Colony Stimulating Factors in Early Feline Infectious Peritonitis Virus Infection of Monocytes and in End Stage Feline Infectious Peritonitis; A Combined In Vivo And In Vitro Approach

 ,
, 
 ,
,
Abstract
1. Introduction
2. Results
2.1. Cytokines Including CSFs Are Constitutively Transcribed in Feline Monocytes with High Variability, and Overall FIPV Infection Has a Selective Effect on Their Transcription Levels
2.2. FIPV Infection of Feline Monocytes Has a Limited Effect on the Transcription of Selected Cytokines during the First 9 h Post Infection
2.3. Cytokine Transcription Levels in Feline Monocytes after FIPV Infection Show Limited Age-Related Variation Which Also Affects the Response to Infection
2.4. Hemolymphatic Organs of Cats with FIP
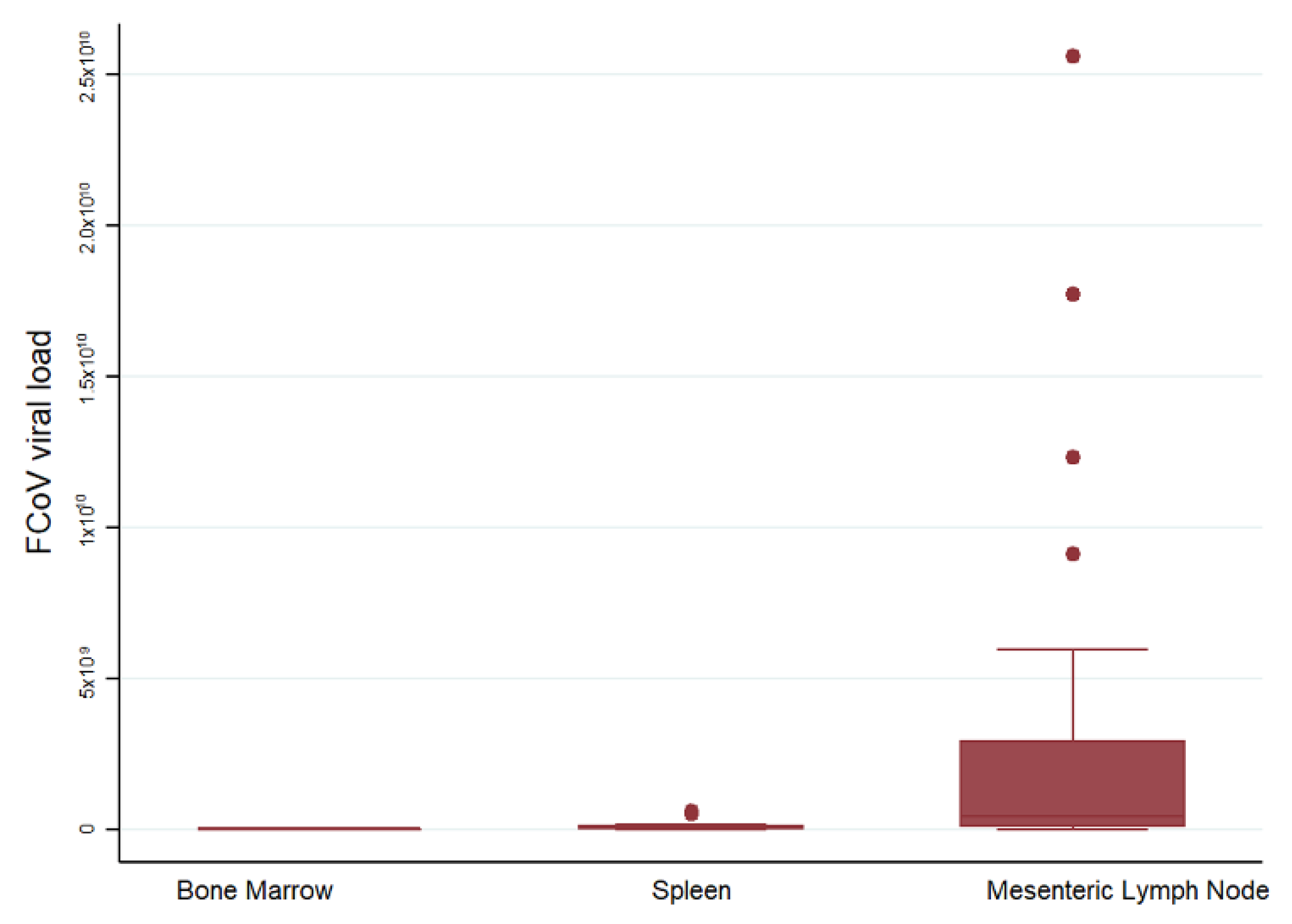
2.4.1. FCoV Levels in Bone Marrow, Spleen, and Mesenteric Lymph Node of Cats with FIP
2.4.2. Changes in G-CSF Transcription Levels in the Bone Marrow, Spleen, and Mesenteric Lymph Node in Cats with FIP
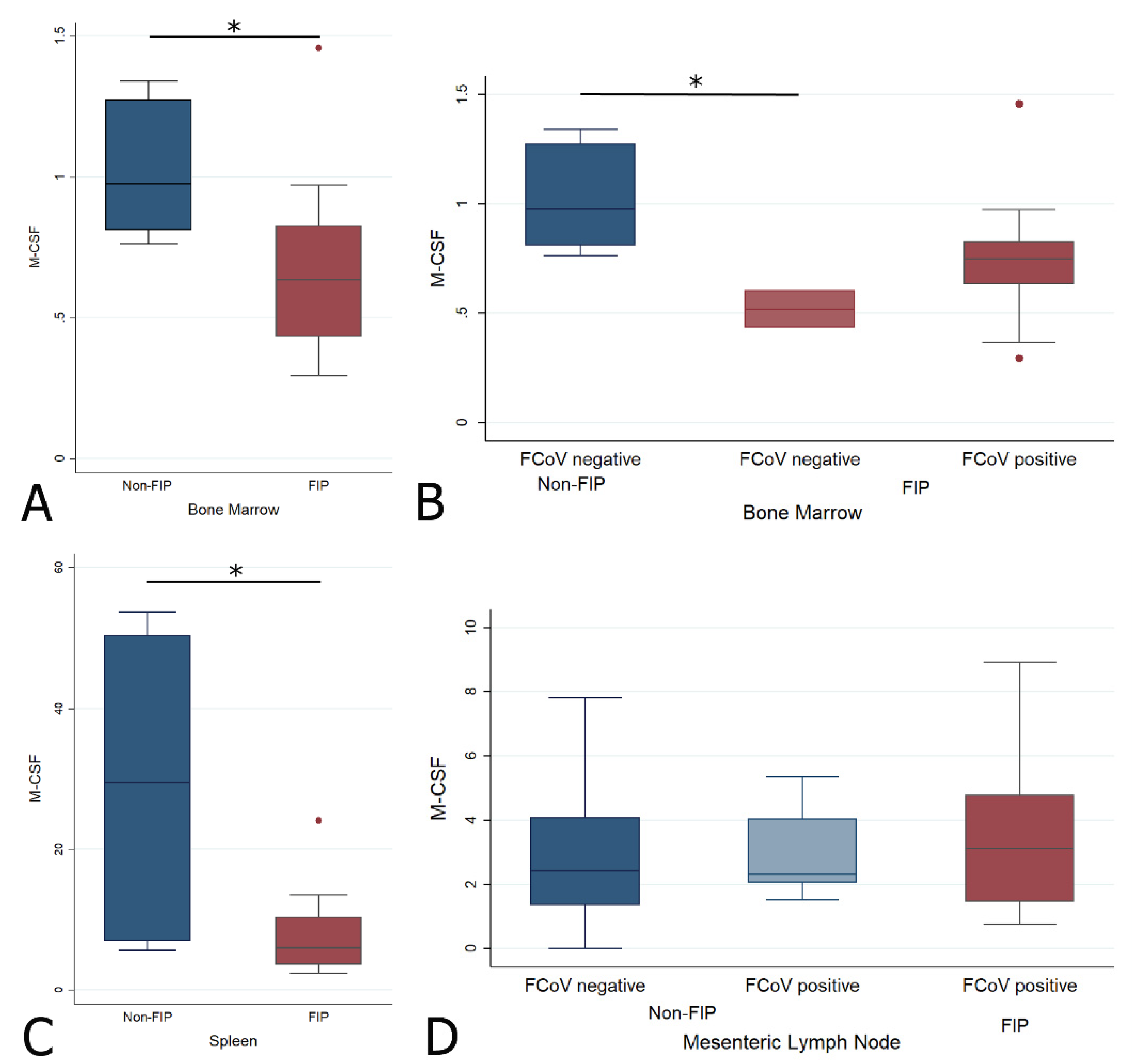
2.4.3. Changes in M-CSF Transcription Levels in the Bone Marrow, Spleen, and Mesenteric Lymph Node in Cats with FIP
2.4.4. Changes in GM-CSF Transcription Levels in the Bone Marrow, Spleen, and Mesenteric Lymph Node in Cats with FIP
2.4.5. Correlations between FCoV Load and CSFs
3. Discussion
4. Materials and Methods
4.1. Cases
4.1.1. Blood Donors
4.1.2. Clinical Cases
4.2. In Vitro Infection Studies
4.3. Quantitative Reverse Transcriptase PCR
4.3.1. RNA Extraction
4.3.2. RT-qPCR for FCoV RNA
4.3.3. DNAse Treatment, Reverse Transcription and TaqMan qPCR for Feline G-CSF, M-CSF and GM-CSF
4.4. Quantification of Cytokine Transcripts
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kipar, A.; May, H.; Menger, S.; Weber, M.; Leukert, W.; Reinacher, M. Morphologic features and development of granulomatous vasculitis in feline infectious peritonitis. Vet. Pathol. 2005, 42, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C. An update on feline infectious peritonitis: Virology and immunopathogenesis. Vet. J. 2014, 201, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Kipar, A.; Meli, M.L. Feline infectious peritonitis: Still an enigma? Vet. Pathol. 2014, 51, 505–526. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Watabe, Y.; Nakayama, H.; Fujiwara, K. Enteritis due to feline infectious peritonitis virus. Jpn. J. Vet. Sci. 1982, 44, 97–106. [Google Scholar] [CrossRef]
