Genetic Diversity and Sequence Polymorphism of Two Genes Encoding Theileria parva Antigens Recognized by CD8+ T Cells among Vaccinated and Unvaccinated Cattle in Malawi

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Theileria parva Screening
2.1.1. Sequencing Analysis
2.1.2. Tp1 Gene
2.1.3. Tp2 Gene
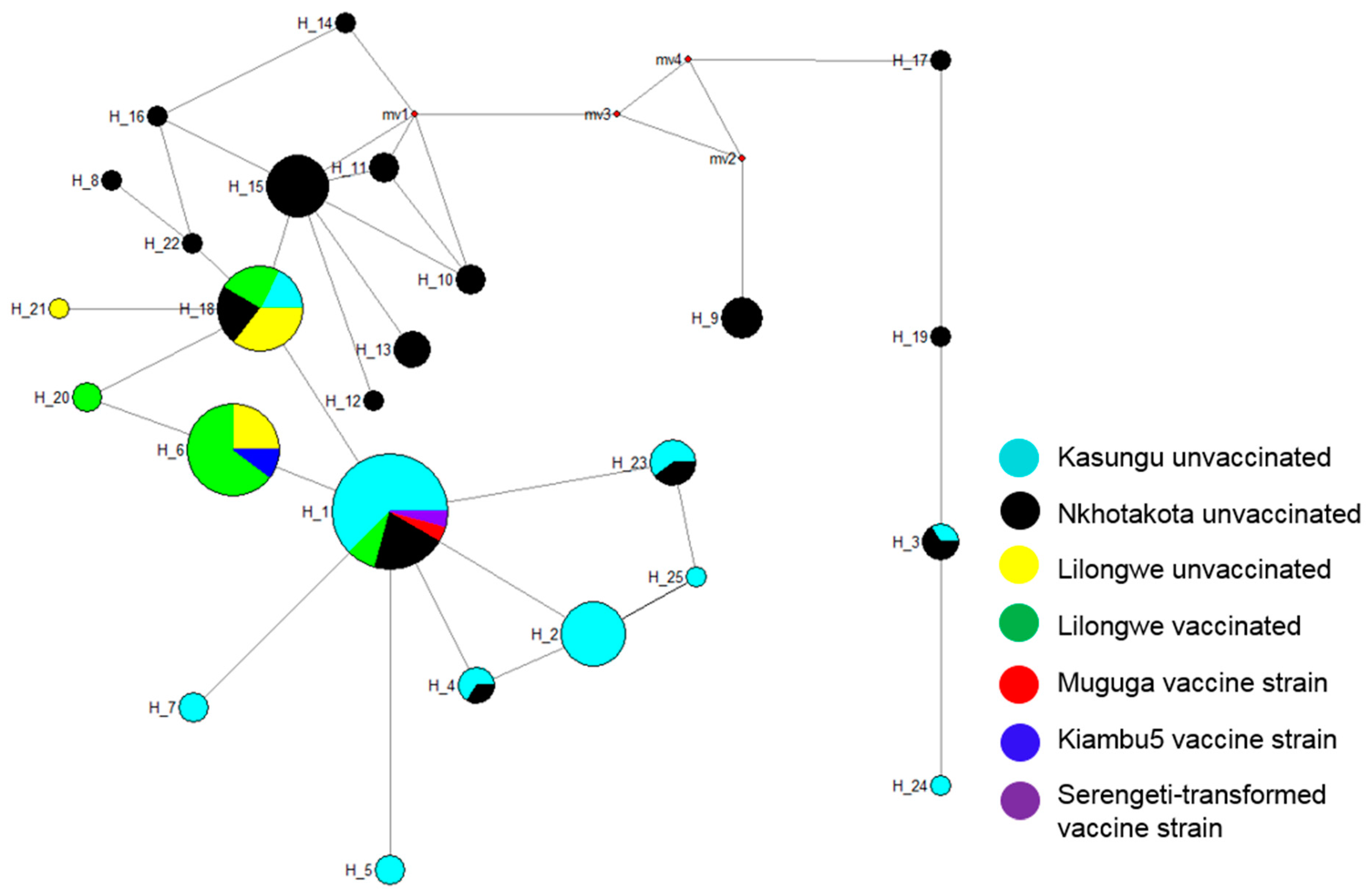
2.1.4. Phylogenetic Analysis of Tp1 and Tp2 Sequences from T. parva in Malawi
3. Discussion
4. Materials and Methods
4.1. Ethical Consideration
4.2. Study Area and Sampling.
4.3. DNA Extraction.
4.4. PCR and Sequencing
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Geysen, D.; Bishop, R.; Skilton, R.; Dolan, T.T.; Morzaria, S. Molecular epidemiology of Theileria parva in the field. Trop. Med. Int. Heal. 1999, 4, A21–A27. [Google Scholar] [CrossRef] [PubMed]
- Nene, V.; Kiara, H.; Lacasta, A.; Pelle, R.; Svitek, N.; Steinaa, L. The biology of Theileria parva and control of East Coast fever – Current status and future trends. Ticks Tick-borne Dis. 2016, 7, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Morrison, W.I. Progress towards understanding the immunobiology of Theileria parasites. Parasitology 2009, 136, 1415–1426. [Google Scholar] [CrossRef] [PubMed]
- Norval, R.A.I.; Perry, B.D.; Young, A.S. The Epidemiology of Theileria in Africa; Academic Press Limited: London, UK, 1992; pp. 279–301. ISBN 0-12-521-740-4. [Google Scholar]
- Dobbelaere, D.; Heussler, V. Transformation of Leukocytes byTheileria parvaandT. annulata. Annu. Rev. Microbiol. 1999, 53, 1–42. [Google Scholar] [CrossRef]
- Tretina, K.; Gotia, H.T.; Mann, D.J.; Silva, J.C. Theileria-transformed bovine leukocytes have cancer hallmarks. Trends Parasitol. 2015, 31, 306–314. [Google Scholar] [CrossRef]
- De Deken, R.; Martin, V.; Saido, A.; Madder, M.; Brandt, J.; Geysen, D. An outbreak of East Coast Fever on the Comoros: A consequence of the import of immunised cattle from Tanzania? Veter- Parasitol. 2007, 143, 245–253. [Google Scholar] [CrossRef]
- Malak, A.; Mpoke, L.; Banak, J.; Muriuki, S.; Skilton, R.; Odongo, D.; Sunter, J.D.; Kiara, H. Prevalence of livestock diseases and their impact on livelihoods in Central Equatoria State, southern Sudan. Prev. Veter- Med. 2012, 104, 216–223. [Google Scholar] [CrossRef]
- Hoogstraal, H. African Ixodoidea. Ticks of the Sudan (with Special Reference to Equatoria Province and with Preliminary Reviews of the Genera Boophilus, Margaropus, and Hyalomma); Department of Navy, Bureau of Medicine and Surgery: Washington, DC, USA, 1956; p. 1101. [Google Scholar]
- Silatsa, B.A.; Simo, G.; Githaka, N.; Kamga, R.; Oumarou, F.; Tiambo, C.K.; Machuka, E.; Domelevo, J.; Odongo, D.; Bishop, R.; et al. First detection of Theileria parva in cattle from Cameroon in the absence of the main tick vector Rhipicephalus appendiculatus. Transbound. Emerg. Dis. 2020, 67 (Suppl. 1), 68–78. [Google Scholar] [CrossRef]
