
Genomic Features and Construction of Streamlined Genome Chassis of Nisin Z Producer Lactococcus lactis N8

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain and Media
2.2. DNA Extraction and Genome Sequencing
2.3. Genome Analysis
2.3.1. Genome Annotation
2.3.2. Pan-Genome and Phylogenetic Analysis
2.3.3. Prophages Prediction
2.3.4. Genomic Islands Prediction
2.3.5. Genome Mining for Secondary Metabolite Biosynthetic Gene Clusters
2.4. Methods of Genome Streamlining
2.5. Resequencing
2.6. Phenotype Testing
2.6.1. Determination of Growth Profiles
2.6.2. Determination of Nisin Titre
2.7. Statistical Analysis
3. Results and Discussion
3.1. General Features of the Genome
3.2. Phylogeny and Core-Pan Genome
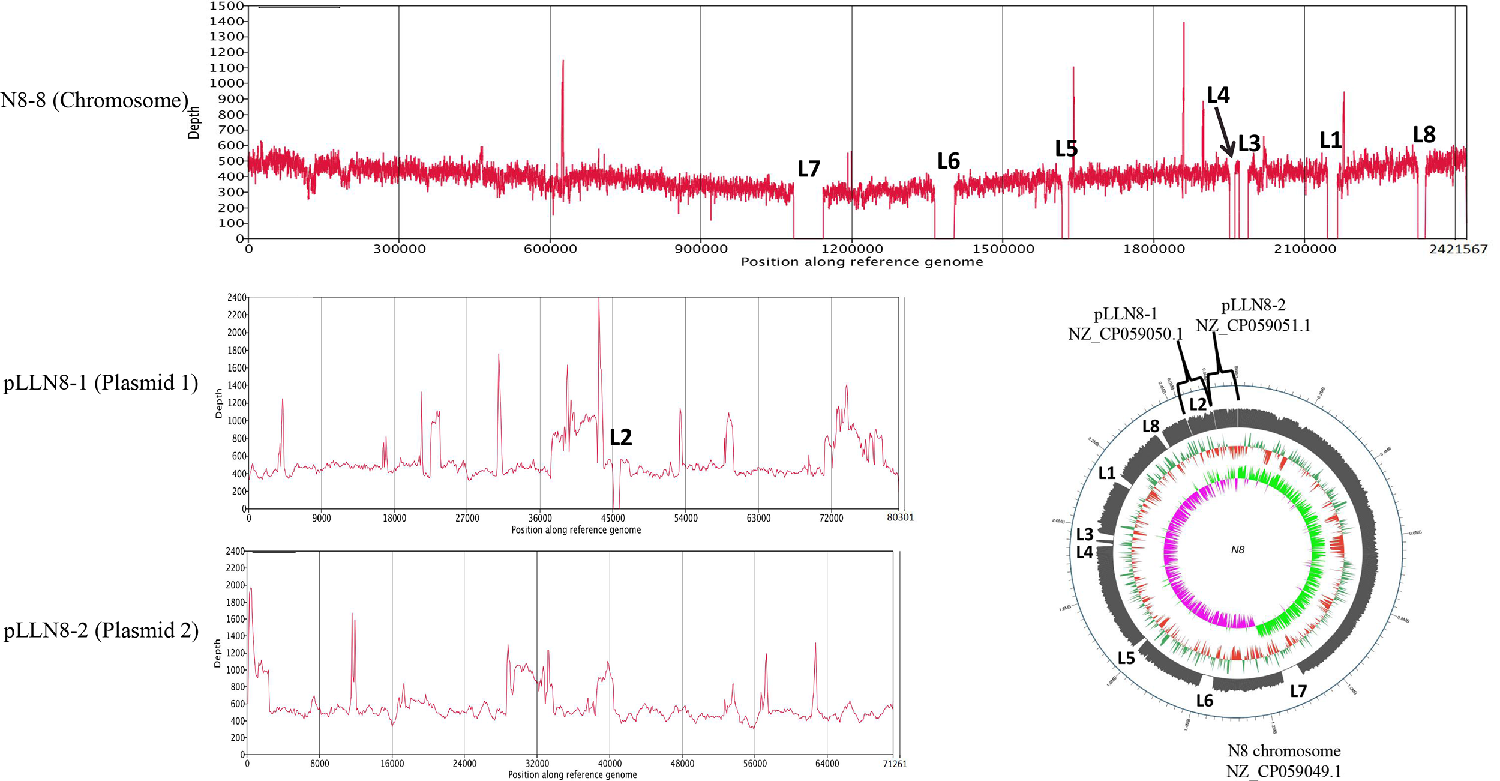
3.3. Megaplasmids of L. lactis N8
3.4. Prophage-Related Fragments of L. lactis N8
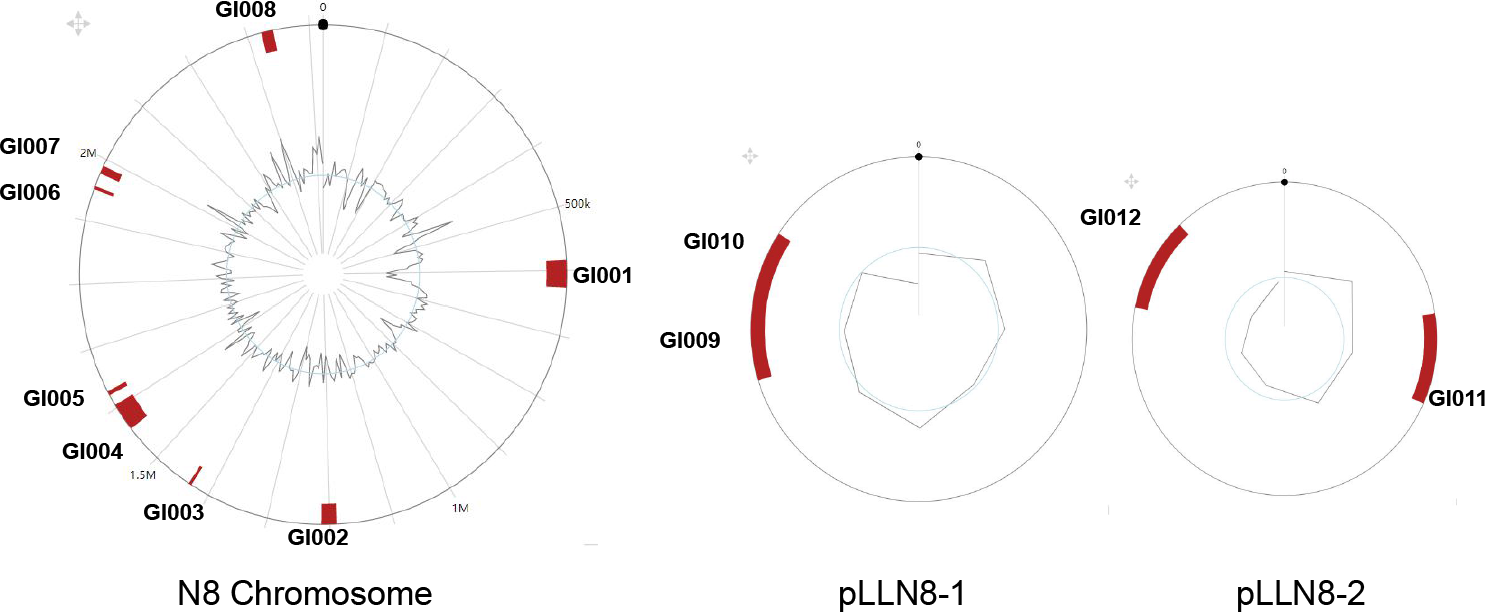
3.5. Genomic Islands of L. lactis N8
3.6. Other Characteristics of L. lactis N8
3.6.1. Cell-Surface Protein
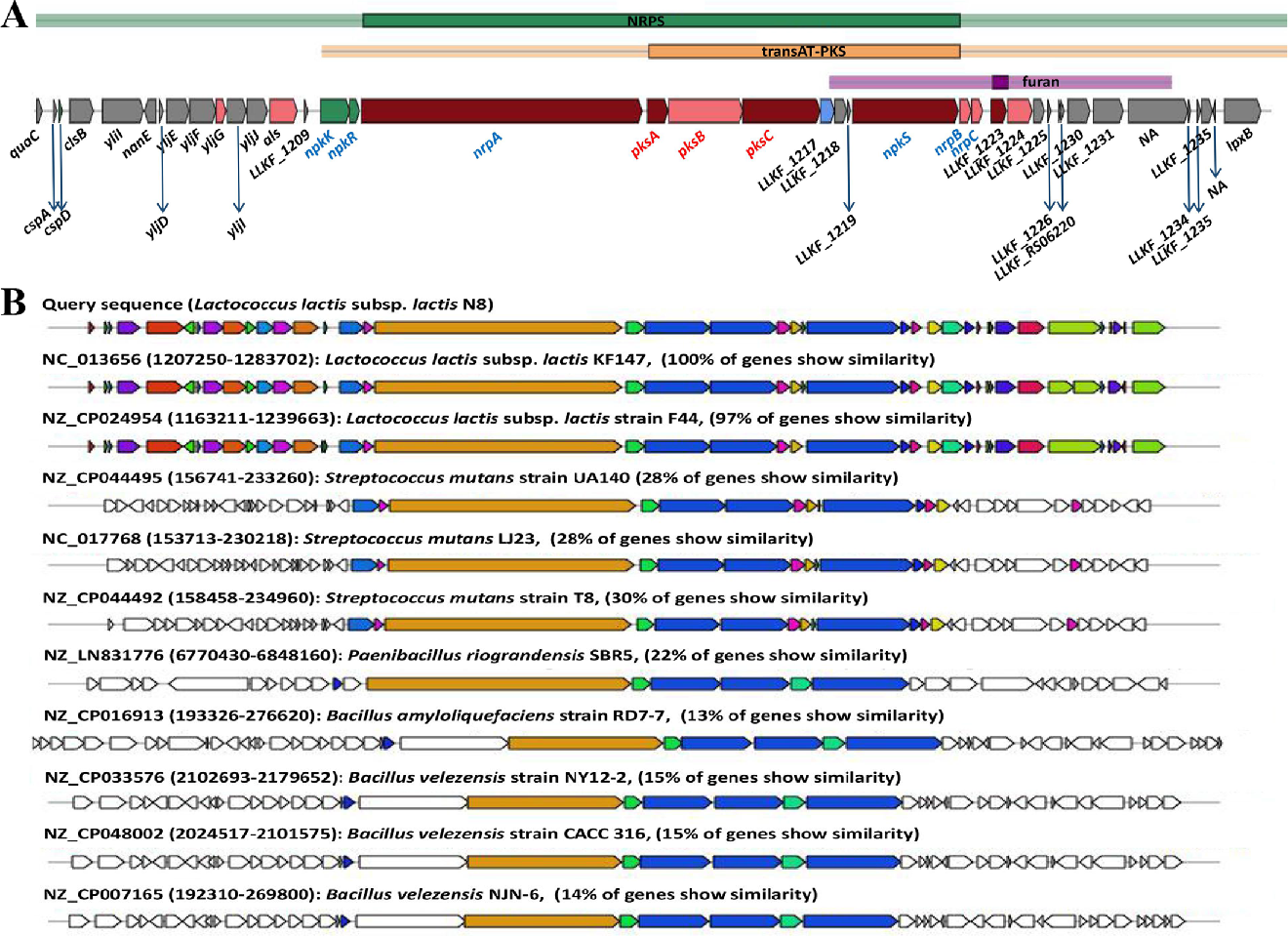
3.6.2. PKS/NRPS
3.6.3. Bacteriocin Gene Cluster
3.7. Construction of Streamlined Genome Chassis of L. lactis N8
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schleifer, K.H.; Kilpper-Bälz, R. Molecular and Chemotaxonomic Approaches to the Classification of Streptococci, Enterococci and Lactococci: A Review. Syst. Appl. Microbiol. 1987, 10, 1–19. [Google Scholar] [CrossRef]
- Pérez, T.; Balcázar, J.L.; Peix, A.; Valverde, A.; Velázquez, E.; De Blas, I.; Ruiz-Zarzuela, I. Lactococcus lactis subsp. tructae subsp. nov. isolated from the intestinal mucus of brown trout (Salmo trutta) and rainbow trout (Oncorhynchus mykiss). Int. J. Syst. Evol. Microbiol. 2011, 61, 1894–1898. [Google Scholar] [CrossRef]
- Ainsworth, S.; Stockdale, S.; Bottacini, F.; Mahony, J.; Van Sinderen, D. The Lactococcus lactis plasmidome: Much learnt, yet still lots to discover. FEMS Microbiol. Rev. 2014, 38, 1066–1088. [Google Scholar] [CrossRef]
- Kelleher, P.; Bottacini, F.; Mahony, J.; Kilcawley, K.N.; Van Sinderen, D. Comparative and functional genomics of the Lactococcus lactis taxon; insights into evolution and niche adaptation. BMC Genom. 2017, 18, 267. [Google Scholar] [CrossRef]
- Qiao, M.; Saris, P.E.J. Evidence for a role of NisT in transport of the lantibiotic nisin produced by Lactococcus lactis N8. FEMS Microbiol. Lett. 1996, 144, 89–93. [Google Scholar] [CrossRef][Green Version]
- Simsek, Ö.; Saris, P.E.J. Cycle changing the medium results in increased nisin productivity per cell in Lactococcus lactis. Biotechnol. Lett. 2009, 31, 415–421. [Google Scholar] [CrossRef]
- Koponen, O.; Tolonen, M.; Qiao, M.; Wahlström, G.; Helin, J.; Saris, P.E.J. NisB is required for the dehydration and NisC for the lanthionine formation in the post-translational modification of nisin. Microbiology 2002, 148, 3561–3568. [Google Scholar] [CrossRef]