- Gunn-Moore, D.A.; Caney, S.M.; Gruffydd-Jones, T.J.; Helps, C.R.; Harbour, D.A. Antibody and cytokine responses in kittens during the development of feline infectious peritonitis (FIP). Vet. Immunol. Immunopathol. 1998, 65, 221–242. [Google Scholar] [CrossRef]
- Kipar, A.; Baptiste, K.; Barth, A.; Reinacher, M. Natural FCoV infection: Cats with FIP exhibit significantly higher viral loads than healthy infected cats. J. Feline Med. Surg. 2006, 8, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Meli, M.L.; Kipar, A.; Müller, C.; Jenal, K.; Gönczi, E.; Borel, N.; Gunn-Moore, D.; Chalmers, S.; Lin, F.; Reinacher, M.; et al. High viral loads despite absence of clinical and pathological findings in cats experimentally infected with feline coronavirus (FCoV) type I and in naturally FCoV-infected cats. J. Feline Med. Surg. 2004, 6, 69–81. [Google Scholar] [CrossRef]
- Can-Şahna, K.; Soydal Ataseven, V.; Pinar, D.; Çiǧdem Oǧuzoǧlu, T. The detection of feline coronaviruses in blood samples from cats by mRNA RT-PCR. J. Feline Med. Surg. 2007, 9, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Kipar, A.; Meli, M.L.; Baptiste, K.E.; Bowker, L.J.; Lutz, H. Sites of feline coronavirus persistence in healthy cats. J. Gen. Virol. 2010, 91, 1698–1707. [Google Scholar] [CrossRef] [PubMed]
- Kipar, A.; Köhler, K.; Leukert, W.; Reinacher, M. A comparison of lymphatic tissues from cats with spontaneous feline infectious peritonitis (FIP), cats with FIP virus infection but no FIP, and cats with no infection. J. Comp. Pathol. 2001, 125, 182–191. [Google Scholar] [CrossRef]
- Kipar, A.; Meli, M.L.; Failing, K.; Euler, T.; Gomes-Keller, M.A.; Schwartz, D.; Lutz, H.; Reinacher, M. Natural feline coronavirus infection: Differences in cytokine patterns in association with the outcome of infection. Vet. Immunol. Immunopathol. 2006, 112, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Kipar, A.; Leutenegger, C.M.; Hetzel, U.; Akens, M.K.; Mislin, C.N.; Reinacher, M.; Lutz, H. Cytokine mRNA levels in isolated feline monocytes. Vet. Immunol. Immunopathol. 2001, 78, 305–315. [Google Scholar] [CrossRef]
