New Insights on the Pathogenesis of Takayasu Arteritis: Revisiting the Microbial Theory


Abstract
1. Introduction
2. Takayasu Arteritis: An Overview
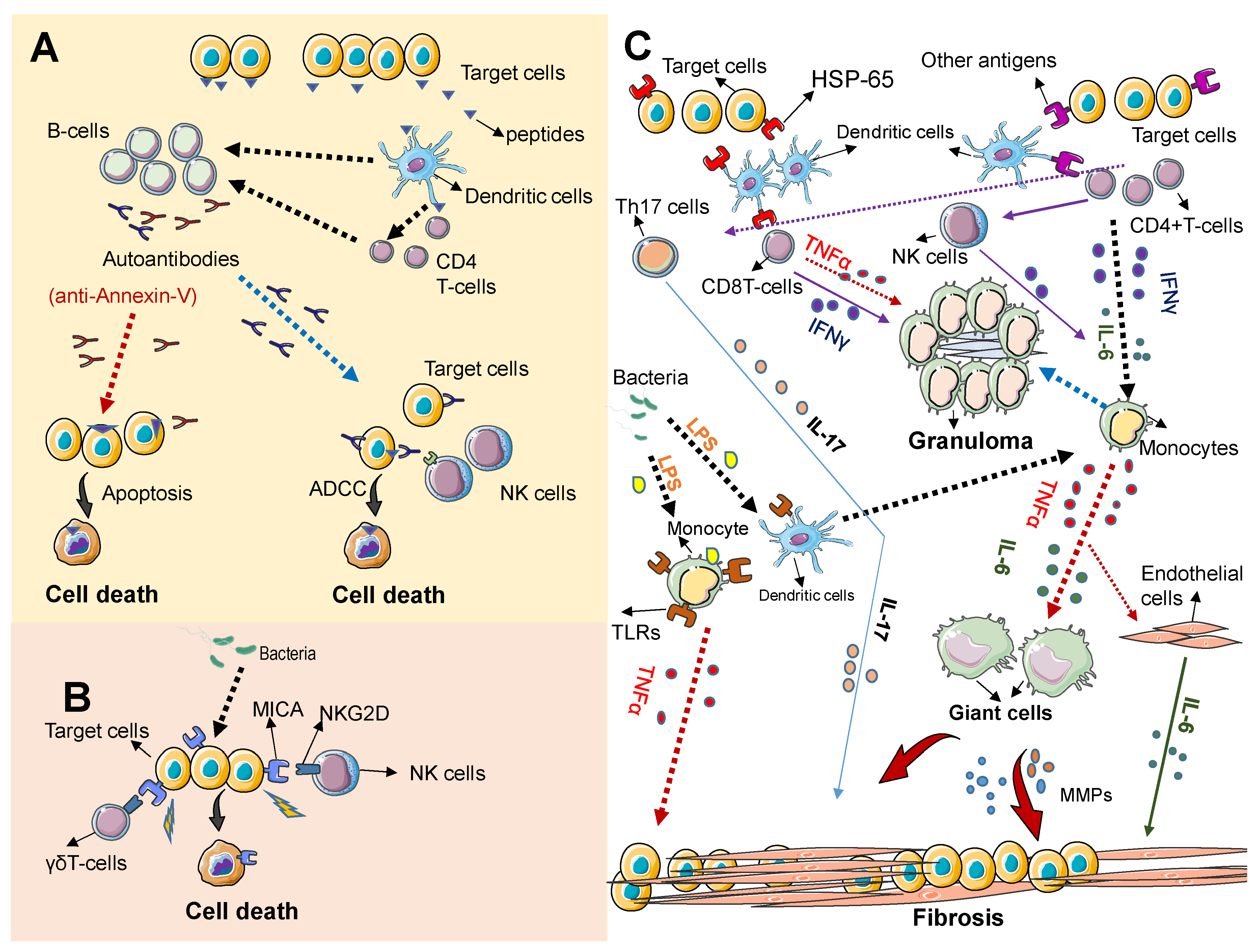
3. Immunopathogenesis of TAK
4. Infections and TAK
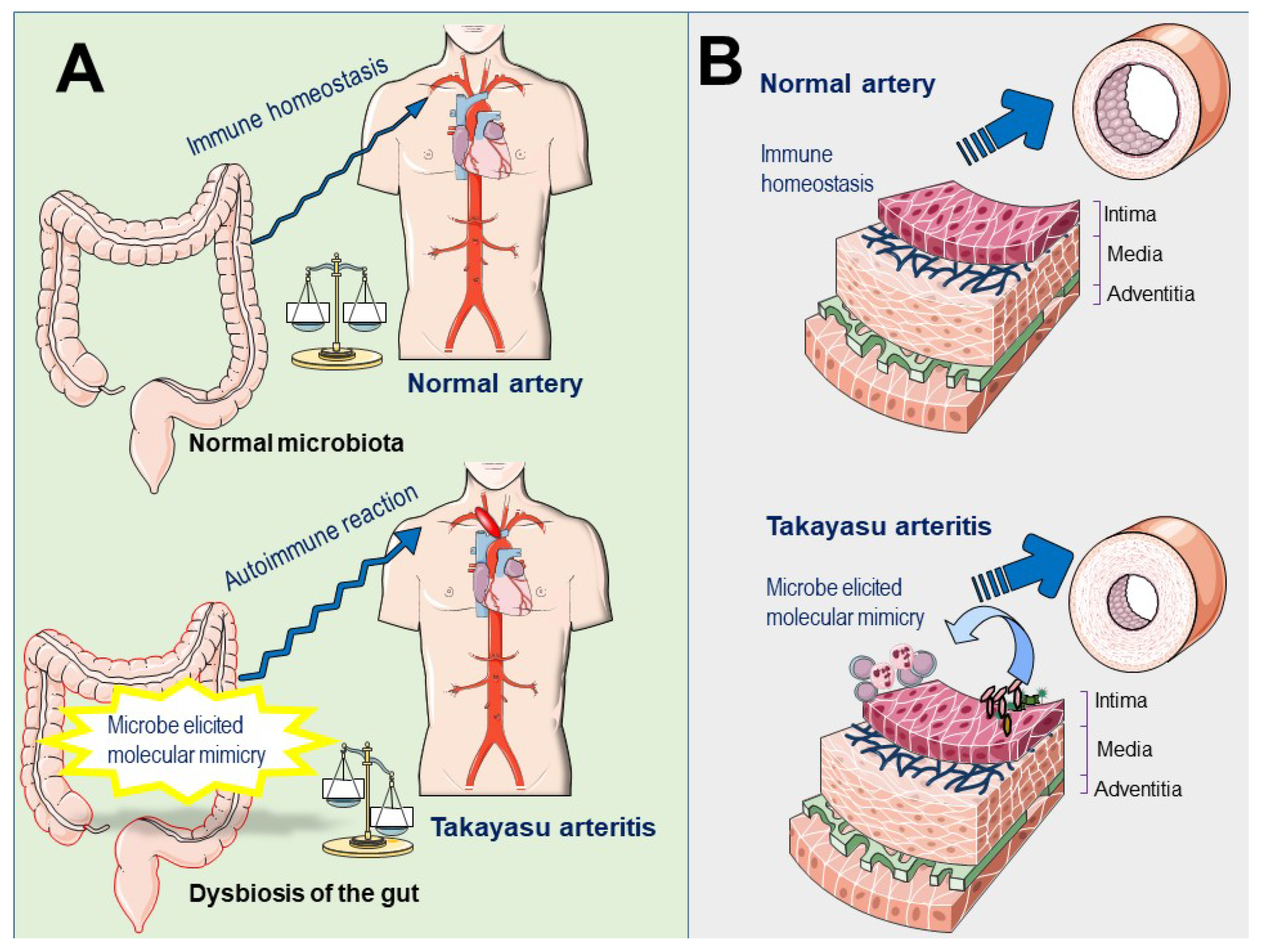
5. Is Microbiota Implicated in the Pathogenesis of TAK?
6. Concluding Remarks and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Terao, C. History of Takayasu arteritis and Dr. Mikito Takayasu. Int. J. Rheum. Dis. 2014, 17, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Numano, F. The story of Takayasu arteritis. Rheumatology (Oxford) 2002, 41, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, K.; Ijiri, S.; Tagawa, S.; Shimizu, K. Takayasu disease on the centenary of its discovery. Jpn. J. Ophthalmol. 2009, 53, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Arend, W.P.; Michel, B.A.; Bloch, D.A.; Hunder, G.G.; Calabrese, L.H.; Edworthy, S.M.; Fauci, A.S.; Leavitt, R.Y.; Lie, J.T.; Lightfoot, R.W. The American College of Rheumatology 1990 criteria for the classification of Takayasu arteritis. Arthritis Rheum. 1990, 33, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Terao, C. Revisited HLA and non-HLA genetics of Takayasu arteritis—Where are we? J. Hum. Genet. 2016, 61, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Mirault, T.; Guillet, H.; Messas, E. Immune response in Takayasu arteritis. Presse Med. 2017, 46, e189–e196. [Google Scholar] [CrossRef] [PubMed]
- Mekinian, A.; Comarmond, C.; Resche-Rigon, M.; Mirault, T.; Kahn, J.E.; Lambert, M.; Sibilia, J.; Néel, A.; Cohen, P.; Hie, M.; et al. Network, Efficacy of Biological-Targeted Treatments in Takayasu Arteritis: Multicenter, Retrospective Study of 49 Patients. Circulation 2015, 132, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Ferfar, Y.; Mirault, T.; Desbois, A.C.; Comarmond, C.; Messas, E.; Savey, L.; Domont, F.; Cacoub, P.; Saadoun, D. Biotherapies in large vessel vasculitis. Autoimmun. Rev. 2016, 15, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Clifford, A.; Hoffman, G.S. Evidence for a vascular microbiome and its role in vessel health and disease. Curr. Opin. Rheumatol. 2015, 27, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Talotta, R.; Atzeni, F.; Ditto, M.C.; Gerardi, M.C.; Sarzi-Puttini, P. The Microbiome in Connective Tissue Diseases and Vasculitides: An Updated Narrative Review. J. Immunol. Res. 2017, 6836498. [Google Scholar] [CrossRef] [PubMed]
- Onen, F.; Akkoc, N. Epidemiology of Takayasu arteritis. Presse Med. 2017, 46, e197–e203. [Google Scholar] [CrossRef] [PubMed]
- Renauer, P.; Sawalha, A.H. The genetics of Takayasu arteritis. Presse Med. 2017, 46, e179–e187. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Luan, H.; Li, L.; Zeng, X.; Wang, T.; Li, Y.; Yuan, H. Relationship of HLA-B*51 and HLA-B*52 alleles and TNF-α-308A/G polymorphism with susceptibility to Takayasu arteritis: A meta-analysis. Clin. Rheumatol. 2017, 36, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Origuchi, T.; Fukui, S.; Umeda, M.; Nishino, A.; Nakashima, Y.; Koga, T.; Kawashiri, S.Y.; Iwamoto, N.; Ichinose, K.; Tamai, M.; et al. The Severity of Takayasu Arteritis Is Associated with the HLA-B52 Allele in Japanese Patients. Tohoku J. Exp. Med. 2016, 239, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Takamura, C.; Ohhigashi, H.; Ebana, Y.; Isobe, M. New human leukocyte antigen risk allele in Japanese patients with Takayasu arteritis. Circ. J. 2012, 76, 1697–1702. [Google Scholar] [CrossRef] [PubMed]
