
Protective Effects of Novel Lactobacillaceae Strains Isolated from Chicken Caeca against Necrotic Enteritis Infection: In Vitro and In Vivo Evidences

 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Isolation of Lactic Acid Bacteria from Chicken Caeca and Their Mass Spectrometry Identification
2.2. Anti-Clostridium perfringens Activity
2.3. Quantification of Lactic Acid Production
2.4. Biofilm Formation
2.5. Resistance to Gastrointestinal Conditions in Chicken
2.6. Adhesion to Intestinal Cells
2.7. qPCR Assay
2.8. Biosafety Aspects of Newly Selected LAB Strains
2.8.1. Hemolytic Activity
2.8.2. Antibiotic Resistance
2.8.3. Cytotoxicity
2.9. Coccidiostat Analysis
2.10. In Vivo Evaluation of the Three Newly Isolated Lactobacillaceae Strains
2.10.1. Chicken Environment and Diet
2.10.2. Vaccination
2.10.3. C. perfringens Infection Protocol
2.10.4. Administration of LAB Strains
2.10.5. Measurements of Feed Uptake and Weight of Birds
2.11. Statistical Analysis
3. Results
3.1. Diversity of LAB Strain in Chicken Ceca
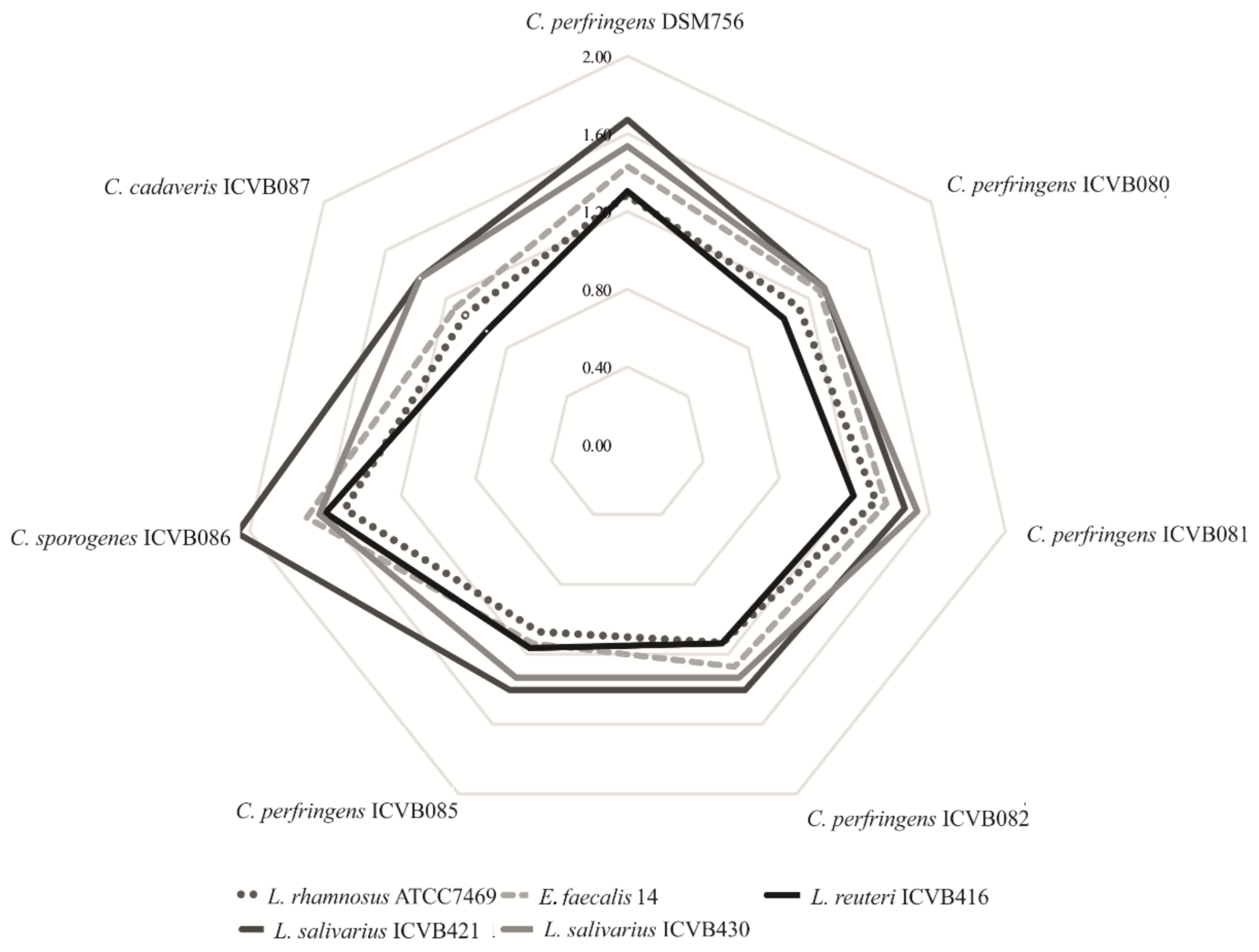
3.2. Newly ISOLATED LAB Strains Displayed Strong Anti-Clostridium perfringens Activity
3.3. Biofilm Formation
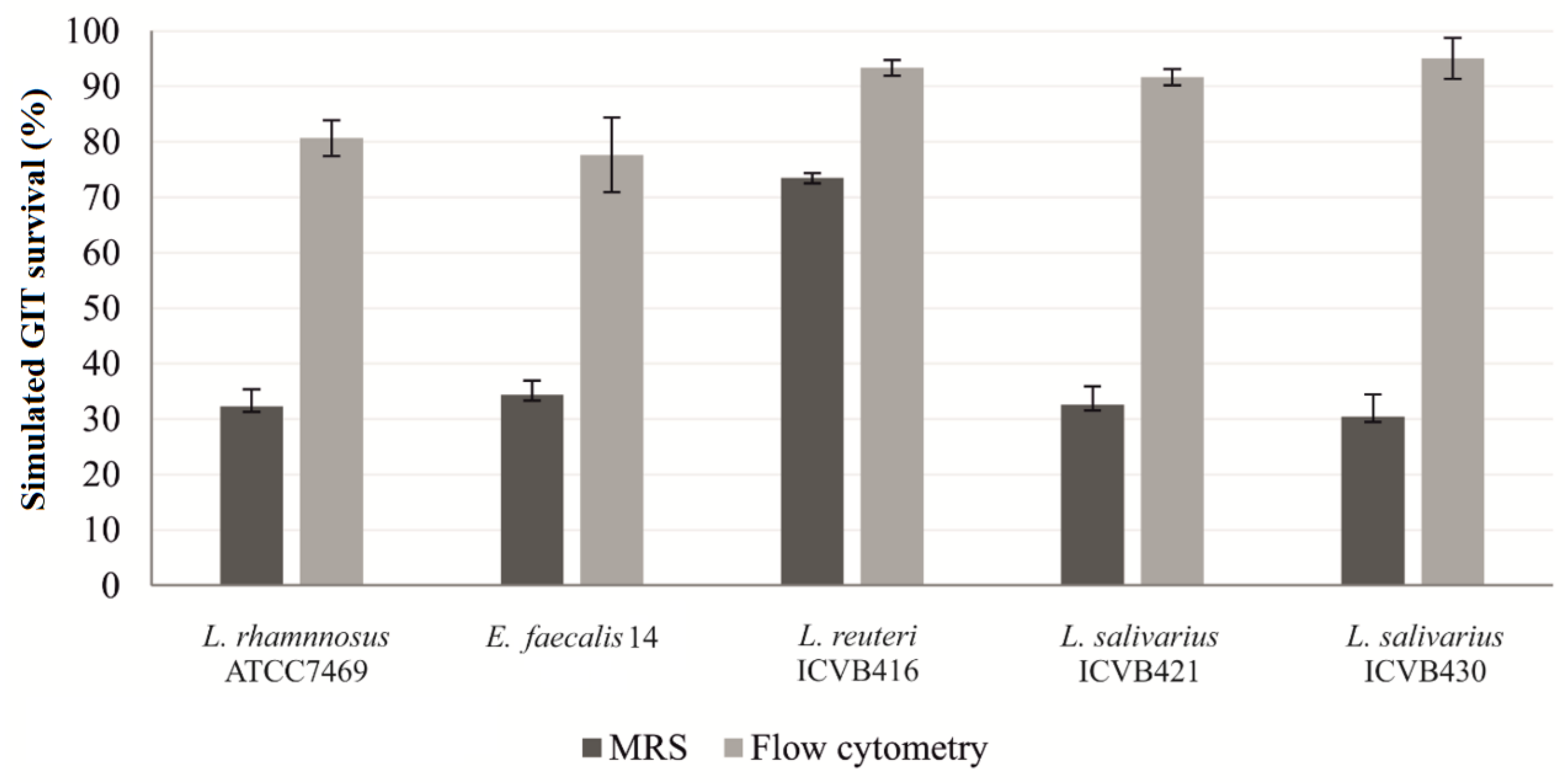
3.4. Resistance to Chicken Gastrointestinal Conditions
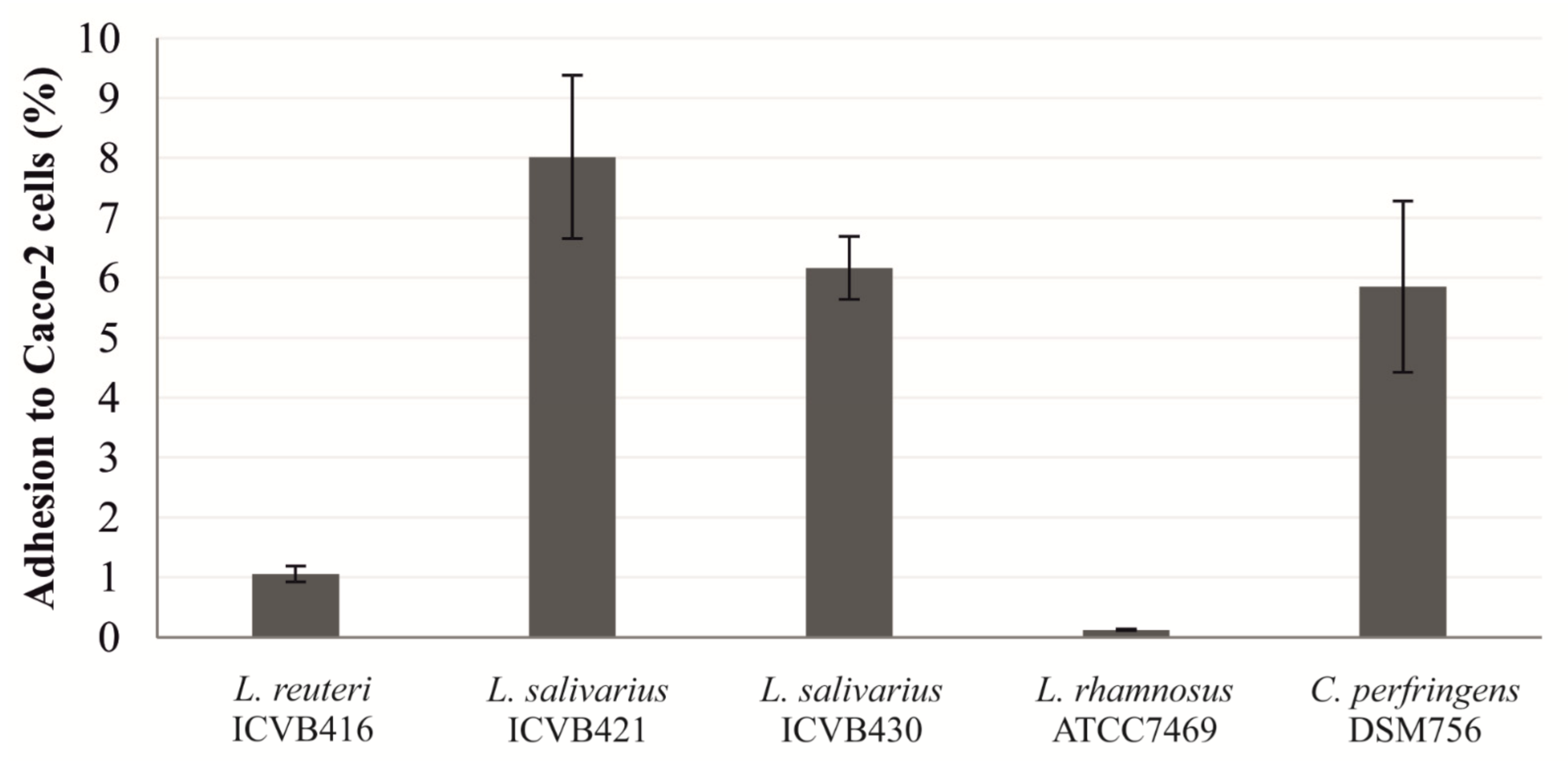
3.5. Adhesion to Intestinal Cells
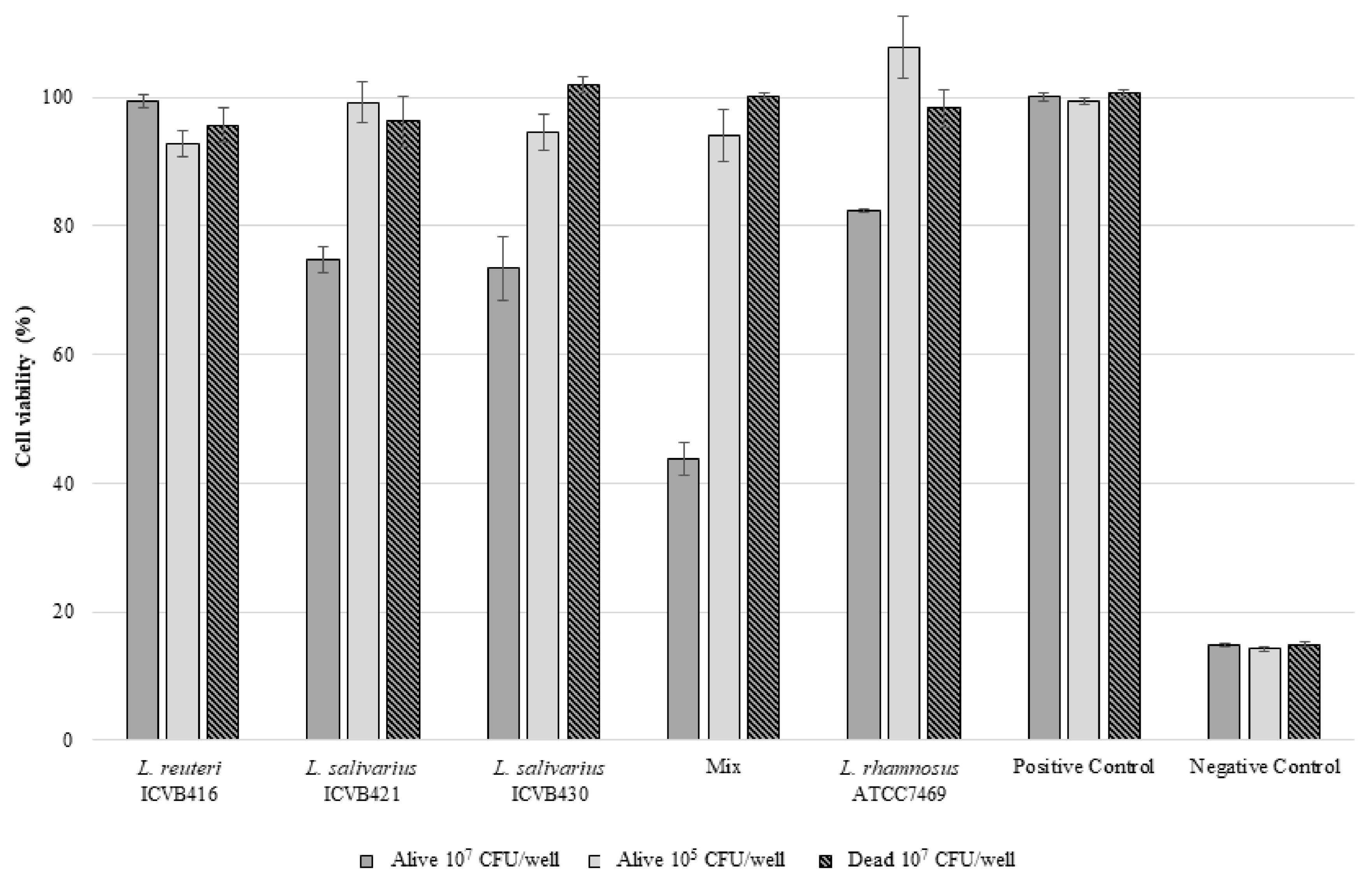
3.6. Safety Assessment of Lactobacillaceae Strains
