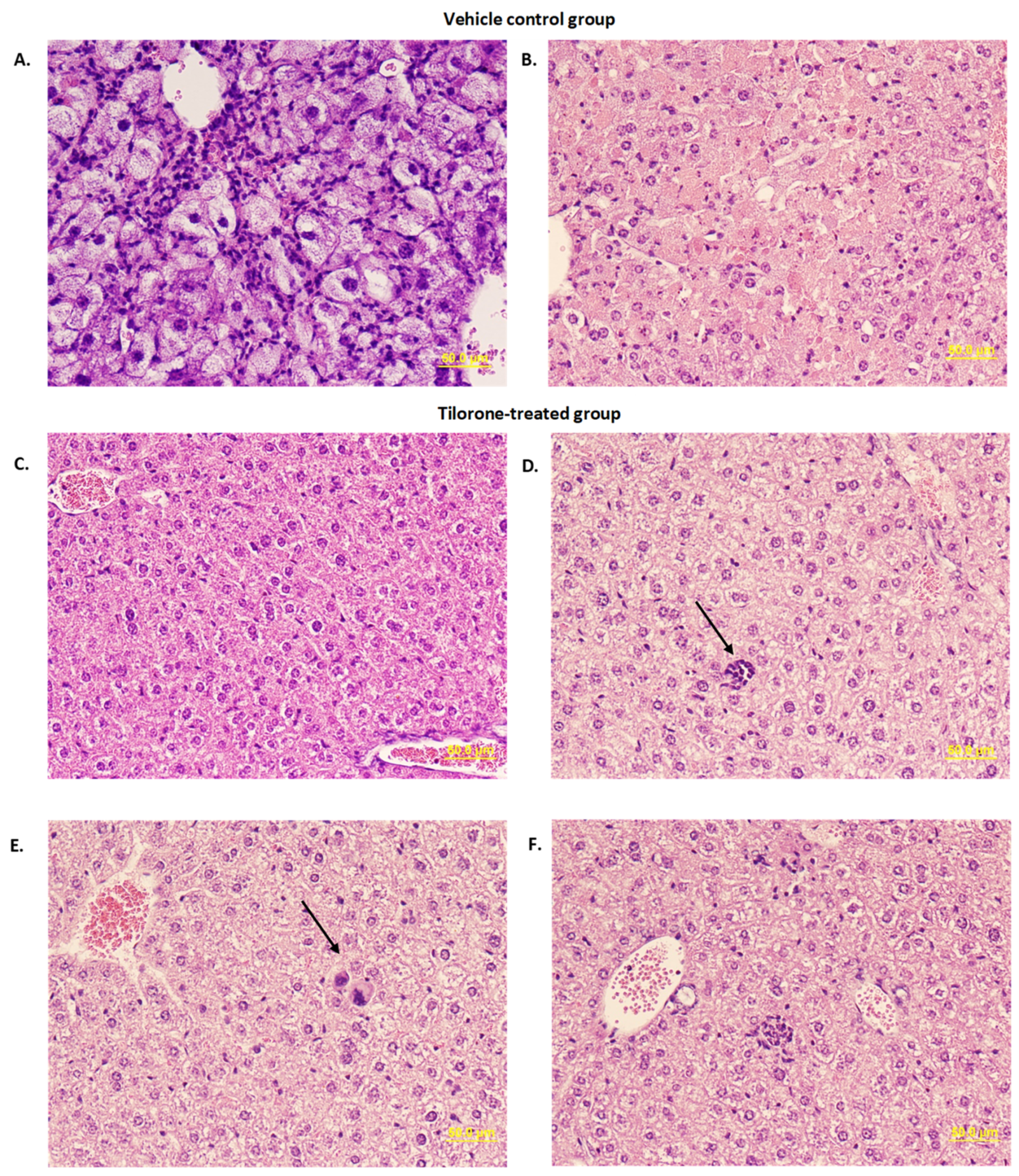
3.5. Histopathological Evaluation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daubney, R.; Hudson, J.R.; Garnham, P.C. Enzootic hepatitis or rift valley fever. An undescribed virus disease of sheep cattle and man from east africa. J. Pathol. Bacteriol. 1931, 34, 545–579. [Google Scholar] [CrossRef]
- Linthicum, K.J.; Britch, S.C.; Anyamba, A. Rift Valley Fever: An Emerging Mosquito-Borne Disease. Annu. Rev. Èntomol. 2016, 61, 395–415. [Google Scholar] [CrossRef] [PubMed]
- Hartman, A. Rift Valley Fever. Clin. Lab. Med. 2017, 37, 285–301. [Google Scholar] [CrossRef] [PubMed]
- Fafetine, J.M.; Coetzee, P.; Mubemba, B.; Nhambirre, O.; Neves, L.; Coetzer, J.A.; Venter, E.H. Rift Valley Fever Outbreak in Livestock, Mozambique, 2014. Emerg. Infect. Dis. 2016, 22, 2165–2167. [Google Scholar] [CrossRef] [Green Version]
- El Mamy, A.B.; Lô, M.M.; Thiongane, Y.; Diop, M.; Isselmou, K.; Doumbia, B.; Baba, M.O.; El Arbi, A.S.; Lancelot, R.; Kane, Y.; et al. Comprehensive Phylogenetic Reconstructions of Rift Valley Fever Virus: The 2010 Northern Mauritania Outbreak in the Camelus dromedarius Species. Vector-Borne Zoonotic Dis. 2014, 14, 856–861. [Google Scholar] [CrossRef] [Green Version]
- Sow, A.; Faye, O.; Ba, Y.; Ba, H.; Diallo, D.; Faye, O.; Loucoubar, C.; Boushab, M.; Barry, Y.; Diallo, M.; et al. Rift Valley Fever Outbreak, Southern Mauritania, 2012. Emerg. Infect. Dis. 2014, 20, 296–299. [Google Scholar] [CrossRef]
- Archer, B.N.; Weyer, J.; Paweska, J.; Nkosi, D.; Leman, P.; Tint, K.S.; Blumberg, L. Outbreak of Rift Valley fever affecting veterinarians and farmers in South Africa, 2008. S. Afr. Med. J. 2011, 101, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Jost, C.C.; Bett, B.; Mariner, J.C.; Kihu, S.; Swai, E.S.; Njogu, G.; Nzietchueng, S. Epidemiological