- González-Galarza, F.F.; Takeshita, L.Y.; Santos, E.J.; Kempson, F.; Maia, M.H.; Da Silva, A.L.; Teles e Silva, A.L.; Ghattaoraya, G.S.; Alfirevic, A.; Jones, A.R.; et al. Allele frequency net 2015 update: New features for HLA epitopes, KIR and disease and HLA adverse drug reaction associations. Nucleic Acids Res. 2015, 43, D784–D788. [Google Scholar] [CrossRef] [PubMed]
- Danda, D.; Goel, R.; Danda, S.; Mohan, H.; Joseph, G.; Kabeerdoss, J.; Nath, S.K. Interleukin-17F and interleukin-6 gene polymorphisms in Asian Indian patients with Takayasu arteritis. Hum. Immunol. 2017, 78, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Chen, S.; Li, P.; Li, J.; Wu, Z.; Li, Y.; Li, L.; Yuan, H.; Tian, X.; Zhang, F. Single nucleotide polymorphisms of IL12B are associated with Takayasu arteritis in Chinese Han population. Rheumatol. Int. 2017, 37, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, F.; Chen, Z.; Yang, Y.; Zhao, J.; Li, M.; Tian, X.; Zeng, X. The clinical characteristics of Chinese Takayasu’s arteritis patients: A retrospective study of 411 patients over 24 years. Arthritis Res. Ther. 2017, 19, 107. [Google Scholar] [CrossRef] [PubMed]
- Direskeneli, H. Clinical assessment in Takayasu’s arteritis: Major challenges and controversies. Clin. Exp. Rheumatol. 2017, 35, 189–193. [Google Scholar] [PubMed]
- Mirouse, A.; Biard, L.; Comarmond, C.; Lambert, M.; Mekinian, A.; Ferfar, Y.; Kahn, J.E.; Benhamou, Y.; Chiche, L.; Koskas, F.; et al. Network, Overall survival and mortality risk factors in Takayasu’s arteritis: A multicenter study of 318 patients. J. Autoimmun. 2018. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.; Butrico, L.; Fugetto, F.; Chibireva, M.D.; Malva, A.; De Caridi, G.; Massara, M.; Barbetta, A.; Cannistrà, M.; De Franciscis, S. Updates in Pathophysiology, Diagnosis and Management of Takayasu Arteritis. Ann. Vasc. Surg. 2016, 35, 210–225. [Google Scholar] [CrossRef] [PubMed]
- Seyahi, E. Takayasu arteritis: An update. Curr. Opin. Rheumatol. 2017, 29, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Miyata, T.; Tanemoto, K. Current Clinical Features of New Patients With Takayasu Arteritis Observed From Cross-Country Research in Japan: Age and Sex Specificity. Circulation 2015, 132, 1701–1709. [Google Scholar] [CrossRef] [PubMed]
- Bicakcigil, M.; Aksu, K.; Kamali, S.; Ozbalkan, Z.; Ates, A.; Karadag, O.; Ozer, H.T.; Seyahi, E.; Akar, S.; Onen, F.; et al. Takayasu’s arteritis in Turkey–clinical and angiographic features of 248 patients. Clin. Exp. Rheumatol. 2009, 27, S59–S64. [Google Scholar] [PubMed]
- Soto, M.E.; Espinola, N.; Flores-Suarez, L.F.; Reyes, P.A. Takayasu arteritis: Clinical features in 110 Mexican Mestizo patients and cardiovascular impact on survival and prognosis. Clin. Exp. Rheumatol. 2008, 26, S9–S15. [Google Scholar] [CrossRef]
- Alibaz-Oner, F.; Direskeneli, H. Update on Takayasu’s arteritis. Presse Med. 2015, 44, e259–e265. [Google Scholar] [CrossRef] [PubMed]
