Streptococcus pneumoniae Nasopharyngeal Carriage among PCV-10-Vaccinated HIV-1-Infected Children with Maintained Serological Memory in Ethiopia

 ,
,
Abstract
1. Introduction
2. Results
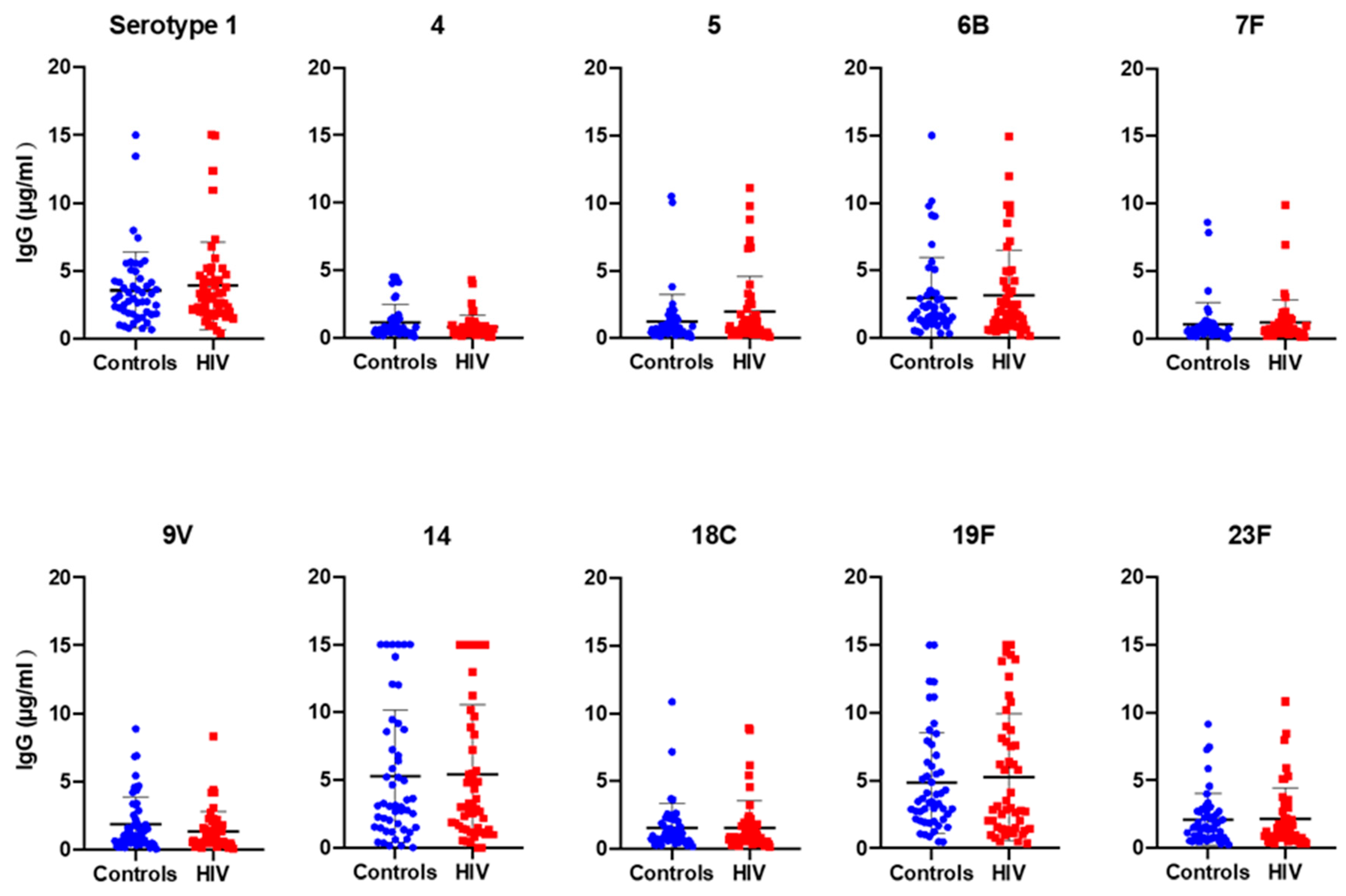
2.1. S. pneumoniae Serotype-Specific Antibody Responses
2.2. Determination of S. pneumoniae Nasopharyngeal Carriage with Whole-Genome-Sequencing-Based Serotyping and Multi-Locus Sequence Typing (MLST)
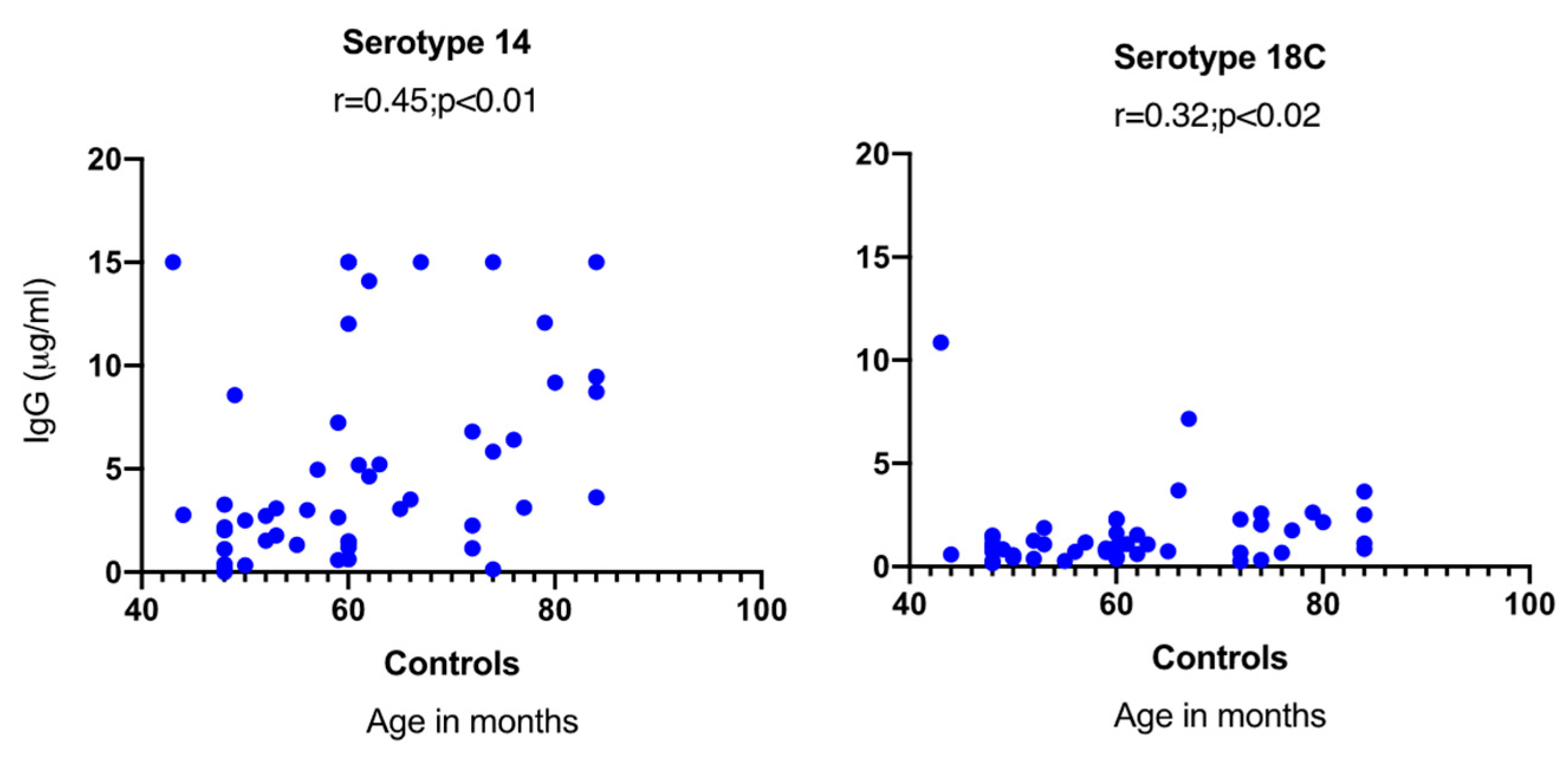
2.3. S. pneumoniae Serotype-Specific Antibody Responses in Relation to Nasopharyngeal Carriage
3. Discussion
4. Material and Methods
4.1. Study Population
4.2. Measurement of S. pneumoniae Serotype Specific IgG in Plasma
4.3. Identification of S. pneumoniae from Nasopharynx Swabs and DNA Isolation
4.4. Whole Genome Sequence-Based S. pneumoniae Serotyping
4.5. Statistical Analysis
4.6. Ethical Statement
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hackel, M.; Lascols, C.; Bouchillon, S.; Hilton, B.; Morgenstern, D.; Purdy, J. Serotype prevalence and antibiotic resistance in Streptococcus pneumoniae clinical isolates among global populations. Vaccine 2013, 31, 4881–4887. [Google Scholar] [CrossRef]
- Donkor, E.S. Understanding the pneumococcus: Transmission and evolution. Front. Cell. Infect. Microbiol. 2013, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Henriques-Normark, B.; Tuomanen, E.I. The Pneumococcus: Epidemiology, microbiology, and pathogenesis. Cold Spring Harb. Perspect. Med. 2013, 3, a010215. [Google Scholar] [CrossRef] [PubMed]
- Weiser, J.N. The pneumococcus: Why a commensal misbehaves. J. Mol. Med. 2010, 88, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Sonego, M.; Pellegrin, M.C.; Becker, G.; Lazzerini, M. Risk factors for mortality from acute lower respiratory infections (ALRI) in children under five years of age in low and middle-income countries: A systematic review and meta-analysis of observational studies. PLoS ONE 2015, 10, e0116380. [Google Scholar] [CrossRef]
- Torres, A.; Blasi, F.; Dartois, N.; Akova, M. Which individuals are at increased risk of pneumococcal disease and why? Impact of COPD, asthma, smoking, diabetes, and/or chronic heart disease on community-acquired pneumonia and invasive pneumococcal disease. Thorax 2015, 70, 984–989. [Google Scholar] [CrossRef]
- Curcio, D.; Cané, A.; Isturiz, R. Redefining risk categories for pneumococcal disease in adults: Critical analysis of the evidence. Int. J. Infect.Dis. 2015, 37, 30–35. [Google Scholar] [CrossRef]
- Feldman, C.; Anderson, R.; Rossouw, T. HIV-related pneumococcal disease prevention in adults. Expert Rev. Respir. Med. 2017, 11, 181–199. [Google Scholar] [CrossRef]