- Takano, T.; Ohyama, T.; Kokumoto, A.; Satoh, R.; Hohdatsu, T. Vascular endothelial growth factor (VEGF), produced by feline infectious peritonitis (FIP) virus-infected monocytes and macrophages, induces vascular permeability and effusion in cats with FIP. Virus Res. 2011, 158, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Ushach, I.; Zlotnik, A. Biological role of granulocyte macrophage colony-stimulating factor (GM-CSF) and macrophage colony-stimulating factor (M-CSF) on cells of the myeloid lineage. J. Leukoc. Biol. 2016, 100, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A. Colony-stimulating factors in inflammation and autoimmunity. Nat. Rev. Immunol. 2008, 8, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A.; Cook, A.D.; Tak, P.P. Anti-colony-stimulating factor therapies for inflammatory and autoimmune diseases. Nat. Rev. Drug Discov. 2017, 16, 53–70. [Google Scholar] [CrossRef]
- Katakura, F.; Nishiya, K.; Wentzel, A.S.; Hino, E.; Miyamae, J.; Okano, M.; Wiegertjes, G.F.; Moritomo, T. Paralogs of common carp granulocyte colony-stigmulating factor (G-CSF) have different functions regarding development traffickingand activation of neutrophils. Front. Immunol. 2019, 10, 255. [Google Scholar] [CrossRef]
- Takano, T.; Azuma, N.; Satoh, M.; Toda, A.; Hashida, Y.; Satoh, R.; Hohdatsu, T. Neutrophil survival factors (TNF-alpha, GM-CSF, and G-CSF) produced by macrophages in cats infected with feline infectious peritonitis virus contribute to the pathogenesis of granulomatous lesions. Arch. Virol. 2009, 154, 775–781. [Google Scholar] [CrossRef]
- Takano, T.; Hohdatsu, T.; Toda, A.; Tanabe, M.; Koyama, H. TNF-alpha, produced by feline infectious peritonitis virus (FIPV)-infected macrophages, upregulates expression of type II FIPV receptor feline aminopeptidase N in feline macrophages. Virology 2007, 364, 64–72. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Smith, K. Mesenteric lymph nodes at the center of immune anatomy. J. Exp. Med. 2006, 203, 497–500. [Google Scholar] [CrossRef]
- Christianson, K.K.; Ingersoll, J.D.; Landon, R.M.; Pfeiffer, N.E.; Gerber, J.D. Characterization of a temperature sensitive feline infectious peritonitis coronavirus. Arch. Virol. 1989, 109, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Gut, M.; Leutenegger, C.M.; Huder, J.B.; Pedersen, N.C.; Lutz, H. One-tube fluorogenic reverse transcription-polymerase chain reaction for the quantitation of feline coronaviruses. J. Virol. Methods 1999, 77, 37–46. [Google Scholar] [CrossRef]