- Guilbride, P.D.; Opwata, B. Observations on the resistance of Jersey-Nganda calves to east coast fever (Theileria parva). Bull. Epizoot. Dis. Afr. Bull. Epizoot. Afr. 1963, 11, 289–298. [Google Scholar]
- Oura, C.A.L.; Bishop, R.; Asiimwe, B.B.; Spooner, P.; Lubega, G.W.; Tait, A. Theileria parva live vaccination: Parasite transmission, persistence and heterologous challenge in the field. Parasitology 2007, 134, 1205–1213. [Google Scholar] [CrossRef]
- Chinombo, D.O.; Mzoma, F.J.; Musisi, F.L. East Coast Fever in Malawi (1983–1987). Theileriosis in Eastern, Central and Southern Africa. In Proceedings of a Workshop on East Coast Fever Immunization, Lilongwe, Malawi, 20–22 September 1988; Dolan, T.T., Ed.; ILRAD: Nairobi, Kenya, 1988; pp. 12–16. [Google Scholar]
- Young, A.S. The epidemiology of Theileriosis in East Africa. In Advances in the Control of Theileriosis; Irvin, A.D., Cunningham, M.P., Young, A.S., Eds.; Springer: Dordrecht, Netherland, 1981; pp. 38–55. [Google Scholar] [CrossRef]
- Moll, G.; Lohding, A.; Young, A. Epidemiology of theilerioses in the Trans-Mara division, Kenya: Husbandry and disease background and preliminary investigations on theilerioses in calves. Prev. Veter- Med. 1984, 2, 801–831. [Google Scholar] [CrossRef]
- Moll, G.; Lohding, A.; Young, A.; Leitch, B. Epidemiology of theileriosis in calves in an endemic area of Kenya. Veter- Parasitol. 1986, 19, 255–273. [Google Scholar] [CrossRef]
- GALVmed. GALVmed & East Coast Fever. The CTTBD ECF Vaccine: Greater opportunity to control East Coast fever and other tick-borne diseases in Africa. Global Alliance for Livestock Veterinary Medicines, Pentlands SciencePark, Bush Loan, Edinburgh EH26 OPZ, UK. 2010. Available online: https://www.fanrpan.org/sites/default/files/GALVmed_newsletter_November2010.pdf (accessed on 13 August 2019).
- Knudsen, P.B. Final Report of Activities of the Malawian-Danish Cattle Survey in Malawi; DANIDA/Government of Malawi: Lilongwe, Malawi, 1971. [Google Scholar]
- Muleya, W.; Namangala, B.; Simuunza, M.; Nakao, R.; Inoue, N.; Kimura, T.; Ito, K.; Sugimoto, C.; Sawa, H. Population genetic analysis and sub-structuring of Theileria parva in the northern and eastern parts of Zambia. Parasites Vectors 2012, 5, 255. [Google Scholar] [CrossRef] [PubMed]
- DAHLD. Policy Document on Livestock in Malawi. Ministry of Agriculture, Irrigation and Water Development (MoAIWD); Department of Animal Health and Livestock Development (DAHLD): Lilongwe, Malawi, 2006. [Google Scholar]