- Weiser, J.N.; Ferreira, D.M.; Paton, J.C. Streptococcus pneumoniae: Transmission, colonization and invasion. Nat. Rev. Microbiol. 2018, 16, 355–367. [Google Scholar] [CrossRef]
- Wahl, B.; O’Brien, K.L.; Greenbaum, A.; Majumder, A.; Liu, L.; Chu, Y.; Lukšić, I.; Nair, H.; McAllister, A.D.; Campbell, H.; et al. Burden of Streptococcus pneumoniae and Haemophilus influenzae type b disease in children in the era of conjugate vaccines: Global, regional, and national estimates for 2000–2015. Lancet Glob. Health 2018, 6, e744–e757. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Pneumococcal Conjugate Vaccines in Infants and Children under 5 Years of Age: WHO Position Paper Weekly Epidemiological Record; WHO: Geneva, Switzerland, 2019; Volume 94, pp. 85–104. [Google Scholar]
- McAllister, D.A.; Liu, L.; Shi, T.; Chu, Y.; Reed, C.; Burrows, J.; Adeloye, D.; Rudan, I.; Black, R.E.; Campbell, H.; et al. Global, Regional, and national estimates of pneumonia morbidity and mortality in children younger than 5 years between 2000 and 2015: A systematic analysis. Lancet Glob. Health 2019, 7, e47–e57. [Google Scholar] [CrossRef]
- Bliss, S.J.; O’Brien, K.L.; Janoff, E.N.; Cotton, M.F.; Musoke, P.; Coovadia, H.; Levine, O.S. The evidence for using conjugate vaccines to protect HIV-infected children against pneumococcal disease. Lancet Infect. Dis. 2008, 8, 67–80. [Google Scholar] [CrossRef]
- Daniels, C.C.; Rogers, P.D.; Shelton, C.M. A review of pneumococcal vaccines: Current polysaccharide vaccine recommendations and future protein antigens. J. Pediatr. Pharmacol. Ther. 2016, 21, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Westerink, M.A.; Schroeder, H.W.; Nahm, M.H. Immune Responses to pneumococcal vaccines in children and adults: Rationale for age-specific vaccination. Aging Dis. 2012, 3, 51–67. [Google Scholar]
- Goldblatt, D.; Borrow, R.; Miller, E. Natural and vaccine-induced immunity and immunologic memory to Neisseria meningitidis serogroup C in young adults. J. Infect. Dis. 2002, 185, 397–400. [Google Scholar] [CrossRef][Green Version]
- Weller, S.; Braun, M.C.; Tan, B.K.; Rosenwald, A.; Cordier, C.; Conley, M.E.; Plebani, A.; Kumararatne, D.S.; Bonnet, D.; Tournilhac, O.; et al. Human blood IgM “memory” B cells are circulating splenic marginal zone B cells harboring a prediversified immunoglobulin repertoire. Blood 2004, 104, 3647–3654. [Google Scholar] [CrossRef]
- Rodgers, G.L.; Klugman, K.P. Surveillance of the impact of pneumococcal conjugate vaccines in developing countries. Hum. Vaccin Immunother. 2016, 12, 417–420. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Correlates of Vaccine-Induced Protection: Methods and Implications; WHO/IVB/13.01; WHO: Geneva, Switzerland, 2013; pp. 1–55. [Google Scholar]