3.7. Coccidiostat Analysis
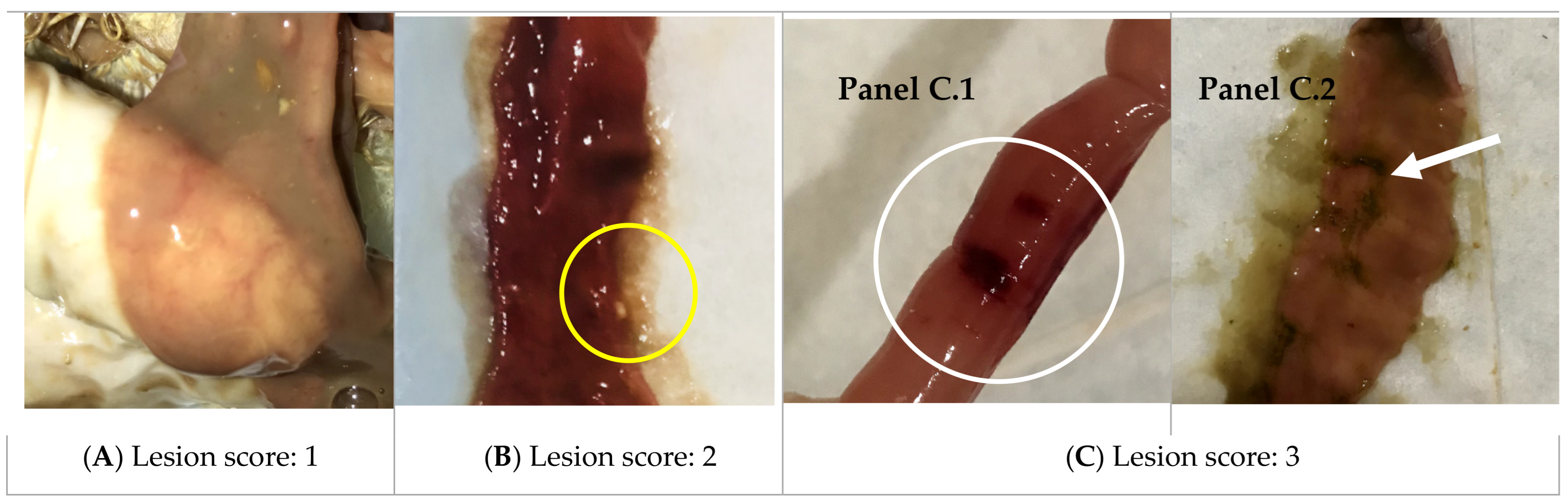
3.8. In Vivo Trials
Zootechnical Performance during the Start-Up Phase (0–14 Days)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laxminarayan, R.; van Boeckel, T.; Teillant, A. The Economic Costs of Withdrawing Antimicrobial Growth Promoters from the Livestock Sector; OECD Food, Agriculture and Fisheries Papers; OECD: Paris, France, 2015; Volume 78. [Google Scholar]
- WHO New Report Calls for Urgent Action to Avert Antimicrobial Resistance Crisis. Available online: https://www.who.int/news/item/29-04-2019-new-report-calls-for-urgent-action-to-avert-antimicrobial-resistance-crisis (accessed on 16 June 2021).
