
Life Cycle and Transmission of Cyclospora cayetanensis: Knowns and Unknowns



Abstract
1. Introduction
2. Etiology
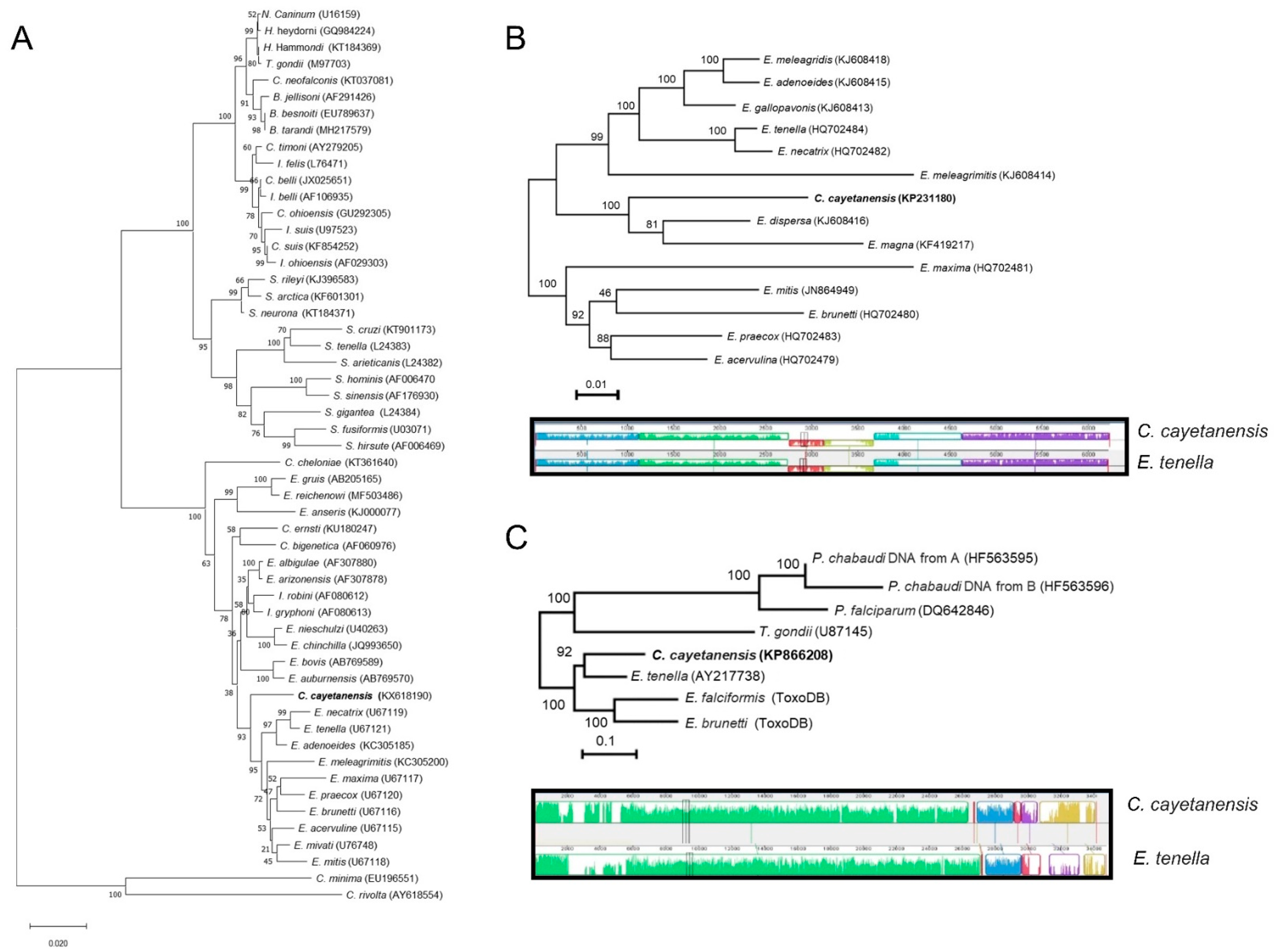
3. Taxonomy
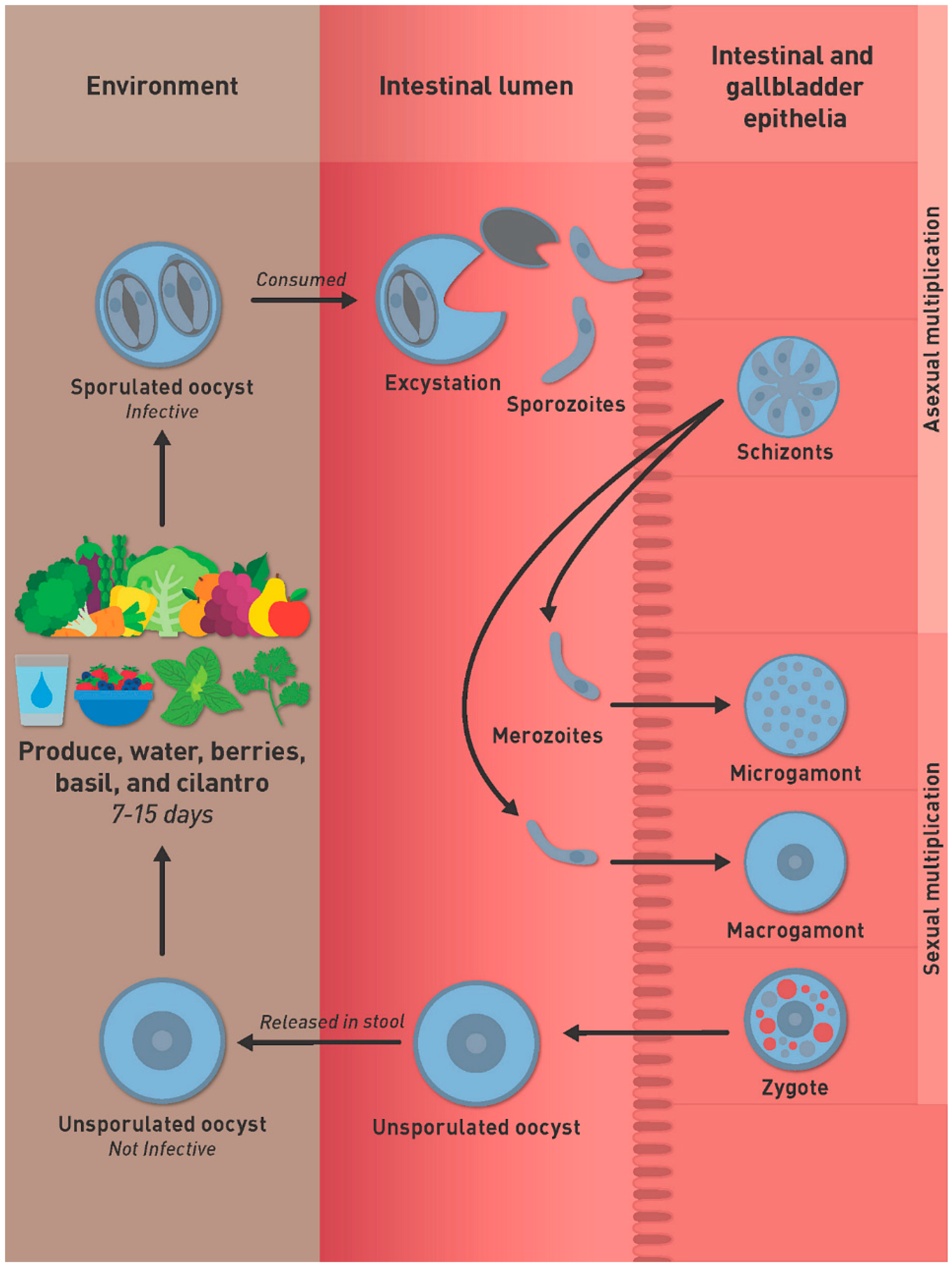
4. Life Cycle of C. cayetanensis
4.1. Oocysts
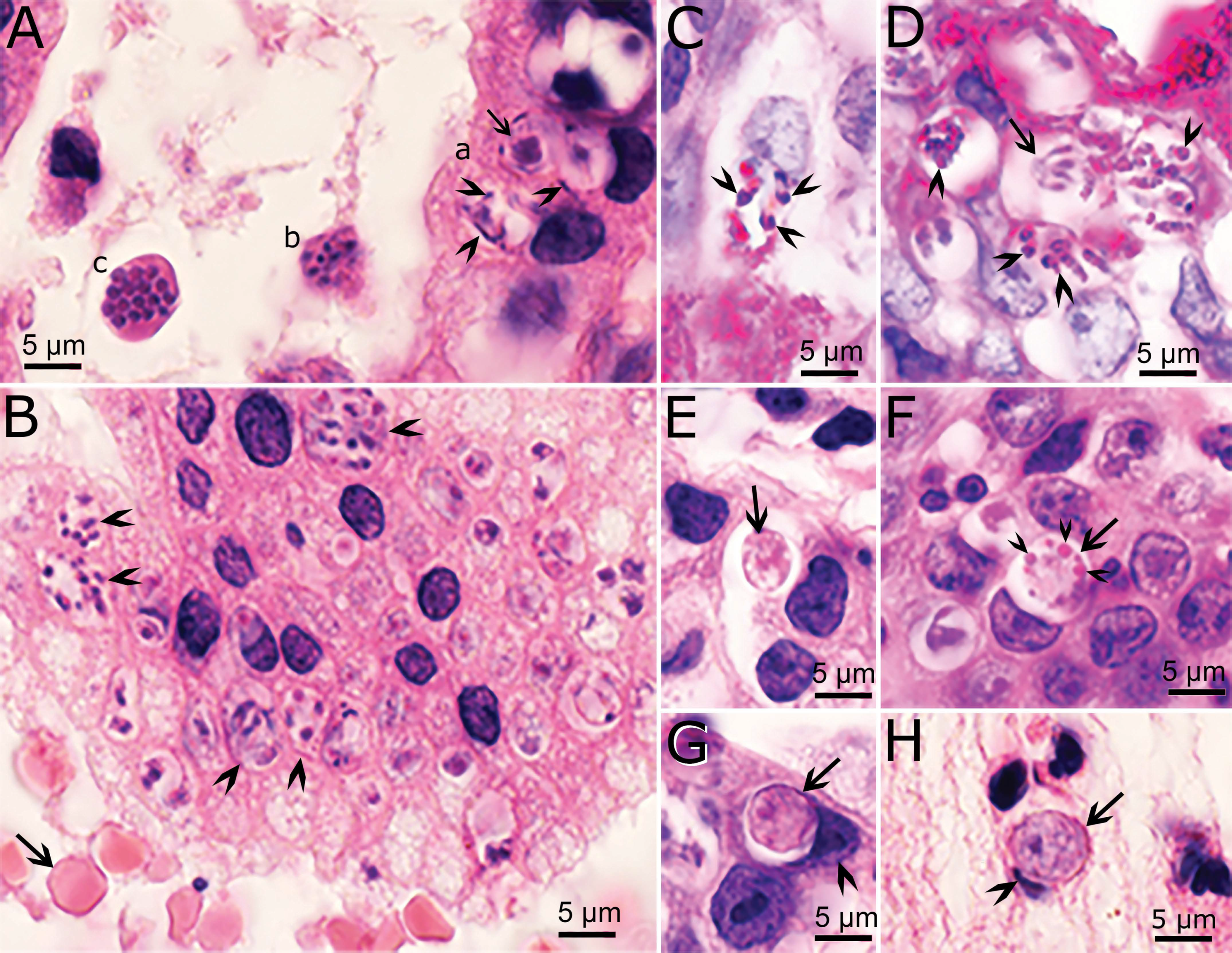
4.2. Asexual Multiplication
Asexual Multiplication in Related Coccidia
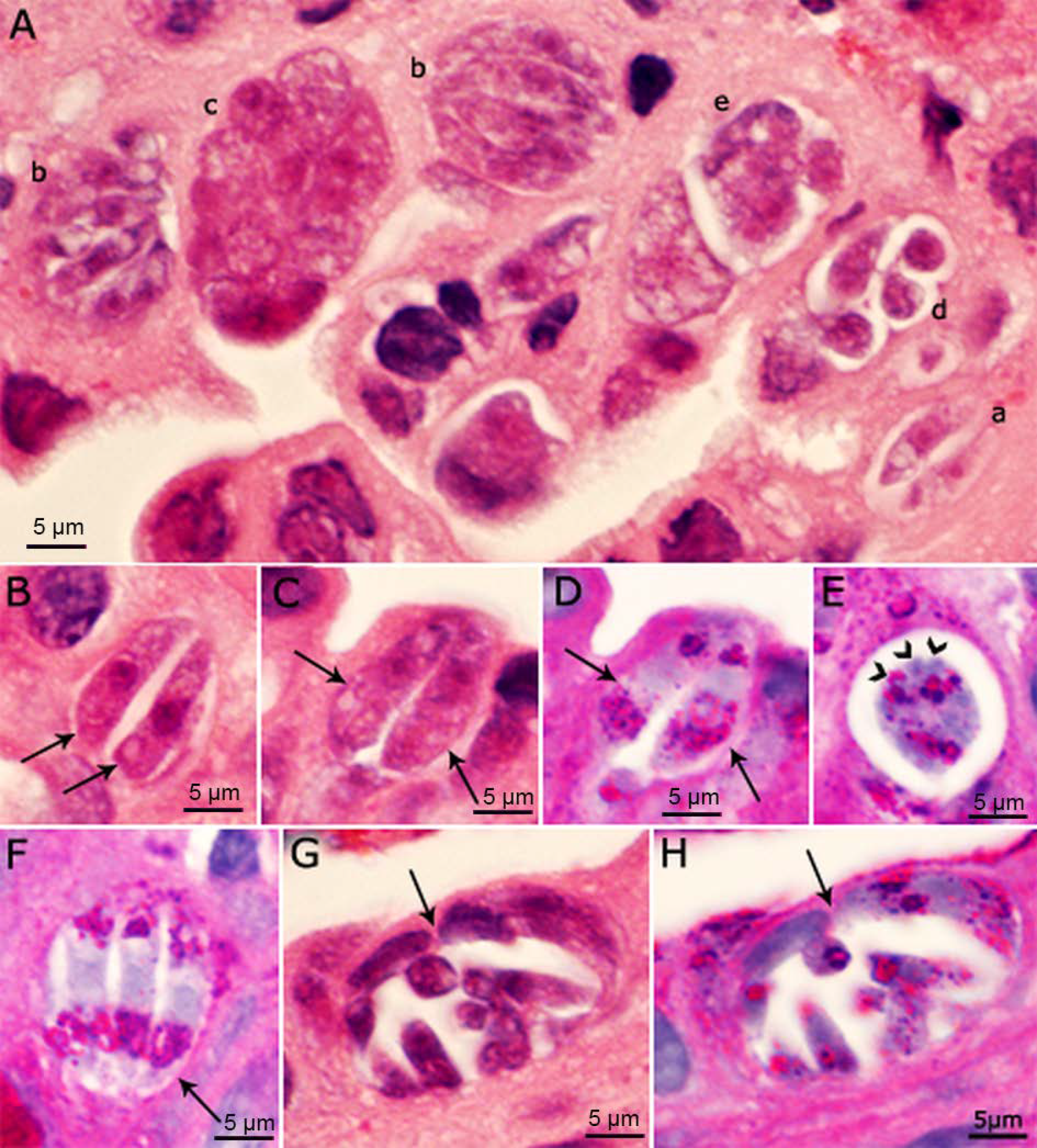
4.3. Sexual Stages
5. Need for Surrogates for Studying C. cayetanensis
6. Molecular Tools and Comparative Genomics
6.1. Small-Subunit rDNA Sequence
6.2. Organellar Genomes
6.3. Nuclear Genome
6.4. Developmentally-Regulated Genes
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Almeria, S.; Cinar, H.N.; Dubey, J.P. Cyclospora cayetanensis and cyclosporiasis: An update. Microorganisms 2019, 7, 317. [Google Scholar] [CrossRef]
- Mathison, B.A.; Pritt, B.S. Cyclosporiasis-updates on clinical presentation, pathology, clinical diagnosis, and treatment. Microorganisms 2021, 9, 1863. [Google Scholar] [CrossRef] [PubMed]
- Giangaspero, A.; Gasser, R.B. Human cyclosporiasis. Lancet Infect. Dis. 2019, 19, e226–e236. [Google Scholar] [CrossRef]
- Li, J.; Cui, Z.; Qi, M.; Zhang, L. Advances in cyclosporiasis diagnosis and therapeutic intervention. Front. Cell Infect. Microbiol. 2020, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Hadjilouka, A.; Tsaltas, D. Cyclospora cayetanensis-major outbreaks from ready to eat fresh fruits and vegetables. Foods 2020, 9, 1703. [Google Scholar] [CrossRef] [PubMed]
- Almeria, S.C.; Cinar, H.N.; Dubey, J.P. Coccidiosis in humans. In Coccidiosis in Livestock, Poultry, Companion Animals, and Humans; Dubey, J.P., Ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2020; pp. 268–312. [Google Scholar]