- Musisi, F.L.; Quiroga, J.C.; Kanhai, G.K.; Kamwendo, S.P.; Mzoma, F.J.; Njuguna, L.M. Theileria parva (Kasoba): Isolation and challenge of cattle recovered from infection with other Theileria parva stocks. Rev. Elev. Med. Vet. Pays Trop. 1996, 49, 42–45. [Google Scholar] [PubMed]
- Perry, B. The control of East Coast fever of cattle by live parasite vaccination: A science-to-impact narrative. One Heal. 2016, 2, 103–114. [Google Scholar] [CrossRef]
- Salih, D.A.; Pellé, R.; Mwacharo, J.M.; Njahira, M.N.; Marcellino, W.L.; Kiara, H.; Malak, A.K.; El Hussein, A.R.M.; Bishop, R.; Skilton, R.A. Genes encoding two Theileria parva antigens recognized by CD8+ T-cells exhibit sequence diversity in South Sudanese cattle populations but the majority of alleles are similar to the Muguga component of the live vaccine cocktail. PLoS ONE 2017, 12, e0171426. [Google Scholar] [CrossRef]
- MacHugh, N.D.; Connelley, T.; Graham, S.P.; Pellé, R.; Formisano, P.; Taracha, E.L.; Ellis, S.A.; McKeever, D.J.; Burrells, A.; Morrison, W.I. CD8+ T-cell responses to Theileria parva are preferentially directed to a single dominant antigen: Implications for parasite strain-specific immunity. Eur. J. Immunol. 2009, 39, 2459–2469. [Google Scholar] [CrossRef]
- Taracha, E.L.; Goddeeris, B.M.; Morzaria, S.P.; Morrison, W.I. Parasite strain specificity of precursor cytotoxic T cells in individual animals correlates with cross-protection in cattle challenged with Theileria parva. Parasite strain specificity of precursor cytotoxic T Cells in individual animals correlates with infection and immunity. Infect. Immun. 1995, 63, 1258–1262. [Google Scholar]
- Graham, S.P.; Pellé, R.; Honda, Y.; Mwangi, D.M.; Tonukari, N.J.; Yamage, M.; Glew, E.J.; De Villiers, E.P.; Shah, T.; Bishop, R.; et al. Theileria parva candidate vaccine antigens recognized by immune bovine cytotoxic T lymphocytes. Proc. Natl. Acad. Sci. USA 2006, 103, 3286–3291. [Google Scholar] [CrossRef]
- Graham, S.P.; Honda, Y.; Pelle, R.; Mwangi, D.M.; Glew, E.J.; De Villiers, E.P.; Shah, T.; Bishop, R.; Van Der Bruggen, P.; Nene, V.; et al. A novel strategy for the identification of antigens that are recognised by bovine MHC class I restricted cytotoxic T cells in a protozoan infection using reverse vaccinology. Immunome Res. 2007, 3, 2. [Google Scholar] [CrossRef][Green Version]
- Graham, S.P.; Pellé, R.; Yamage, M.; Mwangi, D.M.; Honda, Y.; Mwakubambanya, R.S.; De Villiers, E.P.; Abuya, E.; Awino, E.; Gachanja, J.; et al. Characterization of the Fine Specificity of Bovine CD8 T-Cell Responses to Defined Antigens from the Protozoan Parasite Theileria parva. Infect. Immun. 2007, 76, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Pelle, R.; Graham, S.P.; Njahira, M.N.; Osaso, J.; Saya, R.M.; Odongo, D.; Toye, P.G.; Spooner, P.R.; Musoke, A.J.; Mwangi, D.M.; et al. Two Theileria parva CD8 T Cell Antigen Genes Are More Variable in Buffalo than Cattle Parasites, but Differ in Pattern of Sequence Diversity. PLoS ONE 2011, 6, e19015. [Google Scholar] [CrossRef] [PubMed]