- Negash, A.A.; Asrat, D.; Abebe, W.; Hailemariam, T.; Gebre, M.; Verhaegen, J.; Aseffa, A.; Vaneechoutte, M. Pneumococcal carriage, serotype distribution, and risk factors in children with community-acquired pneumonia, 5 years after introduction of the 10-valent pneumococcal conjugate vaccine in ethiopia. Open Forum Infect. Dis. 2019, 6, ofz259. [Google Scholar] [CrossRef]
- Sime, W.T.; Aseffa, A.; Woldeamanuel, Y.; Brovall, S.; Morfeldt, E.; Henriques-Normark, B. Serotype and molecular diversity of nasopharyngeal Streptococcus pneumoniae isolates from children before and after vaccination with the ten-valent pneumococcal conjugate vaccine (PCV10) in Ethiopia. BMC Infect. Dis. 2019, 19, 409. [Google Scholar] [CrossRef]
- Mitsi, E.; Roche, A.M.; Reine, J.; Zangari, T.; Owugha, J.T.; Pennington, S.H.; Gritzfeld, J.F.; Wright, A.D.; Collins, A.M.; van Selm, S.; et al. Agglutination by anti-capsular polysaccharide antibody is associated with protection against experimental human pneumococcal carriage. Mucosal Immunol. 2017, 10, 385–394. [Google Scholar] [CrossRef]
- Malley, R.; Trzcinski, K.; Srivastava, A.; Thompson, C.M.; Anderson, P.W.; Lipsitch, M. CD4+ T cells mediate antibody-independent acquired immunity to pneumococcal colonization. Proc. Natl. Acad. Sci. USA 2005, 102, 4848–4853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Bagrade, L.; Bernatoniene, J.; Clarke, E.; Paton, J.C.; Mitchell, T.J.; Nunez, D.A.; Finn, A. Low CD4 T cell immunity to pneumolysin is associated with nasopharyngeal carriage of pneumococci in children. J. Infect. Dis. 2007, 195, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Glennie, S.J.; Banda, D.; Gould, K.; Hinds, J.; Kamngona, A.; Everett, D.D.; Williams, N.A.; Heyderman, R.S. Defective pneumococcal-specific Th1 responses in HIV-infected adults precedes a loss of control of pneumococcal colonization. Clin. Infect. Dis. 2013, 56, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Sepako, E.; Glennie, S.J.; Jambo, K.C.; Mzinza, D.; Iwajomo, O.H.; Banda, D.; van Oosterhout, J.J.; Williams, N.A.; Gordon, S.B.; Heyderman, R.S. Incomplete recovery of pneumococcal CD4 T cell immunity after initiation of antiretroviral therapy in HIV-infected malawian adults. PLoS ONE 2014, 9, e100640. [Google Scholar] [CrossRef]
- Diavatopoulos, D.A.; Short, K.R.; Price, J.T.; Wilksch, J.J.; Brown, L.E.; Briles, D.E.; Strugnell, R.A.; Wijburg, O.L. Influenza A virus facilitates Streptococcus pneumoniae and disease. FASEB J. 2010, 24, 1789–1798. [Google Scholar] [CrossRef]