- Comarmond, C.; Biard, L.; Lambert, M.; Mekinian, A.; Ferfar, Y.; Kahn, J.E.; Benhamou, Y.; Chiche, L.; Koskas, F.; Cluzel, P.; et al. Network, Long-Term Outcomes and Prognostic Factors of Complications in Takayasu Arteritis: A Multicenter Study of 318 Patients. Circulation 2017, 136, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Keser, G.; Aksu, K.; Direskeneli, H. Takayasu arteritis: An update. Turk. J. Med. Sci. 2018, 48, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Keser, G.; Direskeneli, H.; Aksu, K. Management of Takayasu arteritis: A systematic review. Rheumatology (Oxford) 2014, 53, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Langford, C.A.; Cuthbertson, D.; Ytterberg, S.R.; Khalidi, N.; Monach, P.A.; Carette, S.; Seo, P.; Moreland, L.W.; Weisman, M.; Koening, C.L. A Randomized, Double-Blind Trial of Abatacept (CTLA-4Ig) for the Treatment of Takayasu Arteritis. Arthritis Rheumatol. 2017, 69, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Nakaoka, Y.; Isobe, M.; Takei, S.; Tanaka, Y.; Ishii, T.; Yokota, S.; Nomura, A.; Yoshida, S.; Nishimoto, N. Efficacy and safety of tocilizumab in patients with refractory Takayasu arteritis: Results from a randomised, double-blind, placebo-controlled, phase 3 trial in Japan (the TAKT study). Ann. Rheum. Dis. 2018, 77, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Kotecha, R.; Takami, A.; Espinoza, J.L. Dietary phytochemicals and cancer chemoprevention: A review of the clinical evidence. Oncotarget 2016, 7, 52517–52529. [Google Scholar] [CrossRef] [PubMed]
- Shao, N.; Jia, H.; Li, Y.; Li, J. Curcumin improves treatment outcome of Takayasu arteritis patients by reducing TNF-α: A randomized placebo-controlled double-blind clinical trial. Immunol. Res. 2017, 65, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Hua, M.; Xu, Q.; Ren, T. Resveratrol improves treatment outcome and laboratory parameters in patients with Takayasu arteritis: A randomized double-blind and placebo-controlled trial. Immunobiology 2017, 222, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Muñoz, M.; Fischer, A.; Plickert, R.; Haag, L.M.; Otto, B.; Kühl, A.A.; Loddenkemper, C.; Göbel, U.B.; Heimesaat, M.M. Anti-inflammatory effects of resveratrol, curcumin and simvastatin in acute small intestinal inflammation. PLoS ONE 2010, 5, e15099. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.L.; Trung, L.Q.; Inaoka, P.T.; Yamada, K.; An, D.T.; Mizuno, S.; Nakao, S.; Takami, A. The Repeated Administration of Resveratrol Has Measurable Effects on Circulating T-Cell Subsets in Humans. Oxid. Med. Cell. Longev. 2017, 6781872. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Sun, Y.; Ma, L.; Chen, H.; Wei, L.; Wu, W.; Ji, Z.; Zhang, Z.; Zhao, Z.; Hou, J.; et al. The critical role of IL-6 in the pathogenesis of Takayasu arteritis. Clin. Exp. Rheumatol. 2016, 34, S21–S27. [Google Scholar] [PubMed]
- Saadoun, D.; Garrido, M.; Comarmond, C.; Desbois, A.C.; Domont, F.; Savey, L.; Terrier, B.; Geri, G.; Rosenzwajg, M.; Klatzmann, D.; et al. Cacoub, Th1 and Th17 cytokines drive inflammation in Takayasu arteritis. Arthritis Rheumatol. 2015, 67, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Nityanand, S.; Giscombe, R.; Srivastava, S.; Hjelmström, P.; Sanjeevi, C.B.; Sinha, N.; Grunewald, J.; Lefvert, A.K. A bias in the alphabeta T cell receptor variable region gene usage in Takayasu’s arteritis. Clin. Exp. Immunol. 1997, 107, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.G.; Salmon, M.; Scott, D.L.; Blann, A.; Bacon, P.A.; Walton, K.W.; Oakland, C.D.; Slaney, G.F. Takayasu’s arteritis: A pathogenetic role for cytotoxic T lymphocytes? Clin. Rheumatol. 1986, 5, 517–522. [Google Scholar] [PubMed]