- CDC Antibiotic Resistance Threatens Everyone. Available online: https://www.cdc.gov/drugresistance/index.html (accessed on 10 June 2021).
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [Green Version]
- de Kraker, M.E.A.; Stewardson, A.J.; Harbarth, S. Will 10 Million People Die a Year due to Antimicrobial Resistance by 2050? PLoS Med. 2016, 13, e1002184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Rev. Antimicrob. Resist. 2016. Available online: https://amr-review.org/sites/default/files/160518_Final%20paper_with%20cover.pdf (accessed on 16 June 2021).
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Antibiotic Resistance: Moving from Individual Health Norms to Social Norms in One Health and Global Health. Front. Microbiol. 2020, 11, 1914. [Google Scholar] [CrossRef]
- Keyburn, A.L.; Boyce, J.D.; Vaz, P.; Bannam, T.L.; Ford, M.E.; Parker, D.; Di Rubbo, A.; Rood, J.I.; Moore, R.J. NetB, a New Toxin That Is Associated with Avian Necrotic Enteritis Caused by Clostridium perfringens. PLOS Pathog. 2008, 4, e26. [Google Scholar] [CrossRef] [Green Version]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe 2018, 53, 5–10. [Google Scholar] [CrossRef]
- Navarro, M.A.; McClane, B.A.; Uzal, F.A. Mechanisms of Action and Cell Death Associated with Clostridium perfringens Toxins. Toxins 2018, 10, 212. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, J.; Nagahama, M.; Oda, M. Clostridium perfringens Alpha-Toxin: Characterization and Mode of Action. J. Biochem. 2004, 136, 569–574. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.J. Necrotic enteritis predisposing factors in broiler chickens. Avian Pathol. 2016, 45, 275–281. [Google Scholar] [CrossRef] [Green Version]
- M’Sadeq, S.A.; Wu, S.; Swick, R.; Choct, M. Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide. Anim. Nutr. 2015, 1, 1–11. [Google Scholar] [CrossRef]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.W.; Harper, D.; et al. Alternatives to antibiotics—A pipeline portfolio review. Lancet Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Seal, B.S.; Lillehoj, H.S.; Donovan, D.M.; Gay, C.G. Alternatives to antibiotics: A symposium on the challenges and solutions for animal production. Anim. Heal. Res. Rev. 2013, 14, 78–87. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food. Guidelines for the Evaluation of Probiotics in Food. Available online: https://www.who.int/foodsafety/fs_management/en/probiotic_guidelines.pdf (accessed on 30 April 2020).
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: Lights and shadows. Front. Cell. Infect. Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef] [Green Version]
- Callaway, T.R.; Edrington, T.S.; Anderson, R.C.; Harvey, R.B.; Genovese, K.J.; Kennedy, C.N.; Venn, D.W.; Nisbet, D.J. Probiotics, prebiotics and competitive exclusion for prophylaxis against bacterial disease. Anim. Health Res. Rev. 2008, 9, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Klose, V.; Mohnl, M.; Plail, R.; Schatzmayr, G.; Loibner, A.-P. Development of a competitive exclusion product for poultry meeting the regulatory requirements for registration in the European Union. Mol. Nutr. Food Res. 2006, 50, 563–571. [Google Scholar] [CrossRef]
- Siedler, S.; Rau, M.H.; Bidstrup, S.; Vento, J.M.; Aunsbjerg, S.D.; Bosma, E.F.; McNair, L.; Beisel, C.L.; Neves, A.R. Competitive Exclusion is a major bioprotective mechanism of Lactobacilli against fungal spoilage in fermented milk products. Appl. Environ. Microbiol. 2020, 86, e02312-19. [Google Scholar] [CrossRef] [Green Version]
- Kiczorowska, B.; Samolińska, W.; Al-Yasiry, A.R.M.; Kiczorowski, P.; Winiarska-Mieczan, A. The natural feed additives as immunostimulants in monogastric animal nutrition—A review. Ann. Anim. Sci. 2017, 17, 605–625. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, J.; Laiño, J.E.; Del Valle, M.J.; Vannini, V.; Van Sinderen, D.; Taranto, M.; De Valdez, G.F.; De Giori, G.S.; Sesma, F. B-Group vitamin production by lactic acid bacteria—Current knowledge and potential applications. J. Appl. Microbiol. 2011, 111, 1297–1309. [Google Scholar] [CrossRef]
- Liao, S.F.; Nyachoti, M. Using probiotics to improve swine gut health and nutrient utilization. Anim. Nutr. 2017, 3, 331–343. [Google Scholar] [CrossRef]
- Sharifuzzaman, S.; Austin, B. Probiotics for disease control in aquaculture. Diagn. Control Dis. Fish Shellfish 2017, 189–222. [Google Scholar] [CrossRef]
- Ljungh, A.; Wadström, T. Lactic Acid Bacteria as Probiotics. Curr. Issues Intest. Microbiol. 2006, 7, 73–89. [Google Scholar]
- Park, Y.H.; Hamidon, F.; Rajangan, C.; Soh, K.P.; Gan, C.Y.; Lim, T.S.; Abdullah, W.N.W.; Liong, M.T. Application of probiotics for the production of safe and high-quality poultry meat. Korean J. Food Sci. Anim. Resour. 2016, 36, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Khalique, A.; Zeng, D.; Shoaib, M.; Wang, H.; Qing, X.; Rajput, D.S.; Pan, K.; Ni, X. Probiotics mitigating subclinical necrotic enteritis (SNE) as potential alternatives to antibiotics in poultry. AMB Express 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hartemink, R.; Domenech, V.; Rombouts, F. LAMVAB—A new selective medium for the isolation of lactobacilli from faeces. J. Microbiol. Methods 1997, 29, 77–84. [Google Scholar] [CrossRef]