- Kwambana-Adams, B.; Hanson, B.; Worwui, A.; Agbla, S.; Foster-Nyarko, E.; Ceesay, F.; Ebruke, C.; Egere, U.; Zhou, Y.; Ndukum, M.; et al. Rapid replacement by non-vaccine pneumococcal serotypes may mitigate the impact of the pneumococcal conjugate vaccine on nasopharyngeal bacterial ecology. Sci.Rep. 2017, 7, 8127. [Google Scholar] [CrossRef]
- Eisen, S.; Hayden, C.; Young, C.J.; Gilson, R.; Jungmann, E.; Jacobsen, M.C.; Poulsom, H.; Goldblatt, D.; Klein, N.J.; Baxendale, H.E. B-cell development and pneumococcal immunity in vertically acquired HIV infection. AIDS 2016, 30, 1867–1876. [Google Scholar] [CrossRef]
- Nunes, M.C.; Madhi, S.A. Safety, immunogenicity and efficacy of pneumococcal conjugate vaccine in HIV-infected individuals. Hum. Vaccin Immunother. 2012, 8, 161–173. [Google Scholar] [CrossRef]
- Jallow, S.; Madhi, S.A.; Madimabe, R.; Sipambo, N.; Violari, A.; Kala, U.; Petersen, K.; Naidoo, S.; Verwey, C.; Moore, D.P.; et al. Immunogenicity of 13-valent pneumococcal conjugate vaccine among children with underlying medical conditions. Vaccine 2017, 35, 4321–4329. [Google Scholar] [CrossRef]
- Vardanjani, H.M.; Borna, H.; Ahmadi, A. Effectiveness of pneumococcal conjugate vaccination against invasive pneumococcal disease among children with and those without HIV infection: A systematic review and meta-analysis. BMC Infect. Dis. 2019, 19, 685. [Google Scholar] [CrossRef]
- Donkor, E.S.; Annan, J.A.; Badoe, E.V.; Dayie, N.T.; Labi, A.K.; Slotved, H.C. Pneumococcal carriage among HIV infected children in Accra, Ghana. BMC Infect. Dis. 2017, 17, 133. [Google Scholar] [CrossRef] [PubMed]
- Epping, L.; van Tonder, A.J.; Gladstone, R.A.; Bentley, S.D.; Page, A.J.; Keane, J.A.; The Global Pneumococcal Sequencing Consortium. SeroBA: Rapid high-throughput serotyping of Streptococcus pneumoniae from whole genome sequence data. Microb. Genom. 2018, 4, e000186. [Google Scholar] [CrossRef]
- Voysey, M.; Fanshawe, T.R.; Kelly, D.F.; O’Brien, K.L.; Kandasamy, R.; Shrestha, S.; Thorson, S.; Hinds, J.; Pollard, A.J. Serotype-specific correlates of protection for pneumococcal carriage: An analysis of immunity in 19 countries. Clin. Infect. Dis. 2018, 66, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Madhi, S.A.; Koen, A.; Jose, L.; van Niekerk, N.; Adrian, P.V.; Cutland, C.; François, N.; Ruiz-Guiñazú, J.; Yarzabal, J.P.; Moreira, M.; et al. Vaccination with 10-valent pneumococcal conjugate vaccine in infants according to HIV status. Medicine 2017, 96, e5881. [Google Scholar] [CrossRef] [PubMed]