- Sagar, S.; Ganguly, N.K.; Koicha, M.; Sharma, B.K. Immunopathogenesis of Takayasu arteritis. Heart Vessels Suppl. 1992, 7, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.R.; Bruneval, P.; Angelini, A.; Bartoloni, G.; Basso, C.; Batoroeva, L.; Buja, L.M.; Butany, J.; D’Amati, G.; Fallon, J.T.; et al. Veinot, Consensus statement on surgical pathology of the aorta from the Society for Cardiovascular Pathology and the Association for European Cardiovascular Pathology: I. Inflammatory diseases. Cardiovasc. Pathol. 2015, 24, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.L.; Nguyen, V.H.; Ichimura, H.; Pham, T.T.; Nguyen, C.H.; Pham, T.V.; Elbadry, M.I.; Yoshioka, K.; Tanaka, J.; Trung, L.Q.; et al. A functional polymorphism in the NKG2D gene modulates NK-cell cytotoxicity and is associated with susceptibility to Human Papilloma Virus-related cancers. Sci. Rep. 2016, 6, 39231. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.L.; Minami, M. Sensing Bacterial-Induced DNA Damaging Effects. Front. Immunol. 2018, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Seko, Y.; Sugishita, K.; Sato, O.; Takagi, A.; Tada, Y.; Matsuo, H.; Yagita, H.; Okumura, K.; Nagai, R. Expression of costimulatory molecules (4-1BBL and Fas) and major histocompatibility class I chain-related A (MICA) in aortic tissue with Takayasu’s arteritis. J. Vasc. Res. 2004, 41, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Guerra, N.; Pestal, K.; Juarez, T.; Beck, J.; Tkach, K.; Wang, L.; Raulet, D.H. A selective role of NKG2D in inflammatory and autoimmune diseases. Clin. Immunol. 2013, 149, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Kumar Chauhan, S.; Kumar Tripathy, N.; Sinha, N.; Singh, M.; Nityanand, S. Cellular and humoral immune responses to mycobacterial heat shock protein-65 and its human homologue in Takayasu’s arteritis. Clin. Exp. Immunol. 2004, 138, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Seko, Y.; Minota, S.; Kawasaki, A.; Shinkai, Y.; Maeda, K.; Yagita, H.; Okumura, K.; Sato, O.; Takagi, A.; Tada, Y. Perforin-secreting killer cell infiltration and expression of a 65-kD heat-shock protein in aortic tissue of patients with Takayasu’s arteritis. J. Clin. Investig. 1994, 93, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, B.F.; Mumtaz, I.M.; Loddenkemper, K.; Bruns, A.; Sengler, C.; Hermann, K.G.; Maza, S.; Keitzer, R.; Burmester, G.R.; Buttgereit, F.; et al. Takayasu arteritis is characterised by disturbances of B cell homeostasis and responds to B cell depletion therapy with rituximab. Ann. Rheum. Dis. 2012, 71, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Hadjadj, J.; Canaud, G.; Mirault, T.; Samson, M.; Bruneval, P.; Régent, A.; Goulvestre, C.; Witko-Sarsat, V.; Costedoat-Chalumeau, N.; Guillevin, L.; et al. mTOR pathway is activated in endothelial cells from patients with Takayasu arteritis and is modulated by serum immunoglobulin G. Rheumatology (Oxford) 2018, 57, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Alibaz-Oner, F.; Yentür, S.P.; Saruhan-Direskeneli, G.; Direskeneli, H. Serum cytokine profiles in Takayasu’s arteritis: Search for biomarkers. Clin. Exp. Rheumatol. 2015, 33, S32–S35. [Google Scholar]
- Pan, L.L.; Du, J.; Gao, N.; Liao, H.; Wan, J.; Ci, W.P.; Yang, C.; Wang, T. IL-9-producing Th9 cells may participate in pathogenesis of Takayasu’s arteritis. Clin. Rheumatol. 2016, 35, 3031–3036. [Google Scholar] [CrossRef] [PubMed]