- Zidour, M.; Chevalier, M.; Belguesmia, Y.; Cudennec, B.; Grard, T.; Drider, D.; Souissi, S.; Flahaut, C. Isolation and characterization of bacteria colonizing acartia tonsa copepod eggs and displaying antagonist effects against Vibrio anguillarum, Vibrio alginolyticus and other pathogenic strains. Front. Microbiol. 2017, 8, 1919. [Google Scholar] [CrossRef]
- Stenlid, J.; Karlsson, J.-O.; Högberg, N. Intraspecific genetic variation in Heterobasidion annosum revealed by amplification of minisatellite DNA. Mycol. Res. 1994, 98, 57–63. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Vachée, A.; Le Maréchal, C.; Salvat, G.; Drider, D. Antibiotic resistance, genome analysis and further safe traits of Clostridium perfringens ICVB082; A strain capable of producing an inhibitory compound directed only against a closely related pathogenic strain. Anaerobe 2020, 62, 102177. [Google Scholar] [CrossRef]
- Dec, M.; Puchalski, A.; Urban-Chmiel, R.; Wernicki, A. Screening of Lactobacillus strains of domestic goose origin against bacterial poultry pathogens for use as probiotics. Poult. Sci. 2014, 93, 2464–2472. [Google Scholar] [CrossRef]
- Bendali, F.; Hébraud, M.; Sadoun, D. Anti-bacterial and anti-adherence activities of a probiotic strain of Lactobacillus paracasei subsp. paracasei against Listeria monocytogenes. Int. J. Appl. Microbiol. Biotechnol. Res. 2014, 2, 52. [Google Scholar]
- Caly, D.L.; Chevalier, M.; Flahaut, C.; Cudennec, B.; Al Atya, A.K.; Chataigné, G.; D’Inca, R.; Auclair, E.; Drider, D. The safe enterocin DD14 is a leaderless two-peptide bacteriocin with anti-Clostridium perfringens activity. Int. J. Antimicrob. Agents 2017, 49, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Versalovic, J. Probiotic Lactobacillus reuteri biofilms produce antimicrobial and anti-inflammatory factors. BMC Microbiol. 2009, 9, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Haro, M.; Taggart, M.; Green, A.J.; Mateo, R. Avian digestive tract simulation to study the effect of grit geochemistry and food on pb shot bioaccessibility. Environ. Sci. Technol. 2009, 43, 9480–9486. [Google Scholar] [CrossRef] [PubMed]
- Musikasang, H.; Tani, A.; H-Kittikun, A.; Maneerat, S. Probiotic potential of lactic acid bacteria isolated from chicken gastrointestinal digestive tract. World J. Microbiol. Biotechnol. 2009, 25, 1337–1345. [Google Scholar] [CrossRef]
- Troxell, B.; Petri, N.; Daron, C.; Pereira, R.; Mendoza, M.; Hassan, H.M.; Koci, M.D. Poultry body temperature contributes to invasion control through reduced expression of Salmonella pathogenicity island 1 genes in Salmonella enterica Serovars Typhimurium and Enteritidis. Appl. Environ. Microbiol. 2015, 81, 8192–8201. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-Q.; Zhu, Y.-H.; Zhang, H.-F.; Yue, Y.; Cai, Z.-X.; Lü, Q.-P.; Zhang, L.; Weng, X.-G.; Zhang, F.-J.; Zhou, N.; et al. Risks Associated with high-dose Lactobacillus rhamnosus in an Escherichia coli model of piglet diarrhoea: Intestinal microbiota and immune imbalances. PLoS ONE 2012, 7, e40666. [Google Scholar] [CrossRef]
- Candela, M.; Perna, F.; Carnevali, P.; Vitali, B.; Ciati, R.; Gionchetti, P.; Rizzello, F.; Campieri, M.; Brigidi, P. Interaction of probiotic Lactobacillus and Bifidobacterium strains with human intestinal epithelial cells: Adhesion properties, competition against enteropathogens and modulation of IL-8 production. Int. J. Food Microbiol. 2008, 125, 286–292. [Google Scholar] [CrossRef]
- Wu, S.-B.; Rodgers, N.; Choct, M. Real-time pcr assay for Clostridium perfringens in broiler chickens in a challenge model of necrotic enteritis. Appl. Environ. Microbiol. 2011, 77, 1135–1139. [Google Scholar] [CrossRef] [Green Version]
- Pisano, M.B.; Viale, S.; Conti, S.; Fadda, M.E.; Deplano, M.; Melis, M.P.; Deiana, M.; Cosentino, S. Preliminary evaluation of probiotic properties of Lactobacillus strains isolated from sardinian dairy products. BioMed Res. Int. 2014, 2014, 286390. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zheng, Y.; Gao, H.; Xu, P.; Wang, M.; Li, A.; Miao, M.; Xie, X.; Deng, Y.; Zhou, H.; et al. Identification and characterization of Staphylococcus aureus strains with an incomplete hemolytic phenotype. Front. Cell. Infect. Microbiol. 2016, 6, 146. [Google Scholar] [CrossRef] [Green Version]
- Matuschek, E.; Brown, D.F.J.; Kahlmeter, G. Development of the EUCAST disk diffusion antimicrobial susceptibility testing method and its implementation in routine microbiology laboratories. Clin. Microbiol. Infect. 2014, 20, O255–O266. [Google Scholar] [CrossRef] [Green Version]
- Al Seraih, A.; Belguesmia, Y.; Cudennec, B.; Baah, J.; Drider, D. In silico and experimental data claiming safety aspects and beneficial attributes of the bacteriocinogenic strain Enterococcus faecalis B3A-B3B. Probiotics Antimicrob. Proteins 2017, 10, 456–465. [Google Scholar] [CrossRef]
- CLSI M45Ed3: Test Infrequently Isolated/Fastidious Bacteria. Available online: https://clsi.org/standards/products/microbiology/documents/m45/ (accessed on 24 August 2021).