- Elisa, M.; Hasan, S.D.; Moses, N.; Elpidius, R.; Skilton, R.; Gwakisa, P. Genetic and antigenic diversity of Theileria parva in cattle in Eastern and Southern zones of Tanzania. A study to support control of East Coast fever. Parasitology 2014, 142, 698–705. [Google Scholar] [CrossRef]
- Amzati, G.S.; Djikeng, A.; Odongo, D.; Nimpaye, H.; Sibeko, K.P.; Muhigwa, J.-B.B.; Madder, M.; Kirschvink, N.; Marcotty, T. Genetic and antigenic variation of the bovine tick-borne pathogen Theileria parva in the Great Lakes region of Central Africa. Parasites Vectors 2019, 12, 588–619. [Google Scholar] [CrossRef] [PubMed]
- Kerario, I.I.; Chenyambuga, S.W.; Mwega, E.D.; Rukambile, E.; Simulundu, E.; Simuunza, M. Diversity of two Theileria parva CD8+ antigens in cattle and buffalo-derived parasites in Tanzania. Ticks Tick-borne Dis. 2019, 10, 1003–1017. [Google Scholar] [CrossRef]
- Sibeko, K.P.; Geysen, D.; Oosthuizen, M.; Matthee, C.; Troskie, M.; Potgieter, F.T.; Coetzer, J.A.; Collins, N. Four p67 alleles identified in South African Theileria parva field samples. Veter- Parasitol. 2010, 167, 244–254. [Google Scholar] [CrossRef]
- Bishop, R.P.; Odongo, D.; Spooner, P.R.; Morzaria, S.P.; Oura, C.A.L.; Skilton, R.A. Multilocus genotyping of Theileria parva isolates associated with a live vaccination trial in Kenya provides evidence for transmission of immunizing parasites into local tick and cattle populations. Transbound. Emerg. Dis. 2020, 67 (Suppl. 1), 88–98. [Google Scholar] [CrossRef]
- Gwakisa, P.; Kindoro, F.; Mwega, E.; Kimera, S.; Obara, I.; Ahmed, J.; Clausen, P.; Bishop, R. Monitoring vaccinated cattle for induction and longevity of persistent tick-transmissible infection: Implications for wider deployment of live vaccination against East Coast fever in Tanzania. Transbound. Emerg. Dis. 2020, 67 (Suppl. 1), 79–87. [Google Scholar] [CrossRef]
- Bishop, R.P.; Odongo, D.; Ahmed, J.; Mwamuye, M.; Fry, L.M.; Knowles, D.P.; Nanteza, A.; Lubega, G.; Gwakisa, P.; Clausen, P.; et al. A review of recent research on Theileria parva: Implications for the infection and treatment vaccination method for control of East Coast fever. Transbound. Emerg. Dis. 2020, 67 (Suppl. 1), 56–67. [Google Scholar] [CrossRef]
- Skilton, R.A.; Bishop, R.P.; Katende, J.M.; Mwaura, S.; Morzaria, S.P. The persistence of Theileria parva infection in cattle immunized using two stocks which differ in their ability to induce a carrier state: Analysis using a novel blood spot PCR assay. Parasitology 2002, 124, 265–276. [Google Scholar] [CrossRef]