- Regan, A.D.; Cohen, R.D.; Whittaker, G.R. Activation of p38 MAPK by feline infectious peritonitis virus regulates pro-inflammatory cytokine production in primary blood-derived feline mononuclear cells. Virology 2009, 384, 135–143. [Google Scholar] [CrossRef]
- Takano, T.; Katada, Y.; Moritoh, S.; Ogasawara, M.; Satoh, K.; Satoh, R.; Tanabe, M.; Hohdatsu, T. Analysis of the mechanism of antibody-dependent enhancement of feline infectious peritonitis virus infection: Aminopeptidase N is not important and a process of acidification of the endosome is necessary. J. Gen. Virol. 2008, 89, 1025–1029. [Google Scholar] [CrossRef]
- Fish, E.J.; Diniz, P.P.V.P.; Juan, Y.C.; Bossong, F.; Collisson, E.W.; Drechsler, Y.; Kaltenboeck, B. Cross-sectional quantitative RT-PCR study of feline coronavirus viremia and replication in peripheral blood of healthy shelter cats in Southern California. J. Feline Med. Surg. 2018, 20, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Dewerchin, H.L.; Cornelissen, E.; Nauwynck, H.J. Replication of feline coronaviruses in peripheral blood monocytes. Arch. Virol. 2005, 150, 2483–2500. [Google Scholar] [CrossRef]
- Kipar, A.; Baptiste, K.; Meli, M.L.; Barth, A.; Knietsch, M.; Reinacher, M.; Lutz, H. Age-related dynamics of constitutive cytokine transcription levels of feline monocytes. Exp. Gerontol. 2005, 40, 243–248. [Google Scholar] [CrossRef]
- Malbon, A.J.; Meli, M.L.; Barker, E.N.; Davidson, A.D.; Tasker, S.; Kipar, A. Inflammatory mediators in the mesenteric lymph nodes, site of a possible intermediate phase in the immune response to feline coronavirus and the pathogenesis of feline infectious peritonitis? J. Comp. Pathol. 2019, 166, 69–86. [Google Scholar] [CrossRef]
- Bálint, Á.; Farsang, A.; Szeredi, L.; Zádori, Z.; Belák, S. Recombinant feline coronaviruses as vaccine candidates confer protection in SPF but not in conventional cats. Vet. Microbiol. 2014, 169, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Van Hamme, E.; Desmarets, L.; Dewerchin, H.L.; Nauwynck, H.J. Intriguing interplay between feline infectious peritonitis virus and its receptors during entry in primary feline monocytes. Virus Res. 2011, 160, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.L.; Castle, B.E.; Kehry, M.R. CD40-mediated signaling in monocytic cells: Up-regulation of tumor necrosis factor receptor-associated factor mRNAs and activation of mitogen-activated protein kinase signaling pathways. Int. Immunol. 2001, 13, 273–283. [Google Scholar] [CrossRef]