- EFSA Guidance on the Characterisation of Microorganisms Used as Feed Additives or as Production Organisms. Autorité Européenne de Sécurité des Aliments. Available online: https://www.efsa.europa.eu/fr/efsajournal/pub/5206 (accessed on 24 August 2021).
- Chan, W.Y.; Hickey, E.E.; Khazandi, M.; Page, S.W.; Trott, D.J.; Hill, P.B. In vitro antimicrobial activity of narasin against common clinical isolates associated with canine otitis externa. Vet. Dermatol. 2018, 29, 149-e57. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Scientific Opinion on safety and efficacy of Cygro® 10G (maduramicin ammonium α) for chickens for fattening. EFSA J. 2011, 9, 1952. [Google Scholar] [CrossRef]
- McReynolds, J.L.; Byrd, J.A.; Anderson, R.C.; Moore, R.W.; Edrington, T.S.; Genovese, K.J.; Poole, T.L.; Kubena, L.F.; Nisbet, D.J. Evaluation of immunosuppressants and dietary mechanisms in an experimental disease model for necrotic enteritis. Poult. Sci. 2004, 83, 1948–1952. [Google Scholar] [CrossRef]
- Dromigny, E.; Bourrion, F.; Rugraf, Y.; Bolton, F.J.; Leden, N. New media for detection and counting of clostridia in foods. Lett. Appl. Microbiol. 1997, 24, 19–22. [Google Scholar] [CrossRef]
- Wenk, C.; Pfirter, H.; Bickel, H. Energetic aspects of feed conversion in growing pigs. Livest. Prod. Sci. 1980, 7, 483–495. [Google Scholar] [CrossRef]
- Prescott, J.F.; Sivendra, R.; Barnum, D.A. The use of bacitracin in the prevention and treatment of experimentally-induced necrotic enteritis in the chicken. Can. Vet. J. 1978, 19, 181–183. [Google Scholar]
- Cooper, K.K.; Songer, J.G. Necrotic enteritis in chickens: A paradigm of enteric infection by Clostridium perfringens type A. Anaerobe 2009, 15, 55–60. [Google Scholar] [CrossRef]
- Misiakos, E.P.; Bagias, G.; Patapis, P.; Sotiropoulos, D.; Kanavidis, P.; Machairas, A. Current concepts in the management of necrotizing fasciitis. Front. Surg. 2014, 1, 36. [Google Scholar] [CrossRef] [Green Version]
- Shiroff, A.M.; Herlitz, G.N.; Gracias, V.H. Necrotizing soft tissue infections. J. Intensiv. Care Med. 2012, 29, 138–144. [Google Scholar] [CrossRef]
- Wade, B.; Keyburn, A.L. The true cost of necrotic enteritis. Poultry World 2015, 31, 16–17. [Google Scholar]
- Landoni, M.; Albarellos, G. The use of antimicrobial agents in broiler chickens. Vet. J. 2015, 205, 21–27. [Google Scholar] [CrossRef]
- Mwangi, S.; Timmons, J.; Fitz-Coy, S.; Parveen, S. Characterization of Clostridium perfringens recovered from broiler chicken affected by necrotic enteritis. Poult. Sci. 2019, 98, 128–135. [Google Scholar] [CrossRef]
- Osman, K.; ElHariri, M. Antibiotic resistance of Clostridium perfringens isolates from broiler chickens in Egypt. Rev. Sci. Tech. de l’OIE 2013, 32, 841–850. [Google Scholar] [CrossRef]
- Adhikari, P.; Kiess, A.; Jha, R. An approach to alternative strategies to control avian coccidiosis and necrotic enteritis. J. Appl. Poult. Res. 2020, 29, 515–534. [Google Scholar] [CrossRef]
- Golić, N.; Veljović, K.; Popović, N.; Djokić, J.; Strahinić, I.; Mrvaljević, I.; Terzić-Vidojević, A. In vitro and in vivo antagonistic activity of new probiotic culture against Clostridium difficile and Clostridium perfringens. BMC Microbiol. 2017, 17, 108. [Google Scholar] [CrossRef] [Green Version]
- Ait-Seddik, H.; Bendali, F.; Cudennec, B.; Drider, D. Anti-pathogenic and probiotic attributes of Lactobacillus salivarius and Lactobacillus plantarum strains isolated from feces of Algerian infants and adults. Res. Microbiol. 2017, 168, 244–254. [Google Scholar] [CrossRef]
- Hor, Y.Y.; Liong, M.T. Use of extracellular extracts of lactic acid bacteria and bifidobacteria for the inhibition of dermatological pathogen Staphylococcus aureus. Dermatol. Sin. 2014, 32, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Vodnar, D.C.; Paucean, A.; Dulf, F.V.; Socaciu, C. HPLC Characterization of lactic acid formation and FTIR fingerprint of probiotic bacteria during fermentation processes. Not. Bot. Horti Agrobot. 2010, 38, 109–113. [Google Scholar] [CrossRef]
- Stanojević-Nikolić, S.; Dimić, G.; Mojović, L.; Pejin, J.; Djukić-Vuković, A.; Kocić-Tanackov, S. Antimicrobial activity of lactic acid against pathogen and spoilage microorganisms. J. Food Process. Preserv. 2015, 40, 990–998. [Google Scholar] [CrossRef]
- Wang, J.; Tang, H.; Zhang, C.; Zhao, Y.; Derrien, M.; Rocher, E.; van-Hylckama Vlieg, J.E.T.; Strissel, K.; Zhao, L.; Obin, M.; et al. Modulation of gut microbiota during probiotic-mediated attenuation of metabolic syndrome in high fat diet-fed mice. ISME J. 2015, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gómez, N.C.; Ramiro, J.M.P.; Quecan, B.X.V.; Franco, B.D.G.D.M. Use of potential probiotic lactic acid bacteria (lab) biofilms for the control of Listeria monocytogenes, Salmonella Typhimurium, and Escherichia coli O157:H7 biofilms formation. Front. Microbiol. 2016, 7, 863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarreche, M.P.; Castellano, P.; Vignolo, G. Evaluation of anti-Listeria meat borne Lactobacillus for biofilm formation on selected abiotic surfaces. Meat Sci. 2014, 96, 295–303. [Google Scholar] [CrossRef]