- Takala, T.M.; Koponen, O.; Qiao, M.; Saris, P.E.J. Lipid-free NisI: Interaction with nisin and contribution to nisin immunity via secretion. FEMS Microbiol. Lett. 2004, 237, 171–177. [Google Scholar] [CrossRef]
- Xu, Y.; Li, X.; Li, R.; Li, S.; Ni, H.; Wang, H.; Xu, H.; Zhou, W.; Saris, P.E.J.; Yang, W.; et al. Structure of the nisin leader peptidase NisP revealing a C-terminal autocleavage activity. Acta Crystallogr. Sect. D 2014, 70, 1499–1505. [Google Scholar] [CrossRef]
- Kuipers, O.P.; De Ruyter, P.G.G.A.; Kleerebezem, M.; De Vos, W.M. Quorum sensing-controlled gene expression in lactic acid bacteria. J. Biotechnol. 1998, 64, 15–21. [Google Scholar] [CrossRef]
- Immonen, T.; Saris, P.E.J. Characterization of the nisFEG Operon of the Nisin Z Producing Lactococcus lactis subsp. lactis N8 Strain. DNA Seq. 1998, 9, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.; Qiao, Y.; Liu, F.; Zhang, Y.; Li, R.; Wu, Z.; Xu, H.; Saris, P.E.J.; Qiao, M. Engineering Lactococcus lactis as a multi-stress tolerant biosynthetic chassis by deleting the prophage-related fragment. Microb. Cell Fact. 2020, 19, 225. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Xuanyuan, Z.; Li, R.; Jiang, D.; Li, C.; Xu, H.; Bai, Y.; Zhang, X.; Turakainen, H.; Saris, P.E.J.; et al. Mu transposition complex mutagenesis in Lactococcus lactis–identification of genes affecting nisin production. J. Appl. Microbiol. 2009, 106, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Li, R.; Liu, F.; Xu, H.; Li, B.; Yuan, Y.; Saris, P.E.J.; Qiao, M. Mu insertion in feuD triggers the increase in nisin immunity in Lactococcus lactis subsp. lactis N8. J. Appl. Microbiol. 2016, 120, 402–412. [Google Scholar] [CrossRef]
- Chandrasekar Rajendran, S.C.; Chamlagain, B.; Kariluoto, S.; Piironen, V.; Saris, P.E.J. Biofortification of riboflavin and folate in idli batter, based on fermented cereal and pulse, by Lactococcus lactis N8 and Saccharomyces boulardii SAA655. J. Appl. Microbiol. 2017, 122, 1663–1671. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Fan, H.; Zhang, Y.; Cao, Y.; Li, T.; Qiao, W.; Wu, M.; Ma, T.; Li, G. Production of nisin-containing bacterial cellulose nanomaterials with antimicrobial properties through co-culturing Enterobacter sp. FY-07 and Lactococcus lactis N8. Carbohydr. Polym. 2021, 251, 117131. [Google Scholar] [CrossRef]
- Fu, Y.; Mu, D.; Qiao, W.; Zhu, D.; Wang, X.; Liu, F.; Xu, H.; Saris, P.; Kuipers, O.P.; Qiao, M. Co-expression of Nisin Z and Leucocin C as a Basis for Effective Protection Against Listeria monocytogenes in Pasteurized Milk. Front. Microbiol. 2018, 9, 547. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez-Humarán, L.G.; Aubry, C.; Motta, J.-P.; Deraison, C.; Steidler, L.; Vergnolle, N.; Chatel, J.-M.; Langella, P. Engineering lactococci and lactobacilli for human health. Curr. Opin. Microbiol. 2013, 16, 278–283. [Google Scholar] [CrossRef]