- Bernad, A.; Kopf, M.; Kulbacki, R.; Weich, N.; Koehler, G.; Gutierrez-Ramos, J.C. Interleukin-6 is required in vivo for the regulation of stem cells and committed progenitors of the hematopoietic system. Immunity 1994, 1, 725–731. [Google Scholar] [CrossRef]
- Shang, X.Z.; Issekutz, A.C. Enhancement of Monocyte Transendothelial Migration by Granulocyte-Macrophage Colony-Stimulating Factor: Requirement for Chemoattractant and CD11a/CD18 Mechanisms. Eur. J. Immunol. 1999, 29, 3571–3582. [Google Scholar] [CrossRef]
- Fossati, G.; Mazzucchelli, I.; Gritti, D.; Ricevuti, G.; Edwards, S.W.; Moulding, D.A.; Rossi, M.L. In vitro effects of GM-CSF on mature peripheral blood neutrophils. Int. J. Mol. Med. 1998, 1, 943–951. [Google Scholar] [CrossRef]
- Lotfi, N.; Zhang, G.X.; Esmaeil, N.; Rostami, A. Evaluation of the effect of GM-CSF blocking on the phenotype and function of human monocytes. Sci. Rep. 2020, 10, 1567. [Google Scholar] [CrossRef]
- Castell, J.V.; Geiger, T.; Gross, V.; Andus, T.; Walter, E.; Hirano, T.; Kishimoto, T.; Heinrich, P.C. Plasma clearance, organ distribution and target cells of interleukin-6/hepatocyte-stimulating factor in the rat. Eur. J. Biochem. 1988, 177, 357–361. [Google Scholar] [CrossRef]
- Cebon, J.; Dempsey, P.; Fox, R.; Kannourakis, G.; Bonnem, E.; Burgess, A.; Morstyn, G. Pharmacokinetics of human granulocyte-macrophage colony-stimulating factor using a sensitive immunoassay. Blood 1988, 72, 1340–1347. [Google Scholar] [CrossRef]
- Wanidworanun, C.; Strober, W. Predominant role of tumor necrosis factor-alpha in human monocyte IL-10 synthesis. J. Immunol. 1993, 151, 6853–6861. [Google Scholar]
- Gelain, M.E.; Meli, M.L.; Paltrinieri, S. Whole blood cytokine profiles in cats infected by feline coronavirus and healthy non-FCoV infected specific pathogen-free cats. J. Feline Med. Surg. 2006, 8, 389–399. [Google Scholar] [CrossRef]
- Porter, E.; Tasker, S.; Day, M.J.; Harley, R.; Kipar, A.; Siddell, S.G.; Helps, C.R. Amino acid changes in the spike protein of feline coronavirus correlate with systemic spread of virus from the intestine and not with feline infectious peritonitis. Vet. Res. 2014, 45, 49. [Google Scholar] [CrossRef] [PubMed]