- Tamura, N.; Maejima, Y.; Tezuka, D.; Takamura, C.; Yoshikawa, S.; Ashikaga, T.; Hirao, K.; Isobe, M. Profiles of serum cytokine levels in Takayasu arteritis patients: Potential utility as biomarkers for monitoring disease activity. J. Cardiol. 2017, 70, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Novikov, P.I.; Smitienko, I.O.; Sokolova, M.V.; Alibaz-Oner, F.; Kaymaz-Tahra, S.; Direskeneli, H.; Moiseev, S.V. Certolizumab pegol in the treatment of Takayasu arteritis. Rheumatology (Oxford) 2018. [Google Scholar] [CrossRef] [PubMed]
- Tombetti, E.; Di Chio, M.C.; Sartorelli, S.; Bozzolo, E.; Sabbadini, M.G.; Manfredi, A.A.; Baldissera, E. Anti-cytokine treatment for Takayasu arteritis: State of the art. Intractable Rare Dis. Res. 2014, 3, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Joshi, L.; Ponnana, M.; Sivangala, R.; Chelluri, L.K.; Nallari, P.; Penmetsa, S.; Valluri, V.; Gaddam, S. Evaluation of TNF-α, IL-10 and IL-6 Cytokine Production and Their Correlation with Genotype Variants amongst Tuberculosis Patients and Their Household Contacts. PLoS ONE 2015, 10, e0137727. [Google Scholar] [CrossRef] [PubMed]
- Verbon, A.; Juffermans, N.; Van Deventer, S.J.; Speelman, P.; Van Deutekom, H.; Van Der Poll, T. Serum concentrations of cytokines in patients with active tuberculosis (TB) and after treatment. Clin. Exp. Immunol. 1999, 115, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Dorhoi, A.; Kaufmann, S.H. Tumor necrosis factor alpha in mycobacterial infection. Semin. Immunol. 2014, 26, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Kang, Y.A.; Leem, A.Y.; Park, M.S.; Kim, Y.S.; Kim, S.K.; Chang, J.; Kim, S.Y. Active Tuberculosis Incidence and Characteristics in Patients Treated with Tumor Necrosis Factor Antagonists According to Latent Tuberculosis Infection. Sci. Rep. 2017, 7, 6473. [Google Scholar] [CrossRef] [PubMed]
- Sandhya, P.; Danda, S.; Danda, D.; Lonarkar, S.; Luke, S.S.; Sinha, S.; Joseph, G. Tumour necrosis factor (TNF)-α-308 gene polymorphism in Indian patients with Takayasu’s arteritis—A pilot study. Indian J. Med. Res. 2013, 137, 749–752. [Google Scholar] [PubMed]
- Alibaz-Oner, F.; Aksu, K.; Yentur, S.P.; Keser, G.; Saruhan-Direskeneli, G.; Direskeneli, H. Plasma pentraxin-3 levels in patients with Takayasu’s arteritis during routine follow-up. Clin. Exp. Rheumatol. 2016, 34, S73–S76. [Google Scholar] [PubMed]
- Garlanda, C.; Hirsch, E.; Bozza, S.; Salustri, A.; De Acetis, M.; Nota, R.; Maccagno, A.; Riva, F.; Bottazzi, B.; Peri, G.; et al. Non-redundant role of the long pentraxin PTX3 in anti-fungal innate immune response. Nature 2002, 420, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Bottazzi, B.; Santini, L.; Savino, S.; Giuliani, M.M.; Dueñas Díez, A.I.; Mancuso, G.; Beninati, C.; Sironi, M.; Valentino, S.; Deban, L.; et al. Mantovani, Recognition of Neisseria meningitidis by the long pentraxin PTX3 and its role as an endogenous adjuvant. PLoS ONE 2015, 10, e0120807. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Jeon, J.; Lee, J.H.; Jin, S.; Ha, U.H. Pseudomonas aeruginosa GroEL Stimulates Production of PTX3 by Activating the NF-κB Pathway and Simultaneously Downregulating MicroRNA-9. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.Y.; Lee, G.Y.; Jang, S.Y.; Gwag, H.B.; Choi, S.H.; Jeon, E.S.; Cha, H.S.; Sung, K.; Kim, Y.W.; Kim, S.M.; et al. Comparison of clinical characteristics in patients with Takayasu arteritis with and without concomitant tuberculosis. Heart Vessel. 2016, 31, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Picard, C.; Parrot, A.; Boussaud, V.; Lavolé, A.; Saidi, F.; Mayaud, C.; Carette, M.F. Massive hemoptysis due to Rasmussen aneurysm: Detection with helicoidal CT angiography and successful steel coil embolization. Intens. Care Med. 2003, 29, 1837–1839. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.E.; Del Carmen Ávila-Casado, M.; Huesca-Gómez, C.; Alarcon, G.V.; Castrejon, V.; Soto, V.; Hernandez, S.; Espinola-Zavaleta, N.; Vallejo, M.; Reyes, P.A.; et al. Detection of IS6110 and HupB gene sequences of Mycobacterium tuberculosis and bovis in the aortic tissue of patients with Takayasu’s arteritis. BMC Infect. Dis. 2012, 12, 194. [Google Scholar] [CrossRef] [PubMed]
