Impact of Mosquito Age and Insecticide Exposure on Susceptibility of Aedes albopictus (Diptera: Culicidae) to Infection with Zika Virus




Abstract
1. Introduction
2. Results
2.1. Virus Titer of Blood Meals, Blood Feeding Rate, Sample Size and Mortality Rates
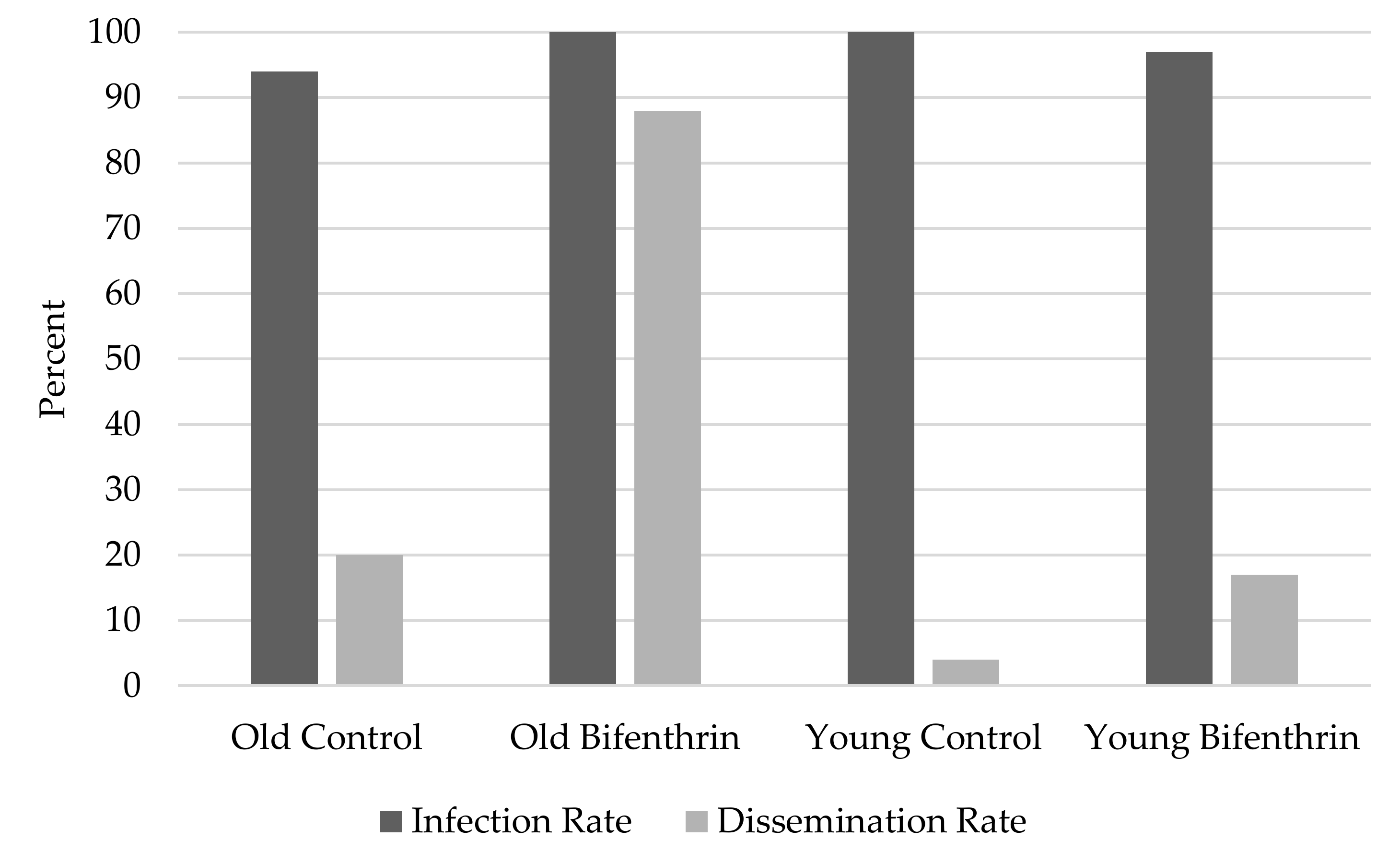
2.2. Infection Rates, Dissemination Rates, Body Virus Titers, and Leg Virus Titers Between Mosquito Ages
3. Discussion
4. Materials and Methods
4.1. Mosquitoes and Virus
4.2. Exposure to Insecticide
4.3. Mosquito Infection
4.4. Blood Meal and Mosquito Processing
4.5. Virus Assays
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weaver, S.C.; Costa, F.; Garcia-Blnco, M.A.; Ko, A.I.; Ribeiro, G.S.; Saade, G.; Shi, P.Y.; Vasilakis, N. Zika virus: History, emergence, biology, and prospects for control. Antivir. Res. 2016, 130, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Available online: www.cdc.gov/zika/vector/range.html (accessed on 5 February 2017).