- Zhu, D.; Fu, Y.; Liu, F.; Xu, H.; Saris, P.E.J.; Qiao, M. Enhanced heterologous protein productivity by genome reduction in Lactococcus lactis NZ9000. Microb. Cell Fact. 2017, 16, 1. [Google Scholar] [CrossRef]
- Zhu, D.; Zhao, K.; Xu, H.; Zhang, X.; Bai, Y.; Saris, P.E.J.; Qiao, M. Construction of thyA deficient Lactococcus lactis using the Cre-loxP recombination system. Ann. Microbiol. 2015, 65, 1659–1665. [Google Scholar] [CrossRef]
- Wan, X.; Takala Timo, M.; Qiao, M.; Saris Per, E.J. Complete Genome Sequence of Nisin-Producing Lactococcus lactis subsp. lactis N8. Microbiol. Resour. Announc. 2021, 10, e01120–e01147. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, D.M.; Crowther, P.J.; Doherty, J.; Jefferson, S.; Decruz, E.; Noyer-Weidner, M.; Smith, S.S.; Michael, M.Z.; Graham, M.W. Quantitative evaluation of Escherichia coli host strains for tolerance to cytosine methylation in plasmid and phage recombinants. Nucleic Acids Res. 1989, 17, 3469–3478. [Google Scholar] [CrossRef] [PubMed]
- Qiao, M.; Ye, S.; Koponen, O.; Ra, R.; Usabiaga, M.; Immonen, T.; Saris, P.E.J. Regulation of the nisin operons in Lactococcus lactis N8. J. Appl. Bacteriol. 1996, 80, 626–634. [Google Scholar] [CrossRef]
- Qiao, M.; Immonen, T.; Koponen, O.; Saris, P.E.J. The cellular location and effect on nisin immunity of the NisI protein from Lactococcus lactis N8 expressed in Escherichia coli and L. lactis. FEMS Microbiol. Lett. 1995, 131, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Lambert Jolanda, M.; Bongers Roger, S.; Kleerebezem, M. Cre-lox-Based System for Multiple Gene Deletions and Selectable-Marker Removal in Lactobacillus plantarum. Appl. Environ. Microbiol. 2007, 73, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Tatusova, T.; Dicuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; Von Mering, C.; Bork, P. Fast Genome-Wide Functional Annotation through Orthology Assignment by eggNOG-Mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Wu, S.; Zhu, Z.; Fu, L.; Niu, B.; Li, W. WebMGA: A customizable web server for fast metagenomic sequence analysis. BMC Genom. 2011, 12, 444. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Simon Fraser University Research Computing Group; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.p.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Tang, X.; Nakata, Y.; Li, H.O.; Zhang, M.; Gao, H.; Fujita, A.; Sakatsume, O.; Ohta, T.; Yokoyama, K. The optimization of preparations of competent cells for transformation of E. coli. Nucleic Acids Res. 1994, 22, 2857–2858. [Google Scholar] [CrossRef]
- Li, T.T.; Tian, W.L.; Gu, C.T. Elevation of Lactococcus lactis subsp. cremoris to the species level as Lactococcus cremoris sp. nov. and transfer of Lactococcus lactis subsp. tructae to Lactococcus cremoris as Lactococcus cremoris subsp. tructae comb. nov. Int. J. Syst. Evol. Microbiol. 2021, 71, 4727. [Google Scholar] [CrossRef]