- Karadag, O.; Aksu, K.; Sahin, A.; Zihni, F.Y.; Sener, B.; Inanc, N.; Kalyoncu, U.; Aydin, S.Z.; Ascioglu, S.; Ocakci, P.T.; et al. Assessment of latent tuberculosis infection in Takayasu arteritis with tuberculin skin test and Quantiferon-TB Gold test. Rheumatol. Int. 2010, 30, 1483–1487. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, L.; Cambau, E.; Brocheriou, I.; Koskas, F.; Kieffer, E.; Piette, J.C.; Amoura, Z. Absence of Mycobacterium tuberculosis in arterial lesions from patients with Takayasu’s arteritis. J. Rheumatol. 2009, 36, 1682–1685. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, E.S.; De Souza, A.W.; Leão, S.C.; Levy-Neto, M.; De Oliveira, R.S.; Drake, W.; De Franco, M.F.; Saldiva, P.H.; Gutierrez, P.S.; Andrade, L.E. Absence of mycobacterial DNA in peripheral blood and artery specimens in patients with Takayasu arteritis. Clin. Rheumatol. 2017, 36, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Belizna, C.C.; Hamidou, M.A.; Levesque, H.; Guillevin, L.; Shoenfeld, Y. Infection and vasculitis. Rheumatology (Oxford) 2009, 48, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Chapelon-Abric, C.; Saadoun, D.; Marie, I.; Comarmond, C.; Desbois, A.C.; Domont, F.; Savey, L.; Cacoub, P. Sarcoidosis with Takayasu arteritis: A model of overlapping granulomatosis. A report of seven cases and literature review. Int. J. Rheum. Dis. 2017. [Google Scholar] [CrossRef] [PubMed]
- Kuberski, T.; Yourison, I. Coccidioidomycosis A Cause of Sarcoidosis. Open Forum Infect. Dis. 2017, 4, ofw117. [Google Scholar] [CrossRef] [PubMed]
- Saidha, S.; Sotirchos, E.S.; Eckstein, C. Etiology of sarcoidosis: Does infection play a role? Yale J. Biol. Med. 2012, 85, 133–141. [Google Scholar] [PubMed]
- Terao, C.; Matsumura, T.; Yoshifuji, H.; Kirino, Y.; Maejima, Y.; Nakaoka, Y.; Takahashi, M.; Amiya, E.; Tamura, N.; Nakajima, T.; et al. Takayasu arteritis and ulcerative colitis: High rate of co-occurrence and genetic overlap. Arthritis Rheumatol. 2015, 67, 2226–2232. [Google Scholar] [CrossRef] [PubMed]
- Sy, A.; Khalidi, N.; Dehghan, N.; Barra, L.; Carette, S.; Cuthbertson, D.; Hoffman, G.S.; Koening, C.L.; Langford, C.A.; McAlear, C.; et al. Vasculitis in patients with inflammatory bowel diseases: A study of 32 patients and systematic review of the literature. Semin. Arthritis Rheum. 2016, 45, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, Y.; Uehara, T.; Kawana, S. Cutaneous Vasculitis in Cogan’s Syndrome: A Report of Two Cases Associated with Chlamydia Infection. J. Nippon. Med. Sch. 2018, 85, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Pryshchep, O.; Ma-Krupa, W.; Younge, B.R.; Goronzy, J.J.; Weyand, C.M. Vessel-specific Toll-like receptor profiles in human medium and large arteries. Circulation 2008, 118, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Ma-Krupa, W.; Gewirtz, A.T.; Younge, B.R.; Goronzy, J.J.; Weyand, C.M. Toll-like receptors 4 and 5 induce distinct types of vasculitis. Circ. Res. 2009, 104, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Dal Canto, A.J.; Virgin, H.W.; Speck, S.H. Ongoing viral replication is required for gammaherpesvirus 68-induced vascular damage. J. Virol. 2000, 74, 11304–11310. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Ohsawa, M.; Hu, S.X.; Kanno, H.; Aozasa, K.; Nose, M. Large-vessel arteritis associated with chronic active Epstein-Barr virus infection. Arthritis Rheum. 1998, 41, 369–373. [Google Scholar] [CrossRef]