- Richards, S.L.; Apperson, C.S.; Ponnusamy, L.; Unnasch, T.R. Impacts of host abundance and availability on host preference of Aedes albopictus. Am. J. Trop. Med. Hyg. 2005, 73, 543–551. [Google Scholar]
- Chouin-Carneiro, T.; Vega-Rua, A.; Vazeille, M.; Yebakima, A.; Girod, R.; Goindin, D.; Dupont-Rouzeyrol, M.; Lourenço-de-Oliveira, R.; Failloux, A.B. Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika virus. PLoS Negl. Trop. Dis. 2016, 10, e0004543. [Google Scholar] [CrossRef] [PubMed]
- Dick, G.W. Epidemiological notes on some viruses isolated in Uganda; yellow fever, Rift Valley fever, Bwamba fever, West Nile, Mengo, Semliki Forest, Bunyamwera, Ntaya, Uganda S and Zika viruses. Trans. R. Soc. Trop. Med. Hyg. 1953, 47, 13–48. [Google Scholar] [CrossRef]
- MacNamara, F.N. Zika virus: A report on three cases of human infection during an epidemic of jaundice in Nigeria. Trans. R. Soc. Trop. Med. Hyg. 1954, 48, 139–145. [Google Scholar] [CrossRef]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Guerbois, M.; Fernandez-Salas, I.; Azar, S.R.; Danis-Lozano, R.; Alpuche-Aranda, C.M.; Leal, G.; Garcia-Malo, I.R.; Diaz-Gonzalez, E.E.; Casas-Martinez, M.; Rossi, S.L.; et al. Outbreak of Zika virus infection, Chiapas State, Mexico, 2015, and first confirmed transmission by Aedes aegypti mosquitoes in the Americas. J. Inf. Dis. 2016, 214, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Kindhauser, M.K.; Allen, T.; Frank, V.; Santhana, R.S.; Dye, C. Zika: The origin and spread of a mosquito-borne virus. Bull. World Hlth. Org. 2016, 94, 675. [Google Scholar] [CrossRef] [PubMed]
- Weger-Lucarelli, J.; Rückert, C.; Chotiwan, N.; Nguyen, C.; Luna, S.M.G.; Fauver, J.R.; Foy, B.D.; Perera, R.; Black, W.C.; Kading, R.C.; et al. Vector competence of American mosquitoes for three strains of Zika virus. PLoS Negl. Trop. Dis. 2016, 10, e0005101. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.L. Life table studies of Aedes albopictus (Skuse). In Proceedings of the Sterility Principle for Insect Control or Eradication Proceedings of A Symposium, Athens, Greece, 14–18 September 1970. [Google Scholar]
- National Pesticide Information Center. Available online: www.npic.orst.edu/factsheets/archive/biftech.html (accessed on 18 January 2017).
- Lee, C.Y. Sublethal effects of insecticides on longevity, fecundity and behavior of insect pests: A review. J. Biosci. 2000, 11, 107–112. [Google Scholar]
- Feyereisen, R. Molecular biology of insecticide resistance. Toxicol Lett. 1995, 82, 83–90. [Google Scholar] [CrossRef]
- Moreno-Madriñán, M.J.; Turell, M. Factors of concern regarding Zika and other Aedes aegypti-transmitted viruses in the United States. J. Med. Entomol. 2017, 54, 251–257. [Google Scholar] [PubMed]
- Richards, S.L.; Balanay, J.G.; Fields, M.; Vandock, K. Baseline insecticide susceptibility screening against six active ingredients for Aedes and Culex (Diptera: Culicidae) mosquito populations. J. Med. Entomol. 2017, 54, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.L.; Houk, E.J.; Kramer, L.D.; Reeves, W.C. Intrinsic factors affecting vector competence of mosquitoes for arboviruses. Ann. Rev. Entomol. 1983, 28, 229–262. [Google Scholar] [CrossRef] [PubMed]
- Bennett, K.E.; Olson, K.E.; de Lourdes Muñoz, M.; Fernandez-Salas, I.; Farfan-Ale, J.A.; Higgs, S.; Black, W.C.; Beaty, B.J. Variation in vector competence for dengue 2 virus among 24 collections of Aedes aegypti from Mexico and the United States. Am. J. Trop. Med. Hyg. 2002, 67, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rúa, A.; Zouache, K.; Girod, R.; Failloux, A.B.; Lourenço-de-Oliveira, R. High vector competence of Aedes aegypti and Aedes albopictus from ten American countries as a crucial factor of the spread of chikungunya. J. Virol. 2014, 370, 6294–6306. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.J.; Wei, H.L.; Hsu, E.L.; Chen, E.R. Vector competence of Aedes albopictus and Ae. aegypti (Diptera: Culicidae) to dengue 1 virus on Taiwan: Development of the virus in orally and parenterally infected mosquitoes. J. Med. Entomol. 1993, 30, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Cook, P.E.; McMeniman, C.J.; O’Neill, S.L. Modifying insect population age structure to control vector-borne disease: In transgenesis and the management of vector-borne disease. Adv. Exp. Med. Biol. 2007, 627, 126–140. [Google Scholar]