- Silfverdal, S.A.; Skerlikova, H.; Zanova, M.; Papúchová, D.; Traskine, M.; Borys, D.; Schuerman, L. Anamnestic immune response in 3- to 4-year-old children previously immunized with 10-valent pneumococcal nontypeable Haemophilus influenzae protein D conjugate vaccine as 2-dose or 3-dose priming and a booster dose in the first year of life. Pediatr. Infect. Dis. J. 2011, 30, e155–e163. [Google Scholar] [CrossRef]
- Prymula, R.; Habib, A.; François, N.; Borys, D.; Schuerman, L. Immunological memory and nasopharyngeal carriage in 4-year-old children previously primed and boosted with 10-valent pneumococcal non-typeable Haemophilus influenzae protein D conjugate vaccine (PHiD-CV) with or without concomitant prophylactic paracetamol. Vaccine 2013, 31, 2080–2088. [Google Scholar] [CrossRef]
- Wysocki, J.; Brzostek, J.; Konior, R.; Panzer, F.G.; François, N.A.; Ravula, S.M.; Kolhe, D.A.; Song, Y.; Dieussaert, I.; Schuerman, L.; et al. Antibody persistence and immunologic memory in children vaccinated with 4 doses of pneumococcal conjugate vaccines: Results from 2 long-term follow-up studies. Hum. Vaccin. Immunother. 2017, 13, 661–675. [Google Scholar] [CrossRef]
- Cohen, R.; Varon, E.; Béchet, S.; Bonacorsi, S.; Levy, C. Comparative impact of pneumococcal conjugate vaccines on pneumococcal meningitis according to underlying conditions. Vaccine 2016, 34, 4850–4856. [Google Scholar] [CrossRef]
- Yildirim, I.; Shea, K.M.; Little, B.A.; Silverio, A.L.; Pelton, S.I. Vaccination, underlying comorbidities, and risk of invasive pneumococcal disease. Pediatrics 2015, 135, 495–503. [Google Scholar] [CrossRef]
- Levy, C.; Varon, E.; Ouldali, N.; Béchet, S.; Bonacorsi, S.; Cohen, R. Changes in invasive pneumococcal disease spectrum after 13-valent pneumococcal conjugate vaccine implementation. Clin. Infect. Dis. 2020, 70, 446–454. [Google Scholar] [CrossRef]
- Ladhani, N.S.; Collins, S.; Djennad, A.; Sheppard, L.C.; Borrow, R.; Fry, K.N.; Andrews, J.N.; Miller, E.; Ramsay, E.M. Rapid increase in non-vaccine serotypes causing invasive pneumococcal disease in England and Wales, 2000–2017: A prospective national observational cohort study. Lancet Infect. Dis. 2018, 18, 441–451. [Google Scholar] [CrossRef]
- Vinuesa, C.G.; Cook, M.C. The molecular basis of lymphoid architecture and B cell responses: Implications for immunodeficiency and immunopathology. Curr. Mol. Med. 2001, 1, 689–725. [Google Scholar] [CrossRef] [PubMed]
- Chiodi, F. Pneumococcal vaccination of HIV-infected young adults is an important global priority. AIDS 2016, 30, 1991–1993. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, I.C.; Bazin, H.; Chassoux, D.; Gray, D.; Lortan, J. Comparative analysis of the development of B cells in marginal zones and follicles. Adv. Exp. Med. Biol. 1985, 186, 139–144. [Google Scholar] [PubMed]