- Odongo, D.; Sunter, J.D.; Kiara, H.K.; Skilton, R.A.; Bishop, R.P. A nested PCR assay exhibits enhanced sensitivity for detection of Theileria parva infections in bovine blood samples from carrier animals. Parasitol. Res. 2009, 106, 357–365. [Google Scholar] [CrossRef]
- Chatanga, E.; Mossaad, E.; Abubaker, H.A.; Alnour, S.A.; Katakura, K.; Nakao, R.; Salim, B. Evidence of multiple point mutations in Theileria annulata cytochrome b gene incriminated in buparvaquone treatment failure. Acta Trop. 2019, 191, 128–132. [Google Scholar] [CrossRef] [PubMed]
- DNA Data Bank of Japan (DDBJ). Available online: http://www.ddbj.nig.ac.jp (accessed on 20 April 2020).
- Mtshali, K.; Khumalo, Z.T.H.; Nakao, R.; Grab, D.J.; Sugimoto, C.; Thekisoe, O. Molecular detection of zoonotic tick-borne pathogens from ticks collected from ruminants in four South African provinces. J. Veter- Med. Sci. 2015, 77, 1573–1579. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Boil. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Boil. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. genalex 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research--an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Boil. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attribute | No. of Cattle | No. of T. parva Positive (%) | p-Value |
|---|---|---|---|
| Origin | 0.000051* | ||
| Kasungu | 199 | 128 (64.3%) | |
| Nkhotakota | 185 | 78 (42.2%) | |
| Lilongwe | 62 | 37 (59.7%) | |
| Age | 0.000348* | ||
| Calves (<3 months) | 12 | 3 (25.0%) | |
| Weaners (3 months–1 year) | 83 | 32 (40.0%) | |
| Adults (>1years) | 351 | 208 (59.3%) | |
| Breed | 0.376194* | ||
| Holstein Friesian | 62 | 37 (59.7%) | |
| Malawi zebu | 384 | 206 (53.6%) | |
| Sex | 0.543121* | ||
| Male | 132 | 69 (52.3%) | |
| Female | 314 | 174 (55.4%) | |
| Vaccination status | 0. 037227* | ||
| Vaccinated | 30 | 22 (73.3) | |
| Non-vaccinated | 32 | 15 (46.9) |
| Epitope 1 (Tp227-37) (4 Variants) | Epitope 2 (Tp240-48) (3 Variants) | Epitope 3 (Tp249-59) (3 Variants) | Epitope 4 (Tp296-104) (2 Variants) | Epitope 5 (Tp298-106) (2 Variants) | Epitope 6 (Tp2138-147) (3 Variants) |
|---|---|---|---|---|---|
| SHEELKKLGML (1,60,63,64,65,66) | DGFDRDALF (1,2,60,62,63,64) | KSSHGMGKVGK (1,2,60,62,63,64) | FAQSLVCVL (1,2,60,62,63,64,65,66) | QSLVCVLMK (1,2,60,62,63,64,65,66) | KTSIPNPCKW (1,2,60,62,64,65,66) |
| SDDELDTLGML (3,61) | PDLDKNRLF (3,61) | LTSHGMGRIGR (3,61) | FAASIKCVA (3,61) | ASIKCVAQY (3,61) | KPSVPNPCDW (3,61) |
| SDEELNKLGML (2) | DGLNKDALF (65,66) | KTSKGMTEVGK (65,66) | KTSILNPCKW (63) | ||
| SREKLKKLGML (62) |
| Primer Name | Primer Sequence (5′ to 3′) | Target Gene/(PCR Type) | Product Size (bp) | Anneal. T. (°C) | Reference |