- Ortega, Y.R.; Gilman, R.H.; Sterling, C.R. A new coccidian parasite (Apicomplexa: Eimeriidae) from humans. J. Parasitol. 1994, 80, 625–629. [Google Scholar] [CrossRef]
- Solarczyk, P. Host range of Cyclospora species: Zoonotic implication. Acta Protozool. 2021, 60, 13–20. [Google Scholar] [CrossRef]
- Eberhard, M.L.; Ortega, Y.R.; Hanes, D.E.; Nace, E.K.; Do, R.Q.; Robl, M.G.; Won, K.Y.; Gavidia, C.; Sass, N.L.; Mansfield, K.; et al. Attempts to establish experimental Cyclospora cayetanensis infection in laboratory animals. J. Parasitol. 2000, 86, 577–582. [Google Scholar] [CrossRef]
- Ortega, Y.R.; Nagle, R.; Gilman, R.H.; Watanabe, J.; Miyagui, J.; Quispe, H.; Kanagusuku, P.; Roxas, C.; Sterling, C.R. Pathologic and clinical findings in patients with cyclosporiasis and a description of intracellular parasite life-cycle stages. J. Infect. Dis. 1997, 176, 1584–1589. [Google Scholar] [CrossRef]
- Dubey, J.P.; Almeria, S.; Mowery, J.; Fortes, J. Endogenous developmental cycle of the human coccidian Cyclospora cayetanensis. J. Parasitol. 2020, 106, 295–307. [Google Scholar] [CrossRef]
- Dubey, J.P.; Lindsay, D.S.; Jenkins, M.C.; Bauer, C. Biology of intestinal coccidia. In Coccidiosis in Livestock, Poultry, Companion Animals, and Humans; Dubey, J.P., Ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2020; pp. 1–36. [Google Scholar]
- Thompson, P.; Rosenthal, B.M. Phylogeny of coccidian parasites. In Coccidiosis in Livestock, Poultry, Companion Animals, and Humans; Dubey, J.P., Ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2020; pp. 37–41. [Google Scholar]
- Dubey, J.P.; Almeria, S. Cystoisospora belli infections in humans: The past 100 years. Parasitology 2019, 146, 1490–1527. [Google Scholar] [CrossRef]