- Gauvin, J.; Chagnon-Choquet, J.; Poudrier, J.; Roger, M.; Montreal primary HIV infection and slow progressor cohorts. Fluctuations in blood marginal zone b-cell frequencies may reflect migratory patterns associated with HIV-1 disease progression status. PLoS ONE 2016, 11, e0155868. [Google Scholar] [CrossRef] [PubMed]
- Amu, S.; Ruffin, N.; Rethi, B.; Chiodi, F. Impairment of B cell function during HIV-1 infection. AIDS 2013, 27, 2323–2334. [Google Scholar] [CrossRef]
- Bekele, Y.; Lemma, M.; Bobosha, K.; Yibeltal, D.; Nasi, A.; Gebre, M.; Nilsson, A.; Aseffa, A.; Howe, R.; Chiodi, F. Homing defects of B cells in HIV-1 infected children impair vaccination responses. Vaccine 2019, 37, 2348–2355. [Google Scholar] [CrossRef]
- Jochems, S.P.; Marcon, F.; Carniel, B.F.; Holloway, M.; Mitsi, E.; Smith, E.; Gritzfeld, J.F.; Solórzano, C.; Reiné, J.; Pojar, S.; et al. Inflammation induced by influenza virus impairs human innate immune control of pneumococcus. Nat. Immunol. 2018, 19, 1299–1308. [Google Scholar] [CrossRef]
- Madhi, S.A.; Kuwanda, L.; Cutland, C.; Holm, A.; Kayhty, H.; Klugman, K.P. Quantitative and qualitative antibody response to pneumococcal conjugate vaccine among African human immunodeficiency virus-infected and uninfected children. Pediatr. Infect. Dis. J. 2005, 24, 410–416. [Google Scholar] [CrossRef]
- Spoulou, V.I.; Tsoumas, D.L.; Papaevangelou, V.G.; Mostrou, G.I.; Theodoridou, M.C. Immunogenicity and immunological memory induced by a 7-valent pneumococcal CRM197 conjugate vaccine in symptomatic HIV-1 infected children. Vaccine 2005, 23, 5289–5293. [Google Scholar] [CrossRef]
- Mubarak, A.; Ahmed, M.S.; Upile, N.; Vaughan, C.; Xie, C.; Sharma, R.; Acar, P.; McCormick, M.S.; Paton, J.C.; Mitchell, T.; et al. A dynamic relationship between mucosal T helper type 17 and regulatory T-cell populations in nasopharynx evolves with age and associates with the clearance of pneumococcal carriage in humans. Clin. Microbiol. Infect. 2016, 22, 736. [Google Scholar] [CrossRef] [PubMed]
- WHO Training Manual for Enzyme Linked Immunosorbent Assay for the Quantitation of Streptococcus Pneumoniae Serotype Specific IgG (Pn PS ELISA). (007sp Version). 2000. Available online: https://www.vaccine.uab.edu/uploads/mdocs/ELISAProtocol(007sp).pdf (accessed on 19 May 2017).
- CDC. Identification and Characterization of Streptococcus Pneumoniae. 2016. Available online: https://www.cdc.gov/meningitis/lab-manual/chpt08-id-characterization-streppneumo.html (accessed on 5 July 2018).