- Malesevic, M.; Stanisavljevic, N.; Miljkovic, M.; Jovcic, B.; Filipic, B.; Studholme, D.J.; Kojic, M. The large plasmidome of Lactococcus lactis subsp. lactis bv. diacetylactis S50 confers its biotechnological properties. Int. J. Food Microbiol. 2021, 337, 108935. [Google Scholar] [CrossRef]
- Cords, B.R.; Mckay, L.L.; Guerry, P. Extrachromosomal Elements in Group N Streptococci1. J. Bacteriol. 1974, 117, 1149–1152. [Google Scholar] [CrossRef]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic systems of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Chopin, A.; Bolotin, A.; Sorokin, A.; Ehrlich, S.D.; Chopin, M.-C. Analysis of six prophages in Lactococcus lactis IL1403: Different genetic structure of temperate and virulent phage populations. Nucleic Acids Res. 2001, 29, 644–651. [Google Scholar] [CrossRef]
- Ventura, M.; Zomer, A.; Canchaya, C.; O’Connell-Motherway, M.; Kuipers, O.; Turroni, F.; Ribbera, A.; Foroni, E.; Buist, G.; Wegmann, U.; et al. Comparative Analyses of Prophage-Like Elements Present in Two Lactococcus lactis Strains. Appl. Environ. Microbiol. 2007, 73, 7771–7780. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, P.; Mahony, J.; Schweinlin, K.; Neve, H.; Franz, C.M.; Van Sinderen, D. Assessing the functionality and genetic diversity of lactococcal prophages. Int. J. Food Microbiol. 2018, 272, 29–40. [Google Scholar] [CrossRef]
- O’Sullivan, D.J.; Lee, J.H.; Dominguez, W. Lactic Acid Bacteria: | Genomics, Genetic Engineering. In Encyclopedia of Dairy Sciences; Fuquay, J.W., McSweeney, P.L., Fox, P.F., Eds.; Academic Press: Cambridge, MA, USA, 2011; pp. 67–77. [Google Scholar]
- Deveau, H.; Labrie Simon, J.; Chopin, M.-C.; Moineau, S. Biodiversity and Classification of Lactococcal Phages. Appl. Environ. Microbiol. 2006, 72, 4338–4346. [Google Scholar] [CrossRef] [PubMed]
- Hacker, J.; Kaper, J.B. Pathogenicity Islands and the Evolution of Microbes. Annu. Rev. Microbiol. 2000, 54, 641–679. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Hsiao, W.W.L.; Brinkman, F.S.L. Detecting genomic islands using bioinformatics approaches. Nat. Rev. Microbiol. 2010, 8, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Vernikos, G.S.; Parkhill, J. Resolving the structural features of genomic islands: A machine learning approach. Genome Res. 2008, 18, 331–342. [Google Scholar] [CrossRef]
- Umenhoffer, K.; Fehér, T.; Balikó, G.; Ayaydin, F.; Pósfai, J.; Blattner, F.R.; Pósfai, G. Reduced evolvability of Escherichia coli MDS42, an IS-less cellular chassis for molecular and synthetic biology applications. Microb. Cell Fact. 2010, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Siezen, R.; Boekhorst, J.; Muscariello, L.; Molenaar, D.; Renckens, B.; Kleerebezem, M. Lactobacillus plantarum gene clusters encoding putative cell-surface protein complexes for carbohydrate utilization are conserved in specific gram-positive bacteria. BMC Genom. 2006, 7, 126. [Google Scholar] [CrossRef] [PubMed]