Assessment of the Rift Valley Fever Outbreak in Kenya and Tanzania in 2006 and 2007. Am. J. Trop. Med. Hyg. 2010, 83, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Kahlon, S.S.; Peters, C.J.; LeDuc, J.; Muchiri, E.M.; Muiruri, S.; Njenga, M.K.; Breiman, R.F.; White, A.C.; King, C.H. Severe Rift Valley Fever May Present with a Characteristic Clinical Syndrome. Am. J. Trop. Med. Hyg. 2010, 82, 371–375. [Google Scholar] [CrossRef] [Green Version]
- Madani, T.A.; Al-Mazrou, Y.Y.; Al-Jeffri, M.H.; Mishkhas, A.A.; Al-Rabeah, A.M.; Turkistani, A.M.; Al-Sayed, M.O.; Abodahish, A.A.; Khan, A.S.; Ksiazek, T.G.; et al. Rift Valley Fever Epidemic in Saudi Arabia: Epidemiological, Clinical, and Laboratory Characteristics. Clin. Infect. Dis. 2003, 37, 1084–1092. [Google Scholar] [CrossRef] [Green Version]
- Strausbaugh, L.J.; Laughlin, L.W.; Meegan, J.M.; Watten, R.H. Clinical studies on Rift Valley fever, Part I: Acute febrile and hemorrhagic-like diseases. J. Egypt. Public Health Assoc. 1978, 53, 181–182. [Google Scholar]
- Al-Hazmi, M.; Ayoola, E.A.; Abdurahman, M.; Banzal, S.; Ashraf, J.; El-Bushra, A.; Hazmi, A.; Abdullah, M.; Abbo, H.; Elamin, A.; et al. Epidemic Rift Valley Fever in Saudi Arabia: A Clinical Study of Severe Illness in Humans. Clin. Infect. Dis. 2003, 36, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Bron, G.; Strimbu, K.; Cecilia, H.; Lerch, A.; Moore, S.; Tran, Q.; Perkins, T.; Bosch, Q.T. Over 100 Years of Rift Valley Fever: A Patchwork of Data on Pathogen Spread and Spillover. Pathogens 2021, 10, 708. [Google Scholar] [CrossRef] [PubMed]
- NIAID Emerging Infectious Diseases/Pathogens. 2018. Available online: https://www.niaid.nih.gov/research/emerging-infectious-diseases-pathogens (accessed on 30 August 2021).