- Pant, C.P.; Yasuno, M. Field studies on the gonotrophic cycle of Aedes aegypti in Bangkok, Thailand. J. Med. Entomol. 1973, 10, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Boete, C.; Koella, J.C. Evolutionary ideas about genetically manipulated mosquitoes and malaria control. Trends Parasitol. 2003, 19, 32–38. [Google Scholar] [CrossRef]
- Lee, H.L.; Argubano, R.A.; Ahmad, R. Effect of sublethal dosages of malathion on the oral susceptibility of Aedes aegypti to dengue-2 virus infection. SE Asian J. Trop. Med. Public. Hlth. 1997, 28, 664–665. [Google Scholar]
- Rajatileka, S.; Burhani, J.; Ranson, H. Mosquito age and susceptibility to insecticides. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Glunt, K.D.; Thomas, M.B.; Read, A.F. The effects of age, exposure history and malaria infection on the susceptibility of Anopheles mosquitoes to low concentrations of pyrethroid. PLoS ONE 2011, 6, e24968. [Google Scholar] [CrossRef] [PubMed]
- Hodjati, M.H.; Curtis, C.F. Evaluation of the effect of mosquito age and prior exposure to insecticide on pyrethroid tolerance in Anopheles mosquitoes (Diptera: Culicidae). Bull. Entomol. Soc. Am. 1999, 89, 329–337. [Google Scholar] [CrossRef]
- Richards, S.L.; Lord, C.C.; Pesko, K.; Tabachnick, W.J. Environmental and biological factors influencing Culex pipiens quinquefasciatus Say (Diptera: Culicidae) vector competence for Saint Louis encephalitis virus. Am. J. Trop. Med. Hyg. 2009, 81, 264–272. [Google Scholar] [PubMed]
- Richards, S.L.; Lord, C.C.; Pesko, K.N.; Tabachnick, W.J. Environmental and biological factors influencing Culex pipiens quinquefasciatus (Diptera: Culicidae) vector competence for West Nile Virus. Am. J. Trop. Med. Hyg. 2010, 83, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.L.; White, A.V.; Balanay, J.G. Potential for sublethal insecticide exposure to impact vector competence of Aedes albopictus (Diptera: Culicidae) for dengue and Zika viruses. Res. Rep. Trop. Med. 2017, 8, 53–57. [Google Scholar] [CrossRef] [PubMed]
- VanDusen, A.E.; Richards, S.L.; Balanay, J.G. Evaluation of bifenthrin barrier spray on foliage in a suburban eastern North Carolina neighborhood. Pest Manag. Sci. 2016, 72, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. 2013. Available online: https://www.cdc.gov/parasites/education_training/lab/bottlebioassay.html (accessed on 20 February 2018).
- Richards, S.L.; Pesko, K.; Alto, B.W.; Mores, C.N. Reduced infection in mosquitoes exposed to blood meals containing previously frozen flaviviruses. Virus Res. 2007, 129, 224–227. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Group | Sample Size (Day 0) | Sample Size (Day 7) | Mortality Rate (%) |
|---|---|---|---|
| Old Control | 29 | 16 | 45 |
| Old Bifenthrin | 63 | 14 | 78 |
| Young Control | 36 | 24 | 33 |
| Young Bifenthrin | 50 | 30 | 40 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knecht, H.; Richards, S.L.; Balanay, J.A.G.; White, A.V. Impact of Mosquito Age and Insecticide Exposure on Susceptibility of Aedes albopictus (Diptera: Culicidae) to Infection with Zika Virus. Pathogens 2018, 7, 67. https://doi.org/10.3390/pathogens7030067
Knecht H, Richards SL, Balanay JAG, White AV. Impact of Mosquito Age and Insecticide Exposure on Susceptibility of Aedes albopictus (Diptera: Culicidae) to Infection with Zika Virus. Pathogens. 2018; 7(3):67. https://doi.org/10.3390/pathogens7030067
Chicago/Turabian StyleKnecht, Heidi, Stephanie L. Richards, Jo Anne G. Balanay, and Avian V. White. 2018. "Impact of Mosquito Age and Insecticide Exposure on Susceptibility of Aedes albopictus (Diptera: Culicidae) to Infection with Zika Virus" Pathogens 7, no. 3: 67. https://doi.org/10.3390/pathogens7030067
APA StyleKnecht, H., Richards, S. L., Balanay, J. A. G., & White, A. V. (2018). Impact of Mosquito Age and Insecticide Exposure on Susceptibility of Aedes albopictus (Diptera: Culicidae) to Infection with Zika Virus. Pathogens, 7(3), 67. https://doi.org/10.3390/pathogens7030067