- Dubey, J.P.; Evason, K.J.; Walther, Z. Endogenous development of Cystoisospora belli in intestinal and biliary epithelium of humans. Parasitology 2019, 146, 865–872. [Google Scholar] [CrossRef]
- Resende, D.V.; Pedrosa, A.L.; Correia, D.; Cabrine-Santos, M.; Lages-Silva, E.; Meira, W.S.; Oliveira-Silva, M.B. Polymorphisms in the 18S rDNA gene of Cystoisospora belli and clinical features of cystoisosporosis in HIV-infected patients. Parasitol. Res. 2011, 108, 679–685. [Google Scholar] [CrossRef]
- Taniuchi, M.; Verweij, J.J.; Sethabutr, O.; Bodhidatta, L.; Garcia, L.; Maro, A.; Kumburu, H.; Gratz, J.; Kibiki, G.; Houpt, E.R. Multiplex polymerase chain reaction method to detect Cyclospora, Cystoisospora, and Microsporidia in stool samples. Diagn. Microbiol. Infect. Dis 2011, 71, 386–390. [Google Scholar] [CrossRef]
- Woon, S.A.; Yang, R.; Ryan, U.; Boan, P.; Prentice, D. Chronic Cystoisospora belli infection in an immunocompetent Myanmar refugee—Microscopy is not sensitive enough. BMC Infect. Dis. 2016, 16, 221. [Google Scholar] [CrossRef] [PubMed]
- Tucker, M.S.; O’Brien, C.N.; Jenkins, M.C.; Rosenthal, B.M. Dynamically expressed genes provide candidate viability biomarkers in a model coccidian. PLoS ONE 2021, 16, e0258157. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Q.; Leasi, F.; Obertegger, U.; Kieneke, A.; Barraclough, T.G.; Fontaneto, D. The widely used small subunit 18S rDNA molecule greatly underestimates true diversity in biodiversity surveys of the meiofauna. Proc. Natl. Acad. Sci. USA 2012, 109, 16208–16212. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, M.L.; da Silva, A.J.; Lilley, B.G.; Pieniazek, N.J. Morphologic and molecular characterization of new Cyclospora species from Ethiopian monkeys: C. cercopitheci sp.n., C. colobi sp. n., and C. papionis sp. n. Emerg. Infect. Dis. 1999, 5, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.M.; Ortega, Y.; Simpson, S.; Kerdahi, K. Genetic characterization of human-pathogenic Cyclospora cayetanensis parasites from three endemic regions at the 18S ribosomal RNA locus. Infect. Genet. Evol. 2014, 22, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Cicek, M.; Yildirim, I.H.; Tas Cengiz, Z.; Karaman, U. Single-strand conformation polymorphism-based genetic characterization of the Cyclospora cayetanensis strains collected from different provinces in Turkey. Ann. Agric. Environ. Med. 2021, 28, 267–270. [Google Scholar] [CrossRef]
- Murphy, H.R.; Cinar, H.N.; Gopinath, G.; Noe, K.E.; Chatman, L.D.; Miranda, N.E.; Wetherington, J.H.; Neal-McKinney, J.; Pires, G.S.; Sachs, E.; et al. Interlaboratory validation of an improved method for detection of Cyclospora cayetanensis in produce using a real-time PCR assay. Food Microbiol. 2018, 69, 170–178. [Google Scholar] [CrossRef]
- Li, N.; Ye, J.; Arrowood, M.J.; Ma, J.; Wang, L.; Xu, H.; Feng, Y.; Xiao, L. Identification and morphologic and molecular characterization of Cyclospora macacae n. sp. from rhesus monkeys in China. Parasitol. Res. 2015, 114, 1811–1816. [Google Scholar] [CrossRef][Green Version]
- Lim, L.; McFadden, G.I. The evolution, metabolism and functions of the apicoplast. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 749–763. [Google Scholar] [CrossRef]