- Mare, L.; Wolfaardt, G.; Dicks, L. Adhesion of Lactobacillus plantarum 423 and Lactobacillus salivarius 241 to the intestinal tract of piglets, as recorded with fluorescent in situ hybridization (FISH), and production of plantaricin 423 by cells colonized to the ileum. J. Appl. Microbiol. 2006, 100, 838–845. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [Green Version]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in human health and diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef]
- Messaoudi, S.; Kergourlay, G.; Dalgalarrondo, M.; Choiset, Y.; Ferchichi, M.; Prevost, H.; Pilet, M.-F.; Chobert, J.-M.; Manai, M.; Dousset, X. Purification and characterization of a new bacteriocin active against Campylobacter produced by Lactobacillus salivarius SMXD51. Food Microbiol. 2012, 32, 129–134. [Google Scholar] [CrossRef]
- Barache, N.; Ladjouzi, R.; Belguesmia, Y.; Bendali, F.; Drider, D. Abundance of Lactobacillus plantarum strains with beneficial attributes in blackberries (Rubus sp.), fresh figs (Ficus carica), and prickly pears (Opuntia ficus-indica) grown and harvested in Algeria. Probiotics Antimicrob. Proteins 2020, 12, 1514–1523. [Google Scholar] [CrossRef]
- De Boever, P.; Wouters, R.; Verschaeve, L.; Berckmans, P.; Schoeters, G.; Verstraete, W. Protective effect of the bile salt hydrolase-active Lactobacillus reuteri against bile salt cytotoxicity. Appl. Microbiol. Biotechnol. 2000, 53, 709–714. [Google Scholar] [CrossRef]
- Martoni, C.J.; Labbé, A.; Ganopolsky, J.G.; Prakash, S.; Jones, M.L. Changes in bile acids, FGF-19 and sterol absorption in response to bile salt hydrolase active L. reuteri NCIMB 30242. Gut Microbes 2015, 6, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.M.T.; Pyle, B.H.; Camper, A.K. Specific and rapid enumeration of viable but nonculturable and viable-culturable gram-negative bacteria by using flow cytometry. Appl. Environ. Microbiol. 2010, 76, 5088–5096. [Google Scholar] [CrossRef] [Green Version]
- Er, S.; KOPARAL, A.T.; Kivanç, M. Cytotoxic effects of various lactic acid bacteria on Caco-2 cells. Turk. J. Boil. 2015, 39, 23–30. [Google Scholar] [CrossRef]
- Gueimonde, M.; Sánchez, B.; Reyes-Gavilán, C.G.D.L.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef]
- Ammor, M.S.; Flórez, A.B.; Mayo, B. Antibiotic resistance in non-enterococcal lactic acid bacteria and bifidobacteria. Food Microbiol. 2007, 24, 559–570. [Google Scholar] [CrossRef]
- Aquilina, G.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; De Knecht, J.; Dierick, N.A.; Gralak, M.A.; Gropp, J.; Halle, I.; Kroker, R.; et al. Scientific Opinion on Safety and Efficacy of Clinacox® 0.5% (Diclazuril) for Turkeys for Fattening. Available online: https://www.efsa.europa.eu/fr/efsajournal/pub/2115 (accessed on 16 June 2021).
- Dec, M.; Puchalski, A.; Stępień-Pyśniak, D.; Marek, A.; Urban-Chmiel, R. Susceptibility of chicken Lactobacillus bacteria to coccidiostats. J. Vet. Med Sci. 2020, 82, 333–336. [Google Scholar] [CrossRef] [Green Version]
- Marounek, M.; Rada, V. Susceptibility of poultry Lactobacilli to ionophore antibiotics. J. Vet. Med. Ser. B 1995, 42, 193–196. [Google Scholar] [CrossRef]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef]
- Song, J.; Xiao, K.; Ke, Y.L.; Jiao, L.F.; Hu, C.H.; Diao, Q.Y.; Shi, B.; Zou, X.T. Effect of a probiotic mixture on intestinal microflora, morphology, and barrier integrity of broilers subjected to heat stress. Poult. Sci. 2014, 93, 581–588. [Google Scholar] [CrossRef]
- Sureshkumar, S.; Lee, H.C.; Jung, S.K.; Kim, D.; Oh, K.B.; Yang, H.; Jo, Y.J.; Lee, H.S.; Lee, S.; Byun, S.J. Inclusion of Lactobacillus salivarius strain revealed a positive effect on improving growth performance, fecal microbiota and immunological responses in chicken. Arch. Microbiol. 2020, 203, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Ashayerizadeh, A.; Dabiri, N.; Mirzadeh, K.H.; Ghorbani, M.R. Effects of dietary inclusion of several biological feed additives on growth response of broiler chickens. J. Cell Anim. Biol. 2011, 5, 61–65. [Google Scholar]
- Nakphaichit, M.; Thanomwongwattana, S.; Phraephaisarn, C.; Sakamoto, N.; Keawsompong, S.; Nakayama, J.; Nitisinprasert, S. The effect of including Lactobacillus reuteri KUB-AC5 during post-hatch feeding on the growth and ileum microbiota of broiler chickens. Poult. Sci. 2011, 90, 2753–2765. [Google Scholar] [CrossRef] [PubMed]
- Mappley, L.J.; Tchórzewska, M.A.; Cooley, W.A.; Woodward, M.J.; La Ragione, R.M. Lactobacilli antagonize the growth, motility, and adherence of Brachyspira pilosicoli: A potential intervention against avian intestinal Spirochetosis. Appl. Environ. Microbiol. 2011, 77, 5402–5411. [Google Scholar] [CrossRef] [Green Version]
