
Functional Amyloids in Adhesion of Non-albicans Candida Species

 ,
,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth Conditions
2.2. Cell Lines and Growth Conditions
2.3. Amyloid Peptide Detection
2.4. In Silico Analysis
2.5. Aggregation Assays
2.6. FaDu Monolayer Adhesion Assays
3. Results
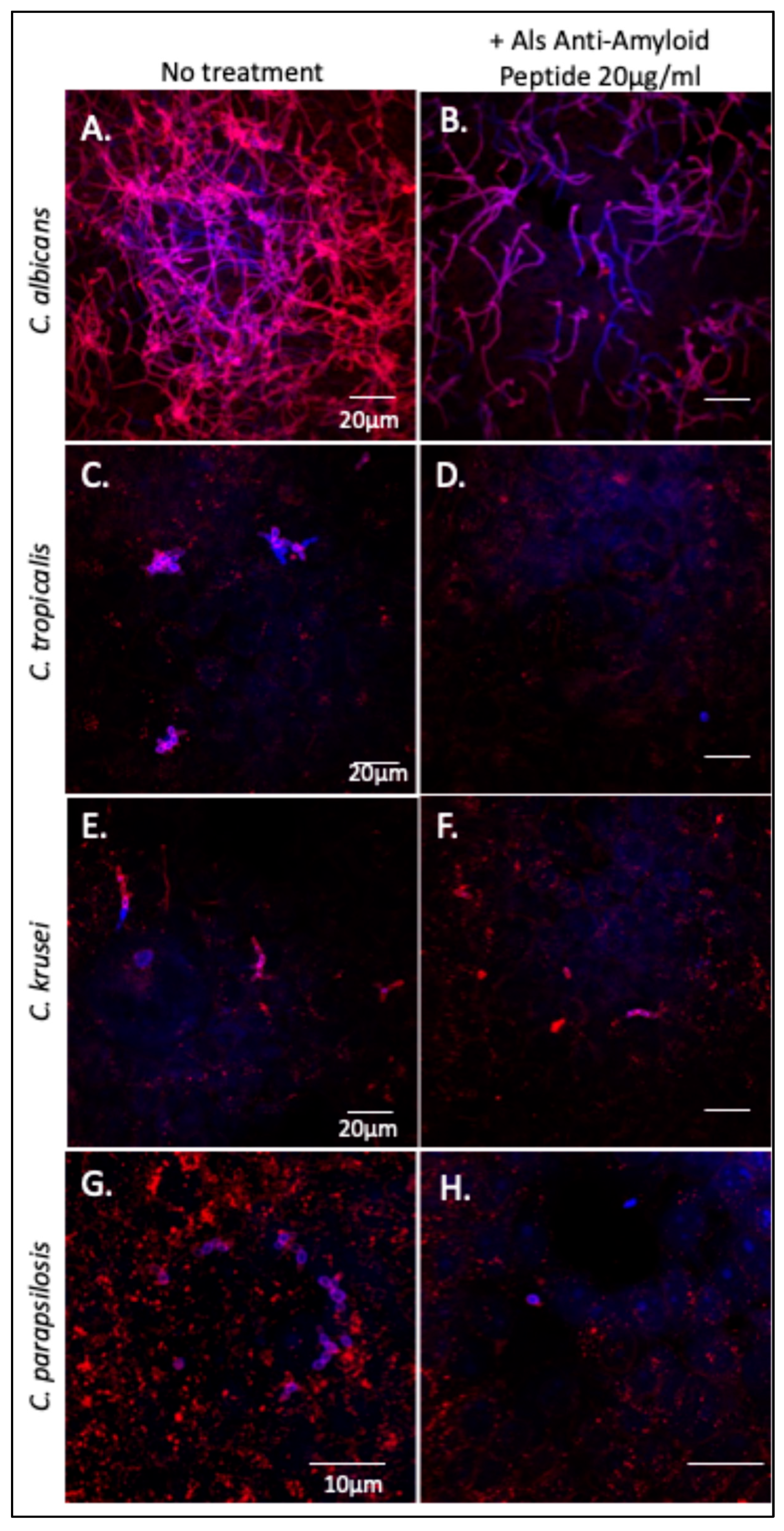
3.1. Als-Amyloid-Binding Peptide Binds to Non-albicans Candida Species
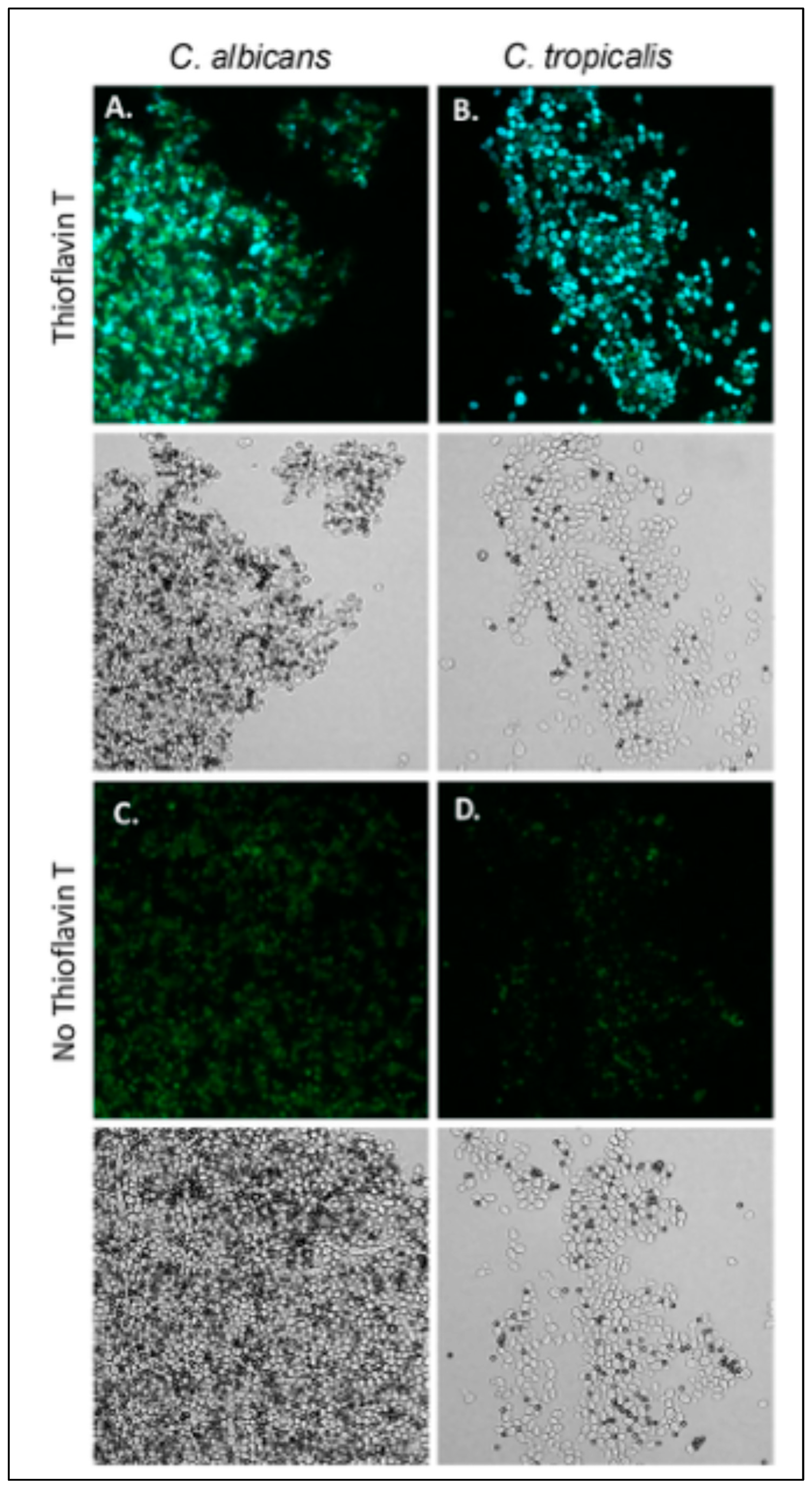
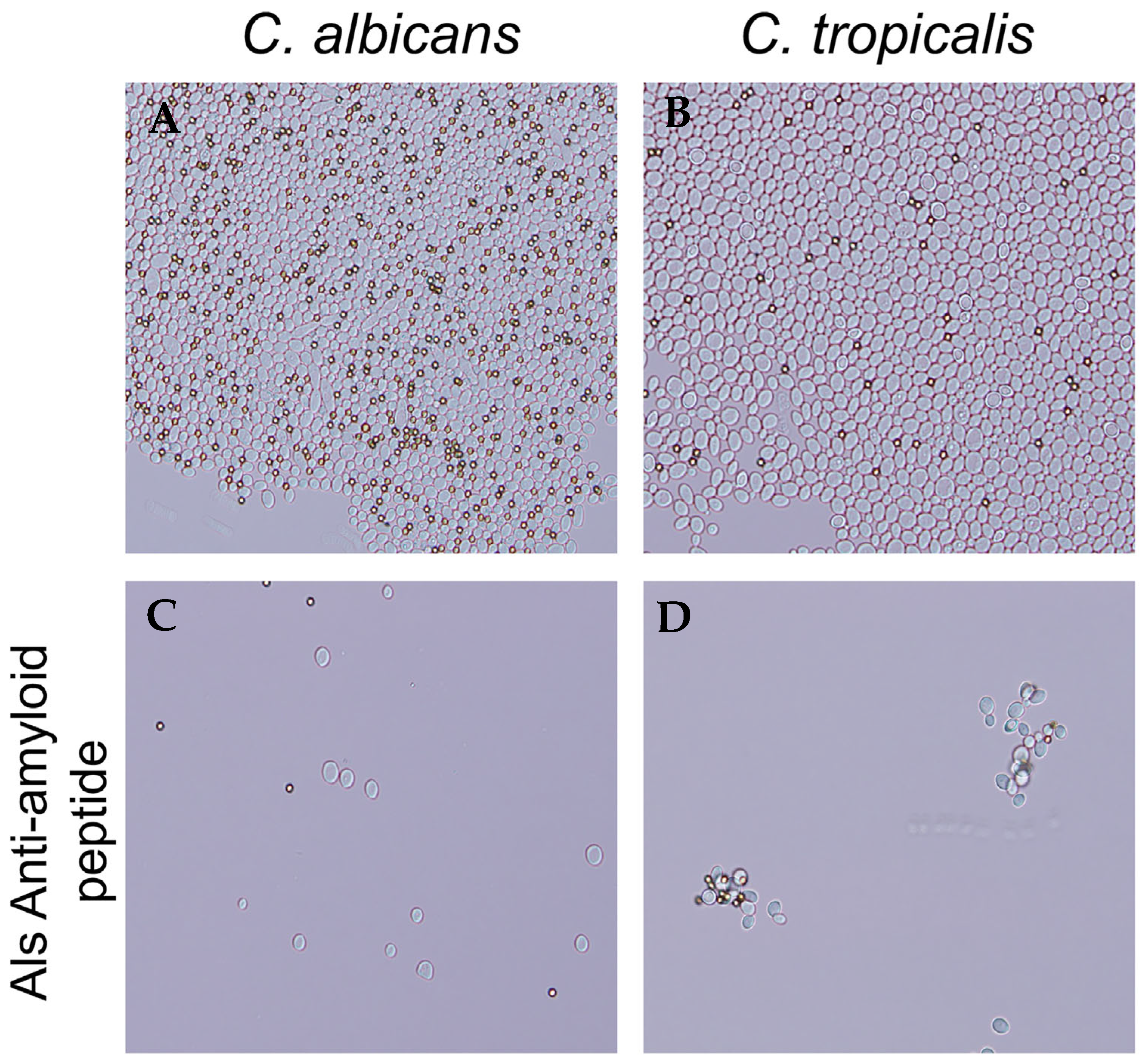
3.2. C. tropicalis Aggregates Are Thioflavin T Positive and Inhibited by the Als-Anti-Amyloid Peptide
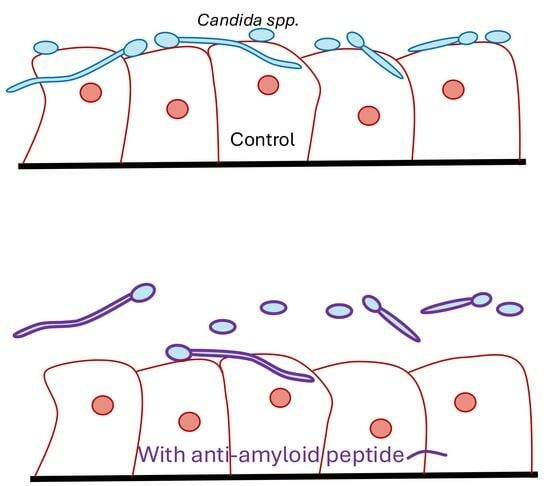
3.3. The Als-Anti-Amyloid Peptide Inhibits Adhesion to Oral Epithelial Cells
4. Discussion
4.1. The Als5-Amyloid-Binding Peptide as a Probe