- Greiner, S.; Sobanski, J.; Bock, R. Why are most organelle genomes transmitted maternally? Bioessays 2015, 37, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Guo, Y.; Zhang, L.; Rowe, L.A.; Roellig, D.M.; Frace, M.A.; Li, N.; Liu, S.; Feng, Y.; Xiao, L. Genetic similarities between Cyclospora cayetanensis and cecum-infecting avian Eimeria spp. in apicoplast and mitochondrial genomes. Parasit. Vectors 2015, 8, 358. [Google Scholar] [CrossRef]
- Ogedengbe, M.E.; Qvarnstrom, Y.; da Silva, A.J.; Arrowood, M.J.; Barta, J.R. A linear mitochondrial genome of Cyclospora cayetanensis (Eimeriidae, Eucoccidiorida, Coccidiasina, Apicomplexa) suggests the ancestral start position within mitochondrial genomes of eimeriid coccidia. Int. J. Parasitol. 2015, 45, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Cinar, H.N.; Gopinath, G.; Jarvis, K.; Murphy, H.R. The complete mitochondrial genome of the foodborne parasitic pathogen Cyclospora cayetanensis. PLoS ONE 2015, 10, e0128645. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, F.S.; Barta, J.R.; Whale, J.; Hofstetter, J.N.; Casillas, S.; Barratt, J.; Talundzic, E.; Arrowood, M.J.; Qvarnstrom, Y. Mitochondrial junction region as genotyping marker for Cyclospora cayetanensis. Emerg. Infect. Dis. 2019, 25, 1314–1319. [Google Scholar] [CrossRef]
- Gopinath, G.R.; Cinar, H.N.; Murphy, H.R.; Durigan, M.; Almeria, M.; Tall, B.D.; DaSilva, A.J. A hybrid reference-guided de novo assembly approach for generating Cyclospora mitochondrion genomes. Gut. Pathog. 2018, 10, 15. [Google Scholar] [CrossRef]
- Cinar, H.N.; Qvarnstrom, Y.; Wei-Pridgeon, Y.; Li, W.; Nascimento, F.S.; Arrowood, M.J.; Murphy, H.R.; Jang, A.; Kim, E.; Kim, R.; et al. Comparative sequence analysis of Cyclospora cayetanensis apicoplast genomes originating from diverse geographical regions. Parasit. Vectors 2016, 9, 611. [Google Scholar] [CrossRef]