- Barker, E.N.; Stranieri, A.; Helps, C.R.; Porter, E.L.; Davidson, A.D.; Day, M.J.; Knowles, T.; Kipar, A.; Tasker, S. Limitations of using feline coronavirus spike protein gene mutations to diagnose feline infectious peritonitis. Vet. Res. 2017, 48, 60. [Google Scholar] [CrossRef]
- Kipar, A.; Bellmann, S.; Kremendahl, J.; Köhler, K.; Reinacher, M. Cellular composition, coronavirus antigen expression and production of specific antibodies in lesions in feline infectious peritonitis. Vet. Immunol. Immunopathol. 1998, 65, 243–257. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pitrak, D.L. Effects of granulocyte colony-stimulating factor and granulocyte-macrophage colony-stimulating factor on the bactericidal functions of neutrophils. Curr. Opin. Hematol. 1997, 4, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Kipar, A.; Bellmann, S.; Gunn-Moore, D.A.; Leukert, W.; Köhler, K.; Menger, S.; Reinacher, M. Histopathological alterations of lymphatic tissues in cats without feline infectious peritonitis after long-term exposure to FIP virus. Vet. Microbiol. 1999, 69, 131–137. [Google Scholar] [CrossRef]
- Takano, T.; Tomiyama, Y.; Katoh, Y.; Nakamura, M.; Satoh, R.; Hohdatsu, T. Mutation of neutralizing/antibody-dependent enhancing epitope on spike protein and 7b gene of feline infectious peritonitis virus: Influences of viral replication in monocytes/macrophages and virulence in cats. Virus Res. 2011, 156, 72–80. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | Organ | Comparison | Probability |
| BM | FIP vs. non-FIP | <0.05 * | |
| FIP FCoV +ve vs. FIP FCoV −ve | 0.099 | ||
| FIP FCoV +ve vs. non-FIP | 0.059 | ||
| FIP FCoV −ve vs. non-FIP | <0.05 * | ||
| Spleen | FIP vs. non-FIP | 0.564 | |
| MLN | FIP vs. non-FIP | <0.01 * | |
| FIP vs. non-FIP FCoV +ve | <0.01 * | ||
| FIP vs. non-FIP FCoV −ve | <0.01 * | ||
| non-FIP FCoV +ve vs. non FIP FCoV −ve | 0.574 | ||
| B | Organ | FIP Median (n) | Non-FIP Median (n) |
| BM | 25,158.38 (11) | 0.872 (6) | |
| Spleen | 1.03 × 107 (14) | 6.298 (6) | |
| MLN | 4.25 × 108 (30) | all cases 3.082 (40) | |
| FCoV +ve 2.249 (10) | |||
| FCoV −ve 3.120 (30) |
| A | Organ | Comparison | Probability |
| BM | FIP vs. non-FIP | <0.05 * | |
| FIP FCoV +ve vs. FIP FCoV −ve | 0.239 | ||
| FIP FCoV +ve vs. non-FIP | 0.077 | ||
| FIP FCoV −ve vs. non-FIP | <0.05 * | ||
| Spleen | FIP vs. non-FIP | <0.05 * | |
| MLN | FIP vs. non-FIP | 0.476 | |
| FIP vs. non-FIP FCoV +ve | 0.975 | ||
| FIP vs. non-FIP FCoV −ve | 0.383 | ||
