Biochemical Characterization of Clinical Strains of Staphylococcus spp. and Their Sensitivity to Polyphenols-Rich Extracts from Pistachio (Pistacia vera L.)


 ,
,  and
and 
Abstract

1. Introduction
2. Results
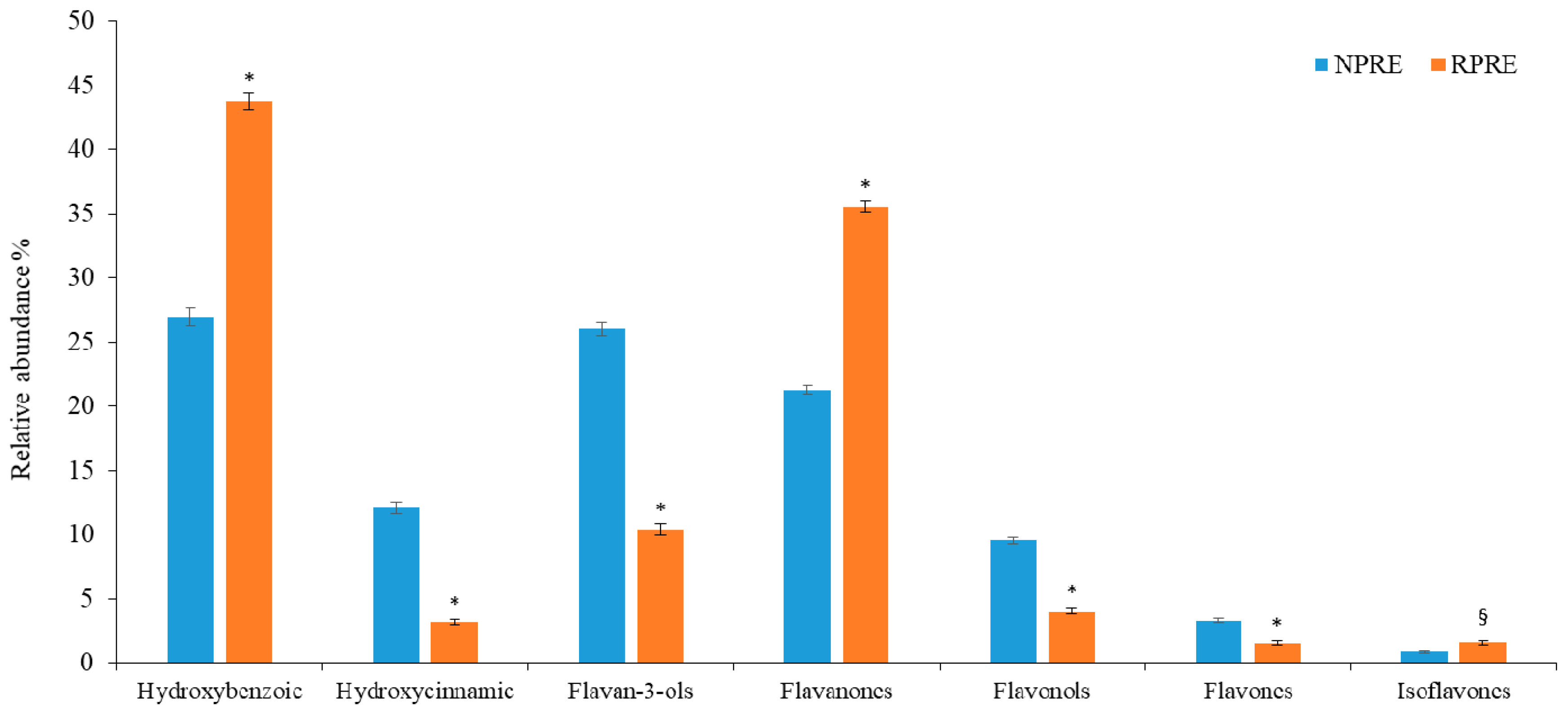
2.1. Polyphenols and Radical Scavenging Activity
2.2. Phenotypic Identification of Staphylococcus Strains
2.3. Antimicrobial Activity of Pistachio Extracts
3. Discussion
4. Materials and Methods
4.1. Pistachio Extracts
4.2. Total Phenols
4.3. Radical Scavenging Activity
4.4. Polyphenolic Profile
4.5. Microbial Strains, Culture Conditions, and Phenotypic Characterization
4.6. Lipase Activity
4.7. Susceptibility Studies
4.8. Effect on Biofilm Formation
4.9. Statistical Analysis
Author Contribution
Funding
Conflict of Interest
References
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Maki, D.G.; Kluger, D.M.; Cmich, C.J. The risk of bloodstream infections in adults with different intravascular devices: A systematic review of 200 published prospective studies. Mayo Clin. Proc. 2006, 81, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Thornton, R.B.; Wiertsema, S.P.; Kirkham, L.A.S.; Rigby, P.J.; Vijayasekaran, S.; Coates, H.L.; Richmond, P.C. Neuthophil extracellular traps and bacterial biofilms in middle ear effusion of children with recurrent acute otitis media: A potential treatment target. PLoS ONE 2013, 8, e53837. [Google Scholar] [CrossRef] [PubMed]
- Beikler, T.; Flemmig, T.F. Oral biofilm-associated diseases: Trends and implications for quality of life, systemic health and expenditures. Periodontology 2000 2011, 55, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Vieira Colombo, A.P.; Magalhães, C.B.; Hartenbach, F.A.; Martins do Souto, R.; Maciel da Silva-Boghossian, C. Periodontal-disease-associated biofilm: A reservoir for pathogens of medical importance. Microb. Pathog. 