- Liu, S.; Wang, L.; Zheng, H.; Xu, Z.; Roellig, D.M.; Li, N.; Frace, M.A.; Tang, K.; Arrowood, M.J.; Moss, D.M.; et al. Comparative genomics reveals Cyclospora cayetanensis possesses coccidia-like metabolism and invasion components but unique surface antigens. BMC Genom. 2016, 17, 316. [Google Scholar] [CrossRef] [PubMed]
- Qvarnstrom, Y.; Wei-Pridgeon, Y.; Li, W.; Nascimento, F.S.; Bishop, H.S.; Herwaldt, B.L.; Moss, D.M.; Nayak, V.; Srinivasamoorthy, G.; Sheth, M.; et al. Draft genome sequences from Cyclospora cayetanensis oocysts purified from a human stool sample. Genome Announc. 2015, 3, e01324-15. [Google Scholar] [CrossRef] [PubMed]
- Baptista, R.P.; Li, Y.; Sateriale, A.; Brooks, K.L.; Tracey, A.; Sanders, M.J.; Ansell, B.R.E.; Jex, A.R.; Cooper, G.W.; Smith, E.D.; et al. Long-read assembly and comparative evidence-based reanalysis of Cryptosporidium genome sequences reveals expanded transporter repertoire and duplication of entire chromosome ends including subtelomeric regions. Genome Res. 2021, 32, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.J.; Hall, N.; Fung, E.; White, O.; Berriman, M.; Hyman, R.W.; Carlton, J.M.; Pain, A.; Nelson, K.E.; Bowman, S.; et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 2002, 419, 498–511. [Google Scholar] [CrossRef] [PubMed]
- Brayton, K.A.; Lau, A.O.; Herndon, D.R.; Hannick, L.; Kappmeyer, L.S.; Berens, S.J.; Bidwell, S.L.; Brown, W.C.; Crabtree, J.; Fadrosh, D.; et al. Genome sequence of Babesia bovis and comparative analysis of apicomplexan hemoprotozoa. PLoS Pathog. 2007, 3, 1401–1413. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, H.; Khan, A.; Behnke, M.S.; Namasivayam, S.; Swapna, L.S.; Hadjithomas, M.; Karamycheva, S.; Pinney, D.; Brunk, B.P.; Ajioka, J.W.; et al. Local admixture of amplified and diversified secreted pathogenesis determinants shapes mosaic Toxoplasma gondii genomes. Nat. Commun. 2016, 7, 10147. [Google Scholar] [CrossRef]