- Khan, G.; Hashim, M.J. Global burden of deaths from Epstein-Barr virus attributable malignancies 1990–2010. Infect. Agent Cancer 2014, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Palm, N.W.; De Zoete, M.R.; Flavell, R.A. Immune-microbiota interactions in health and disease. Clin. Immunol. 2015, 159, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Elson, C.O.; Alexander, K.L. Host-microbiota interactions in the intestine. Dig. Dis. 2015, 33, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.L.; Kotecha, R.; Nakao, S. Microbe-Induced Inflammatory Signals Triggering Acquired Bone Marrow Failure Syndromes. Front. Immunol. 2017, 8, 186. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.L.; Elbadry, M.I.; Nakao, S. An altered gut microbiota may trigger autoimmune-mediated acquired bone marrow failure syndromes. Clin. Immunol. 2016, 171, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Yurkovetskiy, L.A.; Pickard, J.M.; Chervonsky, A.V. Microbiota and autoimmunity: Exploring new avenues. Cell Host Microbe 2015, 17, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.I.; Obeid, M.T.; Jouma, M.J.; Moasis, G.A.; Al-Richane, W.L.; Kindermann, I.; Boehm, M.; Roemer, K.; Mueller-Lantzsch, N.; Gärtner, B.C. Detection of herpes simplex virus, cytomegalovirus and Epstein-Barr virus DNA in atherosclerotic plaques and in unaffected bypass grafts. J. Clin. Virol. 2005, 32, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Weiss, T.W.; Kvakan, H.; Kaun, C.; Prager, M.; Speidl, W.S.; Zorn, G.; Pfaffenberger, S.; Huk, I.; Maurer, G.; Huber, K.; et al. No evidence for a direct role of Helicobacter pylori and Mycoplasma pneumoniae in carotid artery atherosclerosis. J. Clin. Pathol. 2006, 59, 1186–1190. [Google Scholar] [CrossRef] [PubMed]
- Reszka, E.; Jegier, B.; Wasowicz, W.; Lelonek, M.; Banach, M.; Jaszewski, R. Detection of infectious agents by polymerase chain reaction in human aortic wall. Cardiovasc. Pathol. 2008, 17, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Kinumaki, A.; Sekizuka, T.; Hamada, H.; Kato, K.; Yamashita, A.; Kuroda, M. Characterization of the gut microbiota of Kawasaki disease patients by metagenomic analysis. Front. Microbiol. 2015, 6, 824. [Google Scholar] [CrossRef] [PubMed]
- Consolandi, C.; Turroni, S.; Emmi, G.; Severgnini, M.; Fiori, J.; Peano, C.; Biagi, E.; Grassi, A.; Rampelli, S.; Silvestri, E.; Centanni, M.; et al. Behçet’s syndrome patients exhibit specific microbiome signature. Autoimmun. Rev. 2015, 14, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Coit, P.; Mumcu, G.; Ture-Ozdemir, F.; Unal, A.U.; Alpar, U.; Bostanci, N.; Ergun, T.; Direskeneli, H.; Sawalha, A.H. Sequencing of 16S rRNA reveals a distinct salivary microbiome signature in Behçet’s disease. Clin. Immunol. 2016, 169, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Guven-Maiorov, E.; Tsai, C.J.; Nussinov, R. Pathogen mimicry of host protein-protein interfaces modulates immunity. Semin. Cell. Dev. Biol. 2016, 58, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Pianta, A.; Arvikar, S.L.; Strle, K.; Drouin, E.E.; Wang, Q.; Costello, C.E.; Steere, A.C. Two rheumatoid arthritis-specific autoantigens correlate microbial immunity with autoimmune responses in joints. J. Clin. Investig. 2017, 127, 2946–2956. [Google Scholar] [CrossRef] [PubMed]