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Mikheenko, A.; Prjibelski, A.; Saveliev, V.; Antipov, D.; Gurevich, A. Versatile genome assembly evaluation with QUAST-LG. Bioinformatics 2018, 34, i142–i150. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and samtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. J. Comput. Biol. 2013, 20, 714–737. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | PCV-10-Vaccinated | |
|---|---|---|
| Controls (n = 50) | HIV-1-Infected Children (n = 50) | |
| Median age (range) in months | 60 (43–84) | 53.5 (24–89) |
| Gender | ||
| Female | 25 (50%) | 27 (54%) |
| Male | 25 (50%) | 23 (46%) |
| Viral load * | ||
| Aviremic | NA | 26 (52%) |
| Viremic | NA | 20 (40%) |
| Median viral load | NA | 42,095 (266–1,882,592) |
| ART | ||
| ABC + 3TC + NVP | NA | 10.3% |
| AZT + 3TC + NVP | NA | 30.7% |
| AZT + 3TC + EFV | NA | 10.3% |
| AZT + 3TC + LPV/r | NA | 23.1% |
| ABC + 3TC-LPV/r | NA | 25.6% |
| Months on ART: median (range) | NA | 40 (11–83) |
| WHO stage * | ||
| Stage I | NA | 46 |
| Stage III | NA | 1 |
| Body mass index | 15.64 (12.42–19.78) | 14.49 (8.41–36.98) |
| Serotypes PCV-10 | GMC * in Controls µg/mL | GMC in HIV µg/mL | p-Value | Controls N (%) | HIV N (%) | p-Value | Total% |
|---|---|---|---|---|---|---|---|
| 1 | 2.84 | 2.9 | 0.73 | 50 (100) | 50 (100) | >0.99 | 100 |
| 4 | 0.71 | 0.56 | 0.19 | 41 (82) | 34 (68) | 0.16 | 75 |
| 5 | 0.68 | 1 | 0.09 | 40 (80) | 41 (82) | >0.99 | 81 |
| 6B | 1.9 | 1.9 | 0.99 | 49 (98) | 48 (96) | >0.99 | 97 |
| 7F | 0.63 | 0.72 | 0.44 | 38 (76) | 42 (84) | 0.45 | 80 |
| 9V | 1.04 | 0.82 | 0.28 | 44 (88) | 42 (84) | 0.77 | 86 |
| 14 | 2.78 | 2.59 | 0.92 | 47 (94) | 48 (96) | >0.99 | 95 |
| 18C | 1.03 | 0.98 | 0.76 | 45 (90) | 44 (88) | >0.99 | 89 |
| 19F | 3.59 | 3.36 | 0.70 | 50 (100) | 50 (100) | >0.99 | 100 |
| 23F | 1.52 | 1.4 | 0.67 | 48 (96) | 50 (100) | 0.49 | 98 |
| Sample ID | Age (Months) | Allele No.# | MLST Sequence Type# | Serotype¤ | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| aroE | gdh | gki | recP | spi | xpt | ddl | ||||
| C12 | 50 | 7 | 13 | 8 | 5 | 9 | 6 | 8 | 848 | 6C |
| C15 | 49 | 6 | 28 | 4 | 5 | 17 | 20 | 148 | 11162 | 15B |
| C16 | 72 | 6 | 28 | 4 | 5 | 17 | 20 | 148 | 11162 | 15B |
| C19 | 60 | 8 | 5 | 4 | 1 | 6 | 1 | 6 | 2318 | 15A |
| C21 | 84 | 7 | 13 | 8 | 6 | 15 | 2 | 8 | 4168 | 23A |
| C26 | 63 | 1 | 16 | 4 | 17 | 7 | 1 | 14 | 13704 | 20 |
| C29 | 53 | 8 | 37 | 9 | 29 | 2 | 12 | 53 | 344 | NT |
| C30 | 52 | 8 | 37 | 9 | 29 | 2 | 12 | 53 | 344 | NT |
| C33 | 52 | 1 | 2 | 29 | 4 | 43 | 277 | 255 | Novel 1 | 20 |
| C48 | 48 | 6 | 57 | 83 | 28 | 7 | 1 | 15 | 2906 | 6A |
| C49 | 60 | 162 | 13 | 4 | 10 | 17 | 115 | 28 | 5934 | 34 |
| C54 | 48 | 8 | 5 | 36 | 3 | 6 | 1 | 17 | Novel 2 | 35C |
| C67 | 74 | 7 | 13 | 4 | 5 | 7 | 88 | 9 | 373 | 35B |
| HIV02 | 53 | 8 | 5 | 36 | 3 | 15 | 1 | 6 | Novel 3 | 29 |
| HIV04 | 82 | 12 | 5 | 89 | 8 | 6 | 112 | 14 | 989 | 12F |
| HIV05 | 70 | 2 | 42 | 2 | 1 | 6 | 115 | 20 | 6465 | 10F |
| HIV08 | 48 | 2 | 42 | 9 | 1 | 6 | 19 | 20 | 6455 | 6A |
| HIV12 | 75 | 162 | 13 | 196 | 10 | 17 | 115 | 28 | Novel 4 | 34 |
| HIV18 | 40 | 10 | 20 | 14 | 1 | 6 | 1 | 29 | 218 | 7F |