- Blazejewski, T.; Nursimulu, N.; Pszenny, V.; Dangoudoubiyam, S.; Namasivayam, S.; Chiasson, M.A.; Chessman, K.; Tonkin, M.; Swapna, L.S.; Hung, S.S.; et al. Systems-based analysis of the Sarcocystis neurona genome identifies pathways that contribute to a heteroxenous life cycle. mBio 2015, 6, e02445-14. [Google Scholar] [CrossRef]
- Khan, A.; Fujita, A.W.; Randle, N.; Regidor-Cerrillo, J.; Shaik, J.S.; Shen, K.; Oler, A.J.; Quinones, M.; Latham, S.M.; Akanmori, B.D.; et al. Global selective sweep of a highly inbred genome of the cattle parasite Neospora caninum. Proc. Natl. Acad. Sci. USA 2019, 116, 22764–22773. [Google Scholar] [CrossRef]
- Berna, L.; Marquez, P.; Cabrera, A.; Greif, G.; Francia, M.E.; Robello, C. Reevaluation of the Toxoplasma gondii and Neospora caninum genomes reveals misassembly, karyotype differences, and chromosomal rearrangements. Genome Res. 2021, 31, 823–833. [Google Scholar] [CrossRef]
- Xia, J.; Venkat, A.; Bainbridge, R.E.; Reese, M.L.; Le Roch, K.G.; Ay, F.; Boyle, J.P. Third-generation sequencing revises the molecular karyotype for Toxoplasma gondii and identifies emerging copy number variants in sexual recombinants. Genome Res. 2021, 31, 834–851. [Google Scholar] [CrossRef]
- Reid, A.J.; Blake, D.P.; Ansari, H.R.; Billington, K.; Browne, H.P.; Bryant, J.; Dunn, M.; Hung, S.S.; Kawahara, F.; Miranda-Saavedra, D.; et al. Genomic analysis of the causative agents of coccidiosis in domestic chickens. Genome Res. 2014, 24, 1676–1685. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Cryptosporidium parvum | Plasmodium falciparum | Babesia bovis | Sarcocystis neurona | Neospora caninum | Hammondia hammondi | Toxoplasma gondii | Eimeria tenella | Cyclospora cayetanensis |