|---|---|---|---|---|---|
| IL 3231 | ATTTAAGGAACCTGACGTGACTGC | T. parva p104 gene PCR | 496 | 55 | [37] |
| IL 755 | TAAGATGCCGACTATTAATGACACC | ||||
| IL 4234 | GGCCAAGGTCTCCTTCAGAATACG | T. parva p104 gene nested PCR | 277 | 55 | [38] |
| IL 3232 | TGGGTGTGTTTCCTCGTCATCTGC | ||||
| Tp1 F outer | ATGGCCACTTCAATTGCATTTGCC | Tp1 gene PCR | 432 | 50 | [29] |
| Tp1 R outer | TAAATGAAATATTTATGAGCTTC | ||||
| Tp2 F outer | ATGAAATTGGCCGCCAGATTA | Tp2 gene PCR | 525 | 50 | [29] |
| Tp2 R outer | CTATGAAGTGCCGGAGGCTTC | ||||
| Tp1 Inner F1 | CCGCKGATCCTGGATTCT | Tp1 gene semi-nested PCR | 419 | 55 | The outer primers, [29] Inner primers, this study. |
| Tp1 R outer | TAAATGAAATATTTATGAGCTTC | ||||
| Tp1 Inner F2 | CATTTGCCGCKGATCCTG | Tp1 gene alternative semi-nested PCR | 413 | 55 | |
| Tp1 R outer | TAAATGAAATATTTATGAGCTTC | ||||
| Tp2 Inner F | CCGCCAGATTAATTAGCCTTT | Tp2 gene semi-nested PCR | 511 | 57 | |
| Tp2 R outer | CTATGAAGTGCCGGAGGCTTC | ||||
| Tp2 F outer | ATGAAATTGGCCGCCAGATTA | Tp2 gene alternative semi-nested PCR | 505 | 57 | |
| Tp2 Inner R | CCGGAGGCTTCTCCTTTTT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chatanga, E.; Hayashida, K.; Muleya, W.; Kusakisako, K.; Moustafa, M.A.M.; Salim, B.; Katakura, K.; Sugimoto, C.; Nonaka, N.; Nakao, R. Genetic Diversity and Sequence Polymorphism of Two Genes Encoding Theileria parva Antigens Recognized by CD8+ T Cells among Vaccinated and Unvaccinated Cattle in Malawi. Pathogens 2020, 9, 334. https://doi.org/10.3390/pathogens9050334
Chatanga E, Hayashida K, Muleya W, Kusakisako K, Moustafa MAM, Salim B, Katakura K, Sugimoto C, Nonaka N, Nakao R. Genetic Diversity and Sequence Polymorphism of Two Genes Encoding Theileria parva Antigens Recognized by CD8+ T Cells among Vaccinated and Unvaccinated Cattle in Malawi. Pathogens. 2020; 9(5):334. https://doi.org/10.3390/pathogens9050334
Chicago/Turabian StyleChatanga, Elisha, Kyoko Hayashida, Walter Muleya, Kodai Kusakisako, Mohamed Abdallah Mohamed Moustafa, Bashir Salim, Ken Katakura, Chihiro Sugimoto, Nariaki Nonaka, and Ryo Nakao. 2020. "Genetic Diversity and Sequence Polymorphism of Two Genes Encoding Theileria parva Antigens Recognized by CD8+ T Cells among Vaccinated and Unvaccinated Cattle in Malawi" Pathogens 9, no. 5: 334. https://doi.org/10.3390/pathogens9050334
APA StyleChatanga, E., Hayashida, K., Muleya, W., Kusakisako, K., Moustafa, M. A. M., Salim, B., Katakura, K., Sugimoto, C., Nonaka, N., & Nakao, R. (2020). Genetic Diversity and Sequence Polymorphism of Two Genes Encoding Theileria parva Antigens Recognized by CD8+ T Cells among Vaccinated and Unvaccinated Cattle in Malawi. Pathogens, 9(5), 334. https://doi.org/10.3390/pathogens9050334