- Rolin, A.I.; Berrang-Ford, L.; Kulkarni, M.A. The risk of Rift Valley fever virus introduction and establishment in the United States and European Union. Emerg. Microbes Infect. 2013, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.T.; Hill, D.R.; Solomon, T.; Aronson, N.; Endy, T.P. 37-Viral Hemorrhagic Fevers. In Hunter’s Tropical Medicine and Emerging Infectious Diseases, 10th ed.; Hill, D.R., Ryan, E.R., Aronson, N., Endy, T.P., Solomon, T., Eds.; Elsevier Health Sciences: London, UK, 2020; Volume 37, pp. 351–381. [Google Scholar]
- Atkins, C.; Freiberg, A.N. Recent advances in the development of antiviral therapeutics for Rift Valley fever virus infection. Future Virol. 2017, 12, 651–665. [Google Scholar] [CrossRef]
- Ekins, S.; Lane, T.R.; Madrid, P.B. Tilorone: A broad-spectrum antiviral invented in the USA and commercialized in Russia and beyond. Pharm. Res. 2020, 37, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chizhov, N.P.; Smol’Skaia, T.T.; Baĭchenko, P.I.; Luk’Ianova, R.I.; Teslenko, V.M.; Bavra, G.P.; Romanchenko, I.A.; Shatalov, E.B.; Ershov, F.I.; Tazulakhova, E.B. Clinical research on the tolerance for and the interferon-inducing activity of amiksin. Vopr. Virusol. 1990, 35, 411–414. [Google Scholar] [PubMed]
- Sel’Kova, E.P.; Semenenko, T.A.; Nosik, N.N.; Iudina, T.I.; Amarian, M.P.; Lavrukhina, L.A.; Pantiukhova, T.N.; Tarasova, G.I. Effect of amyxin—A domestic analog of tilorone—On characteristics of interferon and immune status of man. Zhurnal Mikrobiol. Epidemiol. I Immunobiol. 2001, 2001, 31–35. [Google Scholar]
- Sel’Kova, E.P.; Iakovlev, V.N.; A Semenenko, T.A.; Filatov, N.; Gotvianskaia, T.P.; Danilina, G.A.; Pantiukhova, T.N.; Nikitina, G.I.; Tur’Ianov, M.K. Evaluation of amyxin effect in prophylaxis of acute respiratory viral infections. Zhurnal Mikrobiol. Epidemiol. I Immunobiol. 2001, 2001, 42–46. [Google Scholar]
- A Semenenko, T.; Selkova, E.P.; Nikitina, G.Y.; Gotvyanskaya, T.P.; I Yudina, T.; Amaryan, M.P.; Nosik, N.N.; Turyanov, M.H. Immunomodulators in the prevention of acute respiratory viral infections. Russ. J. Immunol. RJI Off. J. Russ. Soc. Immunol. 2002, 7, 105–114. [Google Scholar]
- Zakirov, I.G. Use of amixin in the therapy and prevention of some viral infections. Klin. Med. 2002, 80, 54–56. [Google Scholar]
- Ekins, S.; Lingerfelt, M.A.; Comer, J.E.; Freiberg, A.N.; Mirsalis, J.C.; O’Loughlin, K.; Harutyunyan, A.; McFarlane, C.; Green, C.E.; Madrid, P.B. Efficacy of Tilorone Dihydrochloride against Ebola Virus Infection. Antimicrob. Agents Chemother. 2018, 62, e01711-17. [Google Scholar] [CrossRef] [Green Version]
- Kuehne, R.W.; Pannier, W.L.; Stephen, E.L. Evaluation of Various Analogues of Tilorone Hydrochloride Against Venezuelan Equine Encephalitis Virus in Mice. Antimicrob. Agents Chemother. 1977, 11, 92–97. [Google Scholar] [CrossRef] [Green Version]
- Vargin, V.V.; Zschiesche, W.; Semenov, B.F. Effects of tilorone hydrochloride on experimental flavivirus infections in mice. Acta Virol. 1977, 21, 114–118. [Google Scholar]
- Puhl, A.C.; Fritch, E.J.; Lane, T.R.; Tse, L.V.; Yount, B.L.; Sacramento, C.Q.; Fintelman-Rodrigues, N.; Tavella, T.A.; Costa, F.T.M.; Weston, S.; et al. Repurposing the Ebola and Marburg Virus Inhibitors Tilorone, Quinacrine, and Pyronaridine: In Vitro Activity against SARS-CoV-2 and Potential Mechanisms. ACS Omega 2021, 6, 7454–7468. [Google Scholar] [CrossRef]