|---|---|---|---|---|---|---|---|---|---|
| Reference | [34,36] | [34,37] | [34,38] | [39,40] | [39,41,42] | [39] | [39,42,43] | [34,44] | [34,35] |
| No. of chromosomes | 8 | 14 | 4 | NA | 13 | 14 | 13 | 14 | NA |
| Estimated size (Mb) | ~9 | ~23 | 8 | ~127 | ~62 | ~65 | ~65 | 52 | 44 |
| GC content (%) | 30.3 | 19.4 | 41.8 | 51.5 | 54.8 | 52.5 | 52.2 | 52.5 | 51.8 |
| No. of protein-coding genes | 3897 | 5542 | 3671 | 7093 | 7540 | 8004 | 8322 | 8597 | 7457 |
| Mean length of protein-coding genes (bp) | 1799 | 2271 | 1514 | 9121 | 4872 | 4868 | 4778 | 1518 | 1599 |
| Apicoplast genome size (kb) | NA | ~34 | ~33 | ~35 | ~35 | ~35 | ~35 | ~35 | ~34 |
| Mitochondrial genome size (bp) | NA | ~6 | ~6 | NA | ~6 | ~6 | ~6 | ~6 | ~6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubey, J.P.; Khan, A.; Rosenthal, B.M. Life Cycle and Transmission of Cyclospora cayetanensis: Knowns and Unknowns. Microorganisms 2022, 10, 118. https://doi.org/10.3390/microorganisms10010118
Dubey JP, Khan A, Rosenthal BM. Life Cycle and Transmission of Cyclospora cayetanensis: Knowns and Unknowns. Microorganisms. 2022; 10(1):118. https://doi.org/10.3390/microorganisms10010118
Chicago/Turabian StyleDubey, Jitender P., Asis Khan, and Benjamin M. Rosenthal. 2022. "Life Cycle and Transmission of Cyclospora cayetanensis: Knowns and Unknowns" Microorganisms 10, no. 1: 118. https://doi.org/10.3390/microorganisms10010118
APA StyleDubey, J. P., Khan, A., & Rosenthal, B. M. (2022). Life Cycle and Transmission of Cyclospora cayetanensis: Knowns and Unknowns. Microorganisms, 10(1), 118. https://doi.org/10.3390/microorganisms10010118