- Golomb, B.L.; Yu, A.O.; Coates, L.C.; Marco, M.L. The Lactococcus lactis KF147 nonribosomal peptide synthetase/polyketide synthase system confers resistance to oxidative stress during growth on plant leaf tissue lysate. Microbiol. Open 2018, 7, e00531. [Google Scholar] [CrossRef]
- Khayatt, B.I.; Van Noort, V.; Siezen, R.J. The Genome of the Plant-Associated Lactic Acid Bacterium Lactococcus lactis KF147 Harbors a Hybrid NRPS-PKS System Conserved in Strains of the Dental Cariogenic Streptococcus mutans. Curr. Microbiol. 2020, 77, 136–145. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmids | Relevant Descriptions | Reference |
|---|---|---|
| Strains | ||
| E. coli DH5α | Cloning host; F-φ80lacZ∆M15endA1 recA1 endA1 hsdR17 (rK-mK+) supE44 thi-1 gyrA 96 relA1 ∆(lacZYA-argF)U169 deoR λ- | [22] |
| E. coli DH5α-up-down (L1) | Cmr, Emr, E. coli DH5α derivative containing the whole plasmid pNZ5319∆L1 | [12] |
| E. coli DH5α-up-down (L2) | Cmr, Emr, E. coli DH5α derivative containing the whole plasmid pNZ5319∆L2 | This study |
| E. coli DH5α-up-down (L3) | Cmr, Emr, E. coli DH5α derivative containing the whole plasmid pNZ5319∆L3 | This study |
| E. coli DH5α-up-down (L4) | Cmr, Emr, E. coli DH5α derivative containing the whole plasmid pNZ5319∆L4 | This study |
| E. coli DH5α-up-down (L5) | Cmr, Emr, E. coli DH5α derivative containing the whole plasmid pNZ5319∆L5 | This study |
| E. coli DH5α-up-down (L6) | Cmr, Emr, E. coli DH5α derivative containing the whole plasmid pNZ5319∆L6 | This study |
| E. coli DH5α-up-down (L7) | Cmr, Emr, E. coli DH5α derivative containing the whole plasmid pNZ5319∆L7 | This study |
| E. coli DH5α-up-down (L8) | Cmr, Emr, E. coli DH5α derivative containing the whole plasmid pNZ5319∆L8 | This study |
| Micrococcus luteus NCIB 8166 | Indicator strains for Nisin agar gel diffusion assay | [23] |
| L. lactis N8 | Wild-type (WT) Nisin Z producer | [24] |
| L. lactis N8-1 | The first DNA region L1 deletion in L. lactis N8 | [12] |
| L. lactis N8-2 | The L2 deletion in L. lactis N8-1 | This study |
| L. lactis N8-3 | The L3 deletion in L. lactis N8-2 | This study |
| L. lactis N8-4 | The L4 deletion in L. lactis N8-3 | This study |
| L. lactis N8-5 | The L5 deletion in L. lactis N8-4 | This study |
| L. lactis N8-6 | The L6 deletion in L. lactis N8-5 | This study |
| L. lactis N8-7 | The L7 deletion in L. lactis N8-6 | This study |
| L. lactis N8-8 | The L8 deletion in L. lactis N8-7 | This study |
| Plasmids | ||
| pNZ5319 | Cmr, Emr, used as knock-out vector | [25] |
| pNZTS-Cre | Emr, cre gene cloned at the EcoRI and HindIII sites (cat gene deletion vector) | [20] |