| HIV20 | 61 | 2 | 16 | 8 | 6 | 25 | 6 | 28 | Novel 5 | 23A |
| HIV21 | 76 | 25 | 31 | 4 | 5 | 32 | 28 | 44 | 5063 | 18A |
| HIV22 | 95 | 1 | 32 | 227 | 1 | 15 | 26 | 11 | 5823 | 28A |
| HIV23 | 76 | 7 | 13 | 8 | 5 | 3 | 6 | 8 | Novel 6 | 23B1 |
| HIV26 | 43 | 2 | 12 | 9 | 1 | 6 | 20 | 5 | 6476 | 11B |
| HIV27 | 44 | 2 | 13 | 8 | 6 | 25 | 6 | 14 | 2345 | 19A |
| HIV30 | 40 | 7 | 11 | 10 | 1 | 6 | 8 | 1 | 156 | 11A |
| HIV31 | 89 | 162 | 42 | 4 | 10 | 17 | 115 | 28 | Novel 7 | 34 |
| HIV32 | 75 | 1 | 2 | 29 | 1 | 43 | 7 | 8 | Novel 8 | 31 |
| HIV33 | 26 | 12 | 19 | 36 | 17 | 6 | 20 | 14 | 2013 | 19A |
| HIV34 | 31 | 6 | 28 | 4 | 5 | 17 | 20 | 148 | 11162 | 15B |
| HIV35 | 50 | 7 | 11 | 10 | 1 | 6 | 8 | 1 | 156 | 11A |
| HIV38 | 84 | 7 | 13 | 8 | 5 | 6 | 6 | 124 | 988 | 23F |
| HIV40 | 24 | 10 | 34 | 4 | 10 | 495 | 405 | 75 | Novel 9 | 34 |
| HIV41 | 81 | 10 | 13 | 4 | 10 | 495 | 405 | 8 | 11546 | 34 |
| HIV42 | 49 | 54 | 5 | 4 | 1 | 6 | 1 | 111 | 991 | 15A |
| HIV55 | 52 | 58 | 5 | 4 | 1 | 218 | 79 | 5 | Novel 10 | 6C |
| HIV76 | 34 | 7 | 5 | 8 | 5 | 9 | 6 | 8 | Novel 11 | 15A |
| HIV77 | 83 | 2 | 555 | 673 | 1 | 6 | 83 | 31 | Novel 12 | 38 |
| Serotypes in PCV-10 | Antibody Threshold * (µg/mL) | Positive Controls N (%) | Positive HIV-1 Infected N (%) | p-Value | % of Total Children |
|---|---|---|---|---|---|
| 1 | 0.81 | 46 (92%) | 48 (96%) | 0.68 | 94% |
| 4 | 1.16 | 15 (30%) | 11 (22%) | 0.49 | 26% |
| 5 | 0.73 | 23 (46%) | 30 (60%) | 0.22 | 53% |
| 6B | 0.5 | 46 (92%) | 47 (94%) | >0.99 | 93% |
| 7F | 1.6 | 5 (10%) | 8 (16%) | 0.55 | 13% |
| 9V | 1.31 | 22 (44%) | 20 (40%) | 0.84 | 42% |
| 14 | 2.48 | 32 (64%) | 30 (60%) | 0.84 | 62% |
| 18C | 1.32 | 18 (36%) | 18 (36%) | >0.99 | 36% |
| 19F | 2.54 | 35 (70%) | 30 (60%) | 0.40 | 65% |
| 23F | 0.63 | 42 (84%) | 41 (82%) | >0.99 | 83% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemma, M.; Bekele, Y.; Petkov, S.; Hägglund, M.; Petros, B.; Aseffa, A.; Howe, R.; Chiodi, F. Streptococcus pneumoniae Nasopharyngeal Carriage among PCV-10-Vaccinated HIV-1-Infected Children with Maintained Serological Memory in Ethiopia. Pathogens 2020, 9, 159. https://doi.org/10.3390/pathogens9030159
Lemma M, Bekele Y, Petkov S, Hägglund M, Petros B, Aseffa A, Howe R, Chiodi F. Streptococcus pneumoniae Nasopharyngeal Carriage among PCV-10-Vaccinated HIV-1-Infected Children with Maintained Serological Memory in Ethiopia. Pathogens. 2020; 9(3):159. https://doi.org/10.3390/pathogens9030159
Chicago/Turabian StyleLemma, Mahlet, Yonas Bekele, Stefan Petkov, Moa Hägglund, Beyene Petros, Abraham Aseffa, Rawleigh Howe, and Francesca Chiodi. 2020. "Streptococcus pneumoniae Nasopharyngeal Carriage among PCV-10-Vaccinated HIV-1-Infected Children with Maintained Serological Memory in Ethiopia" Pathogens 9, no. 3: 159. https://doi.org/10.3390/pathogens9030159
APA StyleLemma, M., Bekele, Y., Petkov, S., Hägglund, M., Petros, B., Aseffa, A., Howe, R., & Chiodi, F. (2020). Streptococcus pneumoniae Nasopharyngeal Carriage among PCV-10-Vaccinated HIV-1-Infected Children with Maintained Serological Memory in Ethiopia. Pathogens, 9(3), 159. https://doi.org/10.3390/pathogens9030159