| non-FIP FCoV +ve vs. non FIP FCoV −ve | 0.731 | ||
| B | Organ | FIP Median (n) | non-FIP Median (n) |
| BM | 0.635 (11) | 0.975 (6) | |
| Spleen | 5.992 (14) | 29.449 (6) | |
| MLN | 3.133 (30) | all cases 2.422 (40) | |
| FCoV +ve 2.249 (10) | |||
| FCoV −ve 3.120 (30) |
| A | Organ | Comparison | Probability |
| BM | FIP vs. non-FIP | 0.191 | |
| FIP FCoV +ve vs. FIP FcoV −ve | 0.814 | ||
| FIP FcoV +ve vs. non-FIP | 0.157 | ||
| FIP FcoV −ve vs. non-FIP | 0.739 | ||
| Spleen | FIP vs. non-FIP | <0.05 * | |
| MLN | FIP vs. non-FIP | 0.794 | |
| FIP vs. non-FIP FcoV +ve | 0.349 | ||
| FIP vs. non-FIP FcoV −ve | 0.442 | ||
| non-FIP FcoV +ve vs. non FIP FcoV −ve | 0.075 | ||
| B | Organ | FIP Median (n) | Non-FIP Median (n) |
| BM | 3.462 (11) | 0.965 (6) | |
| Spleen | 2.400 (14) | 17.787 (6) | |
| MLN | 46.143 (30) | all cases 41.967 (40) | |
| FcoV +ve 71.609 (10) | |||
| FcoV −ve 36.472 (30) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malbon, A.J.; Michalopoulou, E.; Meli, M.L.; Barker, E.N.; Tasker, S.; Baptiste, K.; Kipar, A. Colony Stimulating Factors in Early Feline Infectious Peritonitis Virus Infection of Monocytes and in End Stage Feline Infectious Peritonitis; A Combined In Vivo And In Vitro Approach. Pathogens 2020, 9, 893. https://doi.org/10.3390/pathogens9110893
Malbon AJ, Michalopoulou E, Meli ML, Barker EN, Tasker S, Baptiste K, Kipar A. Colony Stimulating Factors in Early Feline Infectious Peritonitis Virus Infection of Monocytes and in End Stage Feline Infectious Peritonitis; A Combined In Vivo And In Vitro Approach. Pathogens. 2020; 9(11):893. https://doi.org/10.3390/pathogens9110893
Chicago/Turabian StyleMalbon, Alexandra J., Eleni Michalopoulou, Marina L. Meli, Emi N. Barker, Séverine Tasker, Keith Baptiste, and Anja Kipar. 2020. "Colony Stimulating Factors in Early Feline Infectious Peritonitis Virus Infection of Monocytes and in End Stage Feline Infectious Peritonitis; A Combined In Vivo And In Vitro Approach" Pathogens 9, no. 11: 893. https://doi.org/10.3390/pathogens9110893
APA StyleMalbon, A. J., Michalopoulou, E., Meli, M. L., Barker, E. N., Tasker, S., Baptiste, K., & Kipar, A. (2020). Colony Stimulating Factors in Early Feline Infectious Peritonitis Virus Infection of Monocytes and in End Stage Feline Infectious Peritonitis; A Combined In Vivo And In Vitro Approach. Pathogens, 9(11), 893. https://doi.org/10.3390/pathogens9110893