| pNZ5319∆L1 | Cmr, Emr, upstream and downstream homology arm of L1 amplified from L. lactis N8 genome cloned into pNZ5319 | [12] |
| pNZ5319∆L2 | Cmr, Emr, upstream and downstream homology arm of L2 amplified from L. lactis N8 genome cloned into pNZ5319 | This study |
| pNZ5319∆L3 | Cmr, Emr, upstream and downstream homology arm of L3 amplified from L. lactis N8 genome cloned into pNZ5319 | This study |
| pNZ5319∆L4 | Cmr, Emr, upstream and downstream homology arm of L4 amplified from L. lactis N8 genome cloned into pNZ5319 | This study |
| pNZ5319∆L5 | Cmr, Emr, upstream and downstream homology arm of L5 amplified from L. lactis N8 genome cloned into pNZ5319 | This study |
| pNZ5319∆L6 | Cmr, Emr, upstream and downstream homology arm of L6 amplified from L. lactis N8 genome cloned into pNZ5319 | This study |
| pNZ5319∆L7 | Cmr, Emr, upstream and downstream homology arm of L7 amplified from L. lactis N8 genome cloned into pNZ5319 | This study |
| pNZ5319∆L8 | Cmr, Emr, upstream and downstream homology arm of L8 amplified from L. lactis N8 genome cloned into pNZ5319 | This study |
| Feature | Size/Percentage/Number |
|---|---|
| Genome size (bp) | 2,421,567 |
| G + C content | 35.1% |
| rRNAs | 19 |
| tRNAs | 64 |
| ncRNAs | 4 |
| Total genes | 2521 |
| Coding sequences (CDs) | 2434 |
| Prophages | 7 |
| Genomic islands (GIs) | 12 |
| Plasmids | 2 |
| Organism Name | Strain | BioSample | Number of Plasmids | Total Size of Plasmids | Release Date |
|---|---|---|---|---|---|
| Lactococcus lactis subsp. lactis | N8 | SAMN15500618 | 2 | 0.151562 | 22 October 2020 |
| Lactococcus lactis subsp. lactis | No attributes | SAMN14223931 | 1 | 0.060232 | 10 September 1998 |
| Lactococcus lactis subsp. lactis | CBA3619 | SAMN11843663 | 1 | 0.107586 | 16 August 2019 |
| Lactococcus lactis subsp. lactis | FDAARGOS_865 | SAMN13450395 | 1 | 0.058335 | 15 December 2020 |
| Lactococcus lactis subsp. lactis | FDAARGOS_887 | SAMN13450417 | 4 | 0.180362 | 15 December 2020 |
| Lactococcus lactis subsp. lactis | FDAARGOS_866 | SAMN13450396 | 4 | 0.241108 | 15 December 2020 |
| Lactococcus lactis subsp. lactis | FDAARGOS_1064 | SAMN16357233 | 4 | 0.112979 | 21 December 2020 |
| Lactococcus lactis subsp. lactis | WiKim0098 | SAMN16788728 | 1 | 0.076987 | 18 January 2021 |
| Lactococcus lactis subsp. lactis | A12 | SAMEA4005236 | 4 | 0.126727 | 23 July 2016 |
| Lactococcus lactis subsp. lactis | 229 | SAMN04955249 | 5 | 0.165685 | 5 April 2017 |
| Lactococcus lactis subsp. lactis | 275 | SAMN04955252 | 4 | 0.25801 | 5 April 2017 |
| Lactococcus lactis subsp. lactis | UC063 | SAMN04956302 | 5 | 0.149078 | 5 April 2017 |
| Lactococcus lactis subsp. lactis | UL8 | SAMN04956402 | 3 | 0.037067 | 10 April 2017 |
| Lactococcus lactis subsp. lactis | 14B4 | SAMN08792430 | 1 | 0.0597 | 1 June 2018 |
| Lactococcus lactis subsp. lactis | 184 | SAMN04955247 | 6 | 0.042416 | 11 January 2019 |