2016, 94, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Bisignano, C.; D’Arrigo, M.; Ginestra, G.; Arena, A.; Tomaino, A.; Wickham, M.S. Antimicrobial potential of polyphenols extracted from almond skins. Lett. Appl. Microbiol. 2010, 51, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Bisignano, C.; Filocamo, A.; La Camera, E.; Zummo, S.; Fera, M.T.; Mandalari, G. Antibacterial activities of almond skins on cagA-positive and-negative clinical isolates of Helicobacter pylori. BMC Microbiol. 2013, 13, 103. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Bennett, R.N.; Bisignano, G.; Trombetta, D.; Saija, A.; Faulds, C.B.; Gasson, M.J.; Narbad, A. Antimicrobial activity of flavonoids extracted from bergamot (Citrus bergamia Risso) peel, a byproduct of the essential oil industry. J. Appl. Microbiol. 2007, 103, 2056–2064. [Google Scholar] [CrossRef] [PubMed]
- Filocamo, A.; Bisignano, C.; Mandalari, G.; Navarra, M. In Vitro Antimicrobial Activity and Effect on Biofilm Production of a White Grape Juice (Vitis vinifera) Extract. Evid. Based Complement. Alternat. Med. 2015, 856243. [Google Scholar] [CrossRef]
- Bisignano, C.; Filocamo, A.; Ginestra, G.; Giofre’, S.V.; Navarra, M.; Romeo, R.; Mandalari, G. 3,4-DHPEA-EA from Olea Europaea L. is effective against standard and clinical isolates of Staphylococcus sp. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Furneri, P.M.; Mondello, L.; Mandalari, G.; Paolino, D.; Dugo, P.; Garozzo, A.; Bisignano, G. In vitro antimycoplasmal activity of Citrus bergamia essential oil and its major components. Eur. J. Med. Chem. 2012, 52, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Filocamo, A.; Bisignano, C.; Ferlazzo, N.; Cirmi, S.; Mandalari, G.; Navarra, M. In vitro effect of bergamot (Citrus bergamia) juice against cagA-positive and-negative clinical isolates of Helicobacter pylori. BMC Complement. Altern. Med. 2015, 15, 256. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture. Available online: https://www.ams.usda.gov (accessed on 9 July 2018).