- Shipman, W.D.; Dasoveanu, D.C.; Lu, T.T. Tertiary lymphoid organs in systemic autoimmune diseases: Pathogenic or protective? F1000Research 2017, 6, 196. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Galy, A.; Bruneval, P.; Morvan, M.; Hyafil, F.; Benali, K.; Pasi, N.; Deschamps, L.; Pellenc, Q.; Papo, T.; et al. Tertiary Lymphoid Organs in Takayasu Arteritis. Front. Immunol. 2016, 7, 158. [Google Scholar] [CrossRef] [PubMed]
- Lochner, M.; Ohnmacht, C.; Presley, L.; Bruhns, P.; Si-Tahar, M.; Sawa, S.; Eberl, G. Microbiota-induced tertiary lymphoid tissues aggravate inflammatory disease in the absence of RORgamma t and LTi cells. J. Exp. Med. 2011, 208, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Schwebach, J.R.; Chen, B.; Glatman-Freedman, A.; Casadevall, A.; McKinney, J.D.; Harb, J.L.; McGuire, P.J.; Barkley, W.E.; Bloom, B.R.; Jacobs, W.R. Infection of mice with aerosolized Mycobacterium tuberculosis: Use of a nose-only apparatus for delivery of low doses of inocula and design of an ultrasafe facility. Appl. Environ. Microbiol. 2002, 68, 4646–4649. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.M. Mouse model of tuberculosis. Cold Spring Harb. Perspect. Med. 2014, 5, a018556. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Demographic Data | Ref | |
|---|---|---|
| Predominant gender: | Female (80%) | [11,22,23] |
| Age at onset (years) | 23 (0–65) | [11,22,23] |
| Ethnicity | Rare in western countries (0.4–3 per million people). More common in southeast Asia, Japan, China, India. (40 per million in Japan) | [11,22,23,24] |
| Clinical Findings | * Frequency (%) | |
| Constitutional Manifestations | [19,25,26,27] | |
| Malaise | 29–56 | |
| Fever | 17–32 | |
| Anorexia | 15–34 | |
| Weight loss | 20–25 | |
| Cardiovascular Findings | [19,22,25,26,27,28] | |
| Hypertension | 33–56 | |
| Bruit of carotid arteries | 32–62 | |
| Claudication | 30–80 | |
| Dyspnea | 10–50 | |
| Carotidynia | 10–36 | |
| Central Nervous System | [19,25,26,27,29] | |
| Headache | 50–70 | |
| Dizziness/vertigo | 24–55 | |
| Syncope | 4–19 | |
| Visual disturbances | 15–35 | |
| Stroke | 3–22 | |
| Skin and Musculoskeletal | ||
| Myalgia | 30 | [25] |
| Arthralgia | 28–39 | [19,22,25] |
| Synovitis | 7.7 | [25] |
| Skin rash | 7.8–20 | [19,26] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espinoza, J.L.; Ai, S.; Matsumura, I. New Insights on the Pathogenesis of Takayasu Arteritis: Revisiting the Microbial Theory. Pathogens 2018, 7, 73. https://doi.org/10.3390/pathogens7030073
Espinoza JL, Ai S, Matsumura I. New Insights on the Pathogenesis of Takayasu Arteritis: Revisiting the Microbial Theory. Pathogens. 2018; 7(3):73. https://doi.org/10.3390/pathogens7030073
Chicago/Turabian StyleEspinoza, J. Luis, Suzue Ai, and Itaru Matsumura. 2018. "New Insights on the Pathogenesis of Takayasu Arteritis: Revisiting the Microbial Theory" Pathogens 7, no. 3: 73. https://doi.org/10.3390/pathogens7030073
APA StyleEspinoza, J. L., Ai, S., & Matsumura, I. (2018). New Insights on the Pathogenesis of Takayasu Arteritis: Revisiting the Microbial Theory. Pathogens, 7(3), 73. https://doi.org/10.3390/pathogens7030073