| Lactococcus lactis subsp. lactis | C10 | SAMN04956267 | 3 | 0.056865 | 11 January 2019 |
| Lactococcus lactis subsp. lactis | UC06 | SAMN04956292 | 6 | 0.155687 | 13 January 2019 |
| Lactococcus lactis subsp. lactis | UC08 | SAMN04956293 | 5 | 0.166766 | 16 January 2019 |
| Lactococcus lactis subsp. lactis | UC77 | SAMN04956303 | 5 | 0.178184 | 3 January 2019 |
| Lactococcus lactis subsp. lactis | UC11 | SAMN04956294 | 6 | 0.156382 | 11 September 2019 |
| Lactococcus lactis subsp. lactis | G121 | SAMN14943687 | 3 | 0.123307 | 7 September 2020 |
| Lactococcus lactis subsp. lactis | 223 | SAMN09847869 | 6 | 0.05587 | 25 February 2021 |
| Lactococcus lactis subsp. lactis | WM1 | SAMN09847649 | 5 | 0.180672 | 23 February 2021 |
| Lactococcus lactis subsp. lactis | 267 | SAMN09847870 | 5 | 0.164327 | 27 February 2021 |
| Lactococcus lactis subsp. lactis | DRC3 | SAMN16604567 | 7 | 0.232822 | 10 April 2021 |
| Lactococcus lactis subsp. lactis | Lac3 | SAMN18740314 | 1 | 0.007367 | 13 May 2021 |
| Lactococcus lactis subsp. lactis bv. diacetylactis | FM03 | SAMN06061939 | 7 | 0.0809 | 24 May 2017 |
| Lactococcus lactis subsp. lactis bv. diacetylactis | SD96 | SAMN12502795 | 10 | 0.219931 | 7 October 2019 |
| Lactococcus lactis subsp. lactis bv. diacetylactis | BGBU1-4 | SAMN12627231 | 1 | 0.00633 | 17 November 2019 |
| Lactococcus lactis subsp. lactis bv. diacetylactis | S50 | SAMN10167144 | 6 | 0.240412 | 10 September 2020 |
| Lactococcus lactis subsp. lactis bv. diacetylactis | No attributes | SAMN14226089 | 1 | 0.018977 | 1 August 2011 |
| Lactococcus lactis subsp. lactis bv. diacetylactis | No attributes | SAMN14226088 | 1 | 0.021728 | 1 August 2011 |
| Lactococcus lactis subsp. lactis bv. diacetylactis | No attributes | SAMN14226087 | 1 | 0.022166 | 1 August 2011 |
| Lactococcus lactis subsp. lactis bv. diacetylactis | No attributes | SAMN14226080 | 1 | 0.053876 | 1 August 2011 |
| Lactococcus lactis subsp. lactis | CV56 | SAMN02603398 | 5 | 0.119279 | 1 May 2012 |
| Lactococcus lactis subsp. lactis | KF147 | SAMN02603087 | 1 | 0.03751 | 22 December 2009 |
| Lactococcus lactis subsp. lactis | KLDS 4.0325 | SAMN02603468 | 6 | 0.171622 | 10 May 2018 |
| Lactococcus lactis subsp. lactis | NCDO 2118 | SAMN02471376 | 1 | 0.037571 | 21 January 2015 |
| Average | 3.66 | 0.031197 | |||
| Prophage | Start | End | Size (bp) | Att Core Sequence | G+C Content | Status |
|---|---|---|---|---|---|---|
| LLN8-1 (chromosome) | 1,369,374 | 1,405,566 | 36,193 | TTTAATTTAGAAA | 35.27% | intact |
| LLN8-2 (chromosome) | 1,949,930 | 1,988,317 | 38,388 | AACGTAACTAAAAACGTAACTAA | 35.18% | intact |
| LLN8-3 (chromosome) | 2,128,037 | 2,162,403 | 34,367 | AACTTATTTTTAT | 34.31% | incomplete |
| LLN8-4 (chromosome) | 2,323,015 | 2,342,870 | 19,856 | ACGCTTTTTACTACGTTCG | 34.56% | incomplete |
| LLN8-5 (plasmid1) | 40,333 | 69,929 | 29,597 | AAAATAAAAAGT | 32.05% | incomplete |