- Carter, A.; Adams, M.; La Ragione, R.; Woodward, M.J. Colonisation of poultry by Salmonella Enteritidis S1400 is reduced by combined administration of Lactobacillus salivarius 59 and Enterococcus faecium PXN-33. Vet. Microbiol. 2017, 199, 100–107. [Google Scholar] [CrossRef]
- Gérard, P.; Brézillon, C.; Quéré, F.; Salmon, A.; Rabot, S. Characterization of cecal microbiota and response to an orally administered Lactobacillus probiotic strain in the broiler chicken. J. Mol. Microbiol. Biotechnol. 2007, 14, 115–122. [Google Scholar] [CrossRef]
- Kizerwetter-Swida, M.; Binek, M. Protective effect of potentially probiotic Lactobacillus strain on infection with pathogenic bacteria in chickens. Pol. J. Vet. Sci. 2009, 12, 15–20. [Google Scholar]
- Li, Z.; Wang, W.; Liu, D.; Guo, Y. Effects of Lactobacillus acidophilus on the growth performance and intestinal health of broilers challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- La Ragione, R.; Narbad, A.; Gasson, M.; Woodward, M. In vivo characterization of Lactobacillus johnsonii FI9785 for use as a defined competitive exclusion agent against bacterial pathogens in poultry. Lett. Appl. Microbiol. 2004, 38, 197–205. [Google Scholar] [CrossRef]
- Cao, L.; Yang, X.; Li, Z.; Sun, F.; Wu, X.; Yao, J. Reduced lesions in chickens with Clostridium perfringens-induced necrotic enteritis by Lactobacillus fermentum 1.2029. Poult. Sci. 2012, 91, 3065–3071. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | Penicillin | Cefotaxime | Gentamicin | Vancomycin | Clindamycin | Ciprofloxacin | Tetracycline | Trimethoprim Sulfamethoxazole |
|---|---|---|---|---|---|---|---|---|
| L. reuteri ICVB416 | R (0) | S (45) | S (26) | R (0) | S (40) | S (36) | S (26) | S (21) |
| L. salivarius ICVB421 | R (0) | S (42) | S (28) | R (0) | S (46) | S (26) | S (26) | S (34) |
| L. salivarius ICVB430 | R (0) | S (42) | S (28) | R (0) | S (34) | S (26) | S (26) | R (0) |
| Active Agent | Dosage in Farms | MIC | ||
|---|---|---|---|---|
| L. reuteri ICVB416 | L. salivarius ICVB421 | L. salivarius ICVB430 | ||
| Monensin sodium | 60–125 | 2–4 | 1–2 | 2–4 |
| Narasin | 60–70 | 0.063–0.125 | 0.016–0.063 | 0.016 |
| Salinomycin | 30–70 | 0.5–1 | 0.125–0.25 | 0.25–0.5 |
| Maduramicin ammonium | 5 | 0.5–1 | 0.25 | 0.25–0.5 |
| Lasalocid A sodium | 75–125 | 0.25–0.5 | 1–2 | 0.125–0.5 |
| Diclazuril | 1 | >512 | >512 | >512 |
| Robenidine | 30–36 | 4–16 | 8–16 | 4–16 |
| Average Weight/Bird (g) | Feed Consumption (g) | FCR | ||||
|---|---|---|---|---|---|---|
| D10 | D14 | D0-D10 | D10-D14 | D0-D10 | D10-D14 | |
| Control (No L. addition) | 320.4 ± 12.8 B | 547.4 ± 22.2 AB | 311.50 ± 9.9 AB | 280.21 ± 10.3 | 1.12 ± 0.02 AB | 1.24 ± 0.04 AB |
| L. reuteri ICVB 416 | 317.9 ± 10.7 B | 544.8 ± 24.8 AB | 305,50 ± 12,3 AB | 275.63 ± 12.3 | 1.12 ± 0.02 AB | 1.22 ± 0.05 B |
| L. salivarius ICVB 421 | 329.8 ± 8.4 AB | 564.6 ± 17.9 A | 314.17 ± 10.1 AB | 285.42 ± 9.0 | 1.13 ± 0.01 AB | 1.22 ± 0.04 B |
| L. salivarius ICVB 430 | 326.9 ± 17.4 AB | 555.4 ± 22.9 AB | 316.33 ± 14.7 A | 282.08 ± 12.6 | 1.13 ± 0.02 A | 1.23 ± 0.04 AB |
| ICVB 416 + ICVB 421 | 320.2 ± 9.2 AB | 543.4 ± 20.1 AB | 303.17 ± 9.4 B | 273.75 ± 7.8 | 1.11 ± 0.03 AB | 1.23 ± 0.02 AB |
| ICVB 416 + ICVB 430 | 334.1 ± 15.06 A | 566.5 ± 20.0 A | 314.67 ± 9.1 AB | 284.38 ± 9.8 | 1.10 ± 0.03 B | 1.22 ± 0.04 AB |
| ICVB 416 + ICVB 421 + ICVB 430 | 320.9 ± 12.3 AB | 536.7 ± 16.0 B | 304.79 ± 8.4 AB | 274.58 ± 8.7 | 1.13 ± 0.04 AB | 1.27 ± 0.05 A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Padgett, C.; Bailey, C.; Gancel, F.; Drider, D. Protective Effects of Novel Lactobacillaceae Strains Isolated from Chicken Caeca against Necrotic Enteritis Infection: In Vitro and In Vivo Evidences. Microorganisms 2022, 10, 152. https://doi.org/10.3390/microorganisms10010152
Vieco-Saiz N, Belguesmia Y, Raspoet R, Auclair E, Padgett C, Bailey C, Gancel F, Drider D. Protective Effects of Novel Lactobacillaceae Strains Isolated from Chicken Caeca against Necrotic Enteritis Infection: In Vitro and In Vivo Evidences. Microorganisms. 2022; 10(1):152. https://doi.org/10.3390/microorganisms10010152
Chicago/Turabian StyleVieco-Saiz, Nuria, Yanath Belguesmia, Ruth Raspoet, Eric Auclair, Connor Padgett, Christopher Bailey, Frédérique Gancel, and Djamel Drider. 2022. "Protective Effects of Novel Lactobacillaceae Strains Isolated from Chicken Caeca against Necrotic Enteritis Infection: In Vitro and In Vivo Evidences" Microorganisms 10, no. 1: 152. https://doi.org/10.3390/microorganisms10010152
APA StyleVieco-Saiz, N., Belguesmia, Y., Raspoet, R., Auclair, E., Padgett, C., Bailey, C., Gancel, F., & Drider, D. (2022). Protective Effects of Novel Lactobacillaceae Strains Isolated from Chicken Caeca against Necrotic Enteritis Infection: In Vitro and In Vivo Evidences. Microorganisms, 10(1), 152. https://doi.org/10.3390/microorganisms10010152