- Krueger, R.F.; Mayer, G.D. Tilorone Hydrochloride: An Orally Active Antiviral Agent. Science 1970, 169, 1213–1214. [Google Scholar] [CrossRef]
- Lane, T.R.; Ekins, S. Toward the Target: Tilorone, Quinacrine, and Pyronaridine Bind to Ebola Virus Glycoprotein. ACS Med. Chem. Lett. 2020, 11, 1653–1658. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.; Ashshi, A.; Asghar, A.; El-Rahim, I.A.; El-Shemi, A.; Zafar, T. Seroepidemiological survey on Rift Valley fever among small ruminants and their close human contacts in Makkah, Saudi Arabia, in 2011. Rev. Sci. Tech. L’oie 2014, 33, 903–915. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Update: Outbreak of Rift Valley Fever—Saudi Arabia, August-November 2000. MMWR. Morb. Mortal. Wkly. Rep. 2000, 49, 982–985. [Google Scholar]
- Jeanmaire, E.M.; Rabenarivahiny, R.; Biarmann, M.; Rabibisoa, L.; Ravaomanana, F.; Randriamparany, T.; Andriamandimby, S.F.; Diaw, C.S.; Fenozara, P.; de La Rocque, S.; et al. Prevalence of Rift Valley Fever Infection in Ruminants in Madagascar After the 2008 Outbreak. Vector-Borne Zoonotic Dis. 2011, 11, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Sissoko, D.; Giry, C.; Gabrie, P.; Tarantola, A.; Pettinelli, F.; Collet, L.; D’Ortenzio, E.; Renault, P.; Pierre, V. Rift Valley Fever, Mayotte, 2007–2008. Emerg. Infect. Dis. 2009, 15, 568–570. [Google Scholar] [CrossRef]
- Simons, R.R.L.; Croft, S.; Rees, E.; Tearne, O.; Arnold, M.E.; Johnson, N. Using species distribution models to predict potential hot-spots for Rift Valley Fever establishment in the United Kingdom. PLoS ONE 2019, 14, e0225250. [Google Scholar] [CrossRef]
- Chevalier, V.; Pepin, M.; Plée, L.; Lancelot, R. Rift Valley fever—A threat for Europe? Eurosurveillance 2010, 15, 19506. [Google Scholar] [CrossRef]
- Caroline, A.L.; Powell, D.S.; Bethel, L.M.; Oury, T.D.; Reed, D.; Hartman, A.L. Broad Spectrum Antiviral Activity of Favipiravir (T-705): Protection from Highly Lethal Inhalational Rift Valley Fever. PLoS Negl. Trop. Dis. 2014, 8, e2790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, C.; Lin, K.; Wilhelmsen, C.; Friedrich, B.; Nalca, A.; Keeney, A.; Donnelly, G.; Shamblin, J.; Hensley, L.; Olinger, G.; et al. Aerosol Exposure to Rift Valley Fever Virus Causes Earlier and More Severe Neuropathology in the Murine Model, which Has Important Implications for Therapeutic Development. PLoS Negl. Trop. Dis. 2013, 7, e2156. [Google Scholar] [CrossRef] [Green Version]
- Scharton, D.; Bailey, K.W.; Vest, Z.; Westover, J.B.; Kumaki, Y.; Van Wettere, A.; Furuta, Y.; Gowen, B.B. Favipiravir (T-705) protects against peracute Rift Valley fever virus infection and reduces delayed-onset neurologic disease observed with ribavirin treatment. Antivir. Res. 2014, 104, 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, T.R.; Dyall, J.; Mercer, L.; Goodin, C.; Foil, D.H.; Zhou, H.; Postnikova, E.; Liang, J.Y.; Holbrook, M.R.; Madrid, P.B.; et al. Repurposing Pyramax®, quinacrine and tilorone as treatments for Ebola virus disease. Antivir. Res. 2020, 182, 104908. [Google Scholar] [CrossRef] [PubMed]
- Mayer, G.D.; Krueger, R.F. Tilorone Hydrochloride: Mode of Action. Science 1970, 169, 1214–1215. [Google Scholar] [CrossRef]