- Mandalari, G.; Bisignano, C.; Filocamo, A.; Chessa, S.; Sarò, M.; Torre, G.; Faulks, R.M.; Dugo, P. Bioaccessibility of pistachio polyphenols, xanthophylls, and tocopherols during simulated human digestion. Nutrition 2013, 29, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Bisignano, C.; Filocamo, A.; Faulks, R.M.; Mandalari, G. In vitro antimicrobial activity of pistachio (Pistacia vera L.) polyphenols. FEMS Microbiol. Lett. 2013, 341, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Hatamnia, A.A.; Abbaspour, N.; Darvishzadeh, R. Antioxidant activity and phenolic profile of different parts of Bene (Pistacia atlantica subsp. kurdica) fruits. Food Chem. 2014, 145, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Tomaino, A.; Martorana, M.; Arcoraci, T.; Monteleone, D.; Giovinazzo, C.; Saija, A. Antioxidant activity and phenolic profile of pistachio (Pistacia vera L., variety Bronte) seeds and skins. Biochimie 2010, 92, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Gomber, C. Superoxide dismutase, protease and lipase expression in clinical isolates of Staphylococcus aureus: A tool for antimicrobial drug discovery. Mol. Cell. Biochem. 2010, 341, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Ippolito, G.; Leone, S.; Lauria, F.N.; Nicastri, E.; Wenzel, R.P. Methicillin-resistant Staphylococcus aureus: The superbug. Int. J. Infect. Dis. 2010, 14, S7–S11. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Choi, K.H.; Min, J.; Kim, H.J.; Jee, J.P.; Park, B.J. Functionalized ZnO Nanoparticles with Gallic Acid for Antioxidant and Antibacterial Activity against Methicillin-Resistant S. aureus. Nanomaterials (Basel) 2017, 7, 365. [Google Scholar] [CrossRef] [PubMed]
- Salaheen, S.; Peng, M.; Joo, J.; Teramoto, H.; Biswas, D. Eradication and Sensitization of Methicillin Resistant Staphylococcus aureus to Methicillin with Bioactive Extracts of Berry Pomace. Front. Microbiol. 2017, 8, 253. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Ouyang, P.; Yuan, Z.; Yin, Z.; Fu, H.; Lin, J.; He, C.; Shu, G.; Yuan, Z.; Xu, S.; et al. Eriodyctiol protects against Staphylococcus aureus-induced lung cell injury by inhibiting alpha-hemolysin expression. World J. Microbiol. Biotechnol. 2018, 34, 64. [Google Scholar]
- Monforte, M.T.; Smeriglio, A.; Germanò, M.P.; Pergolizzi, S.; Circosta, C.; Galati, E.M. Evaluation of antioxidant, antiinflammatory, and gastroprotective properties of Rubus fruticosus L. fruit juice. Phytother. Res. 2018, 32, 1404–1414. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Denaro, M.; Barreca, D.; D’Angelo, V.; Germanò, M.P.; Trombetta, D. Polyphenolic profile and biological activities of black carrot crude extract (Daucus carota L. ssp. sativus var. atrorubens Alef.). Fitoterapia 2018, 124, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Paterniti, I.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Bisignano, C.; Gugliandolo, E.; Carughi, A.; Esposito, E.; Mandalari, G.; Cuzzocrea, S. The Anti-Inflammatory and Antioxidant Potential of Pistachios (Pistacia vera L.) In Vitro and In Vivo. Nutrients 2017, 9, 915. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Fusco, R.; Gugliandolo, E.; D’Amico, R.; Campolo, M.; Latteri, S.; Carughi, A.; Mandalari, G.; Cuzzocrea, S. The Antioxidant Activity of Pistachios Reduces Cardiac Tissue Injury of Acute Ischemia/Reperfusion (I/R) in Diabetic Streptozotocin (STZ)-Induced Hyperglycaemic Rats. Front. Pharmacol. 2018, 9, 51. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, P.D.; Shah, S.; Anderson, J.C.; Hara, Y.; Hamilton-Miller, J.M.; Taylor, P.W. Modulation of beta-lactam resistance in Staphylococcus aureus by catechins and gallates. Int. J. Antimicrob. Agents 2004, 23, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Yam, T.S.; Hamilton-Miller, J.M.; Shah, S. The effect of a component of tea (Camellia sinensis) on methicillin resistance, PBP2’ synthesis, and beta-lactamase production in Staphylococcus aureus. J. Antimicrob. Chemother. 1998, 42, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Hamilton-Miller, J.M.; Shah, S. Activity of the tea component epicatechin gallate and analogues against methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2000, 46, 852–853. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Mandalari, G.; Bisignano, C.; Filocamo, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Polyphenolic content and biological properties of Avola almond (Prunus dulcis Mill. D.A. Webb) skin and its industrial by products. Ind. Crops Prod. 2016, 83, 283–293. [Google Scholar] [CrossRef]
- Smeriglio, A.; Denaro, M.; Barreca, D.; Calderaro, A.; Bisignano, C.; Ginestra, G.; Bellocco, E.; Trombetta, D. In Vitro Evaluation of the Antioxidant, Cytoprotective, and Antimicrobial Properties of Essential Oil from Pistacia vera L. Variety Bronte Hull. Int. J. Mol. Sci. 2017, 18, 1212. [Google Scholar] [CrossRef] [PubMed]
- Bisignano, C.; Mandalari, G.; Smeriglio, A.; Trombetta, D.; Pizzo, M.M.; Pennisi, R.; Sciortino, M.T. Almond Skin Extracts Abrogate HSV-1 Replication by Blocking Virus Binding to the Cell. Viruses 2017, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Susceptibility Testing; Twentieth Informational Supplement; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- Nostro, A.; Guerrini, A.; Marino, A.; Tacchini, M.; Di Giulio, M.; Grandini, A.; Akin, M.; Cellini, L.; Bisignano, G.; Saraçoğlu, H.T. In vitro activity of plant extracts against biofilm-producing food-related bacteria. Int. J. Food Microbiol. 2016, 238, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.H.; Martinez Garcia, R.; Abrão, F.; Fernandez, Y.A.; Pires, R.H.; Ambrósio, S.R.; Veneziani, R.C.S.; Martins, C.H.G. Bactericidal Kinetics and Antibiofilm Efficacy of Pimarane-Type Diterpenes from Viguiera arenaria Against Cariogenic Bacteria. Pharmacogn. J. 2018, 10, 429–434. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Compound | NPRE | RPRE |
|---|---|---|
| Hydroxybenzoic acids | ||
| Gallic acid | 0.99 ± 0.035 | 1.77 ± 0.055 |
| Protocathecuic acid | 1.01 ± 0.042 | 1.08 ± 0.047 |
| Hydroxybenzoic acid | 0.17 ±0.011 | 0.19 ± 0.008 |
| Vanillic acid | 0.02 ± 0.001 | - |
| Hydroxycinnamic acids | ||
| Chlorogenic acid | 0.15 ± 0.011 | 0.19 ± 0.010 |
| Caffeic acid | 0.71 ± 0.032 | 0.01 ± 0.001 |
| Cumaric acid | 0.12 ± 0.010 | 0.02 ± 0.001 |
| Flavanones | ||
| Eryodictiol | 0.23 ± 0.012 | 0.21 ± 0.013 |
| Eryodictiol-7-O-glucoside | 1.34 ± 0.088 | 2.19 ± 0.074 |
| Naringenin | 0.05 ± 0.002 | 0.03 ± 0.002 |
| Naringin | 0.10 ± 0.004 | 0.04 ± 0.001 |
| Flavonols | ||
| Kaempferol-3-O-rutinoside | 0.05 ± 0.002 | 0.04 ± 0.002 |
| Quercetin | 0.18 ± 0.010 | 0.07 ± 0.003 |
| Quercetin-3-O-rutinoside | 0.30 ± 0.016 | 0.13 ± 0.008 |
| Quercetin-3-O-glucoside | 0.24 ± 0.012 | 0.04 ± 0.002 |
| Flavones | ||
| Amentoflavone | 0.17 ± 0.007 | 0.05 ± 0.003 |
| Luteolin | 0.04 ± 0.002 | 0.04 ± 0.002 |
| Apigenin | 0.06 ± 0.001 | 0.01 ± 0.001 |
| Isoflavones | ||
| Daidzein | 0.06 ± 0.002 | 0.10 ± 0.005 |