| LLN8-6 (plasmid2) | 8658 | 53,552 | 44,895 | TTTCGAACATTT | 36.09% | questionable |
| LLN8-7 (plasmid2) | 53,728 | 69,125 | 15,398 | AGGTTCTGTTGCAAAGTT | 35.32% | questionable |
| Genomic Island | Start | End | Size (bp) | G+C Content % |
|---|---|---|---|---|
| GI001 (chromosome) | 583,454 | 626,486 | 43,032 | 30.63 |
| GI002 (chromosome) | 1,188,697 | 1,213,410 | 24,713 | 32.54 |
| GI003 (chromosome) | 1,431,036 | 1,435,913 | 4877 | 39.57 |
| GI004 (chromosome) | 1,560,579 | 1,604,865 | 44,286 | 35.98 |
| GI005 (chromosome) | 1,622,309 | 1,629,361 | 7052 | 30.91 |
| GI006 (chromosome) | 1,950,028 | 1,955,065 | 5037 | 30.06 |
| GI007 (chromosome) | 1,976,159 | 1,989,281 | 13,122 | 33.38 |
| GI008 (chromosome) | 2,322,308 | 2,341,772 | 19,464 | 34.55 |
| GI009 (plasmid 1) | 56,384 | 60,867 | 4483 | 37.20 |
| GI010 (plasmid 1) | 58,996 | 67,736 | 8740 | 33.58 |
| GI011 (plasmid 2) | 15,929 | 22,625 | 6696 | 36.81 |
| GI012 (plasmid 2) | 55,774 | 62,660 | 6886 | 32.64 |
| L. lactis | Removed (bp) | Cumulative (bp) | Deletion (%) | μmax (h−1) | Generation Time (min) |
|---|---|---|---|---|---|
| N8 | 0 | 0 | 0% | 0.44 ± 0.04 | 45.7 ± 2.7 |
| N8-1 | 19,739 | 19,739 | 0.77% | 0.48 ± 0.05 | 41.5 ± 5.0 |
| N8-2 | 1638 | 21,377 | 0.83% | 0.49 ± 0.06 | 42.7 ± 4.1 |
| N8-3 | 18,628 | 40,005 | 1.55% | 0.50 ± 0.03 | 40.7 ± 5.0 |
| N8-4 | 10,659 | 50,664 | 1.97% | 0.47 ± 0.03 | 40.0 ± 5.1 |
| N8-5 | 13,502 | 64,166 | 2.49% | 0.44 ± 0.02 | 41.3 ± 4.9 |
| N8-6 | 39,203 | 103,369 | 4.02% | 0.53 ± 0.09 | 41.1 ± 3.2 |
| N8-7 | 58,594 | 161,963 | 6.29% | 0.52 ± 0.03 | 39.3 ± 5.2 |
| N8-8 | 14,465 | 176,428 | 6.86% | 0.50 ± 0.05 | 39.0 ± 4.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, W.; Liu, F.; Wan, X.; Qiao, Y.; Li, R.; Wu, Z.; Saris, P.E.J.; Xu, H.; Qiao, M. Genomic Features and Construction of Streamlined Genome Chassis of Nisin Z Producer Lactococcus lactis N8. Microorganisms 2022, 10, 47. https://doi.org/10.3390/microorganisms10010047
Qiao W, Liu F, Wan X, Qiao Y, Li R, Wu Z, Saris PEJ, Xu H, Qiao M. Genomic Features and Construction of Streamlined Genome Chassis of Nisin Z Producer Lactococcus lactis N8. Microorganisms. 2022; 10(1):47. https://doi.org/10.3390/microorganisms10010047
Chicago/Turabian StyleQiao, Wanjin, Fulu Liu, Xing Wan, Yu Qiao, Ran Li, Zhenzhou Wu, Per Erik Joakim Saris, Haijin Xu, and Mingqiang Qiao. 2022. "Genomic Features and Construction of Streamlined Genome Chassis of Nisin Z Producer Lactococcus lactis N8" Microorganisms 10, no. 1: 47. https://doi.org/10.3390/microorganisms10010047
APA StyleQiao, W., Liu, F., Wan, X., Qiao, Y., Li, R., Wu, Z., Saris, P. E. J., Xu, H., & Qiao, M. (2022). Genomic Features and Construction of Streamlined Genome Chassis of Nisin Z Producer Lactococcus lactis N8. Microorganisms, 10(1), 47. https://doi.org/10.3390/microorganisms10010047