- Ekins, S.; Madrid, P.B. Tilorone, a Broad-Spectrum Antiviral for Emerging Viruses. Antimicrob. Agents Chemother. 2020, 64, e00440-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salata, C.; Calistri, A.; Parolin, C.; Baritussio, A.; Palù, G. Antiviral activity of cationic amphiphilic drugs. Expert Rev. Anti-Infect. Ther. 2017, 15, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.K.; Gieselmann, V.; Hasilik, A.; VON Figura, K. Tilorone Acts as a Lysosomotropic Agent in Fibroblasts. Hoppe-Seyler’s Z. Physiol. Chem. 1984, 365, 859–866. [Google Scholar] [CrossRef]
- Smith, D.R.; Steele, K.E.; Shamblin, J.; Honko, A.; Johnson, J.; Reed, C.; Kennedy, M.; Chapman, J.L.; Hensley, L.E. The pathogenesis of Rift Valley fever virus in the mouse model. Virology 2010, 407, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Warren, T.K.; Wells, J.; Panchal, R.G.; Stuthman, K.S.; Garza, N.L.; Van Tongeren, S.A.; Dong, L.; Retterer, C.J.; Eaton, B.P.; Pegoraro, G.; et al. Protection against filovirus diseases by a novel broad-spectrum nucleoside analogue BCX4430. Nature 2014, 508, 402–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kende, M.; Alving, C.R.; Rill, W.L.; Swartz, G.M.; Canonico, P.G. Enhanced efficacy of liposome-encapsulated ribavirin against Rift Valley fever virus infection in mice. Antimicrob. Agents Chemother. 1985, 27, 903–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedict, A.; Bansal, N.; Senina, S.; Hooper, I.; Lundberg, L.; De La Fuente, C.; Narayanan, A.; Gutting, B.W.; Kehn-Hall, K. Repurposing FDA-approved drugs as therapeutics to treat Rift Valley fever virus infection. Front. Microbiol. 2015, 6, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Time [HPI] | CC50 [μM] |
|---|---|---|
| Vero CCL81 | 24 | >100 |
| 48 | 30.76 | |
| 72 | 34.86 | |
| A549 | 24 | >100 |
| 48 | 11.23 | |
| 72 | 6.64 |
| RVFV Strain | Cell Line | EC50 (μM) | EC90 (μM) | SI 1 |
|---|---|---|---|---|
| MP12 | Vero CCL81 | 0.67 | 3.08 | >149 |
| A549 | 1.41 | 8.87 | >71 | |
| ZH501 | Vero CCL81 | 6.45 | 17.78 | >16 |
| A549 | 6.31 | 31.62 | >16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, K.N.; Kalveram, B.; Smith, J.K.; Zhang, L.; Juelich, T.; Atkins, C.; Ikegami, T.; Freiberg, A.N. Tilorone-Dihydrochloride Protects against Rift Valley Fever Virus Infection and Disease in the Mouse Model. Microorganisms 2022, 10, 92. https://doi.org/10.3390/microorganisms10010092
Johnson KN, Kalveram B, Smith JK, Zhang L, Juelich T, Atkins C, Ikegami T, Freiberg AN. Tilorone-Dihydrochloride Protects against Rift Valley Fever Virus Infection and Disease in the Mouse Model. Microorganisms. 2022; 10(1):92. https://doi.org/10.3390/microorganisms10010092
Chicago/Turabian StyleJohnson, Kendra N., Birte Kalveram, Jennifer K. Smith, Lihong Zhang, Terry Juelich, Colm Atkins, Tetsuro Ikegami, and Alexander N. Freiberg. 2022. "Tilorone-Dihydrochloride Protects against Rift Valley Fever Virus Infection and Disease in the Mouse Model" Microorganisms 10, no. 1: 92. https://doi.org/10.3390/microorganisms10010092
APA StyleJohnson, K. N., Kalveram, B., Smith, J. K., Zhang, L., Juelich, T., Atkins, C., Ikegami, T., & Freiberg, A. N. (2022). Tilorone-Dihydrochloride Protects against Rift Valley Fever Virus Infection and Disease in the Mouse Model. Microorganisms, 10(1), 92. https://doi.org/10.3390/microorganisms10010092