| Genistein | 0.01 ± 0.001 | 0.01 ± 0.004 |
| Flavanols | ||
| Epicatechin | 0.07 ± 0.002 | 0.04 ± 0.002 |
| Catechin | 2.04 ± 0.080 | 0.69 ± 0.035 |
| Total amount | 8.11 | 6.95 |
| Strain | Origin | Coagulase | Lipase | API System |
|---|---|---|---|---|
| 1 | knee | + | − | S. aureus |
| 2 | hip | + | + | S. aureus |
| 3 | knee | + | + | S. aureus |
| 4 | OOS | + | + | S. aureus |
| 5 | knee | − | − | S. sp. |
| 6 | OOS | − | − | S. xylosus |
| 7 | hip | + | − | S. aureus |
| 8 | OOS | + | + | S. aureus |
| 9 | knee | + | + | S. aureus |
| 10 | knee | + | − | S. aureus |
| 11 | hip | + | + | S. aureus |
| 12 | knee | − | − | S. epidermidis |
| 13 | OOS | − | − | S. lugdunensis |
| 14 | knee | − | − | S. epidermidis |
| 15 | hip | + | − | S. aureus |
| 16 | OOS | + | + | S. aureus |
| 17 | OOS | + | + | S. aureus |
| 18 | knee | + | + | S. aureus |
| 19 | knee | + | − | S. aureus |
| 20 | knee | + | − | S. aureus |
| 21 | knee | + | − | S. aureus |
| 22 | knee | − | + | S. hominis |
| 23 | hip | + | − | S. aureus |
| 24 | knee | − | + | S. hominis |
| 25 | knee | + | + | S. aureus |
| 26 | OOS | + | + | S. aureus |
| 27 | hip | + | + | S. aureus |
| 28 | OOS | + | + | S. aureus |
| 29 | hip | − | − | S. lugdunensis |
| 30 | knee | + | − | S. aureus |
| 31 | knee | + | − | S. aureus |
| ATCC 6538P | + | + | S. aureus |
| Strain | NPRE | RPRE |
|---|---|---|
| 1 | 2000 | 2000 |
| 2 | >2000 | 2000 |
| 3 | >2000 | 2000 |
| 4 | 250 | 500 |
| 5 | >2000 | >2000 |
| 6 | 2000 | 2000 |
| 7 | >2000 | 1000 |
| 8 | >2000 | 2000 |
| 9 | 2000 | 2000 |
| 10 | 2000 | 2000 |
| 11 | >2000 | 1000 |
| 12 | 2000 | 2000 |
| 13 | 2000 | 2000 |
| 14 | 62.5 | 500 |
| 15 | 1000 | 1000 |
| 16 | >2000 | >2000 |
| 17 | >2000 | 1000 |
| 18 | 2000 | 2000 |
| 19 | 2000 | 2000 |
| 20 | 2000 | 2000 |
| 21 | >2000 | 2000 |
| 22 | >2000 | >2000 |
| 23 | 2000 | 2000 |
| 24 | 2000 | 2000 |
| 25 | >2000 | 2000 |
| 26 | >2000 | 2000 |
| 27 | >2000 | >2000 |
| 28 | >2000 | >2000 |
| 29 | >2000 | >2000 |
| 30 | 1000 | 1000 |
| 31 | 2000 | 2000 |
| ATCC 6538P | 125 | 31.25 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
La Camera, E.; Bisignano, C.; Crisafi, G.; Smeriglio, A.; Denaro, M.; Trombetta, D.; Mandalari, G. Biochemical Characterization of Clinical Strains of Staphylococcus spp. and Their Sensitivity to Polyphenols-Rich Extracts from Pistachio (Pistacia vera L.). Pathogens 2018, 7, 82. https://doi.org/10.3390/pathogens7040082
La Camera E, Bisignano C, Crisafi G, Smeriglio A, Denaro M, Trombetta D, Mandalari G. Biochemical Characterization of Clinical Strains of Staphylococcus spp. and Their Sensitivity to Polyphenols-Rich Extracts from Pistachio (Pistacia vera L.). Pathogens. 2018; 7(4):82. https://doi.org/10.3390/pathogens7040082
Chicago/Turabian StyleLa Camera, Erminia, Carlo Bisignano, Giuseppe Crisafi, Antonella Smeriglio, Marcella Denaro, Domenico Trombetta, and Giuseppina Mandalari. 2018. "Biochemical Characterization of Clinical Strains of Staphylococcus spp. and Their Sensitivity to Polyphenols-Rich Extracts from Pistachio (Pistacia vera L.)" Pathogens 7, no. 4: 82. https://doi.org/10.3390/pathogens7040082
APA StyleLa Camera, E., Bisignano, C., Crisafi, G., Smeriglio, A., Denaro, M., Trombetta, D., & Mandalari, G. (2018). Biochemical Characterization of Clinical Strains of Staphylococcus spp. and Their Sensitivity to Polyphenols-Rich Extracts from Pistachio (Pistacia vera L.). Pathogens, 7(4), 82. https://doi.org/10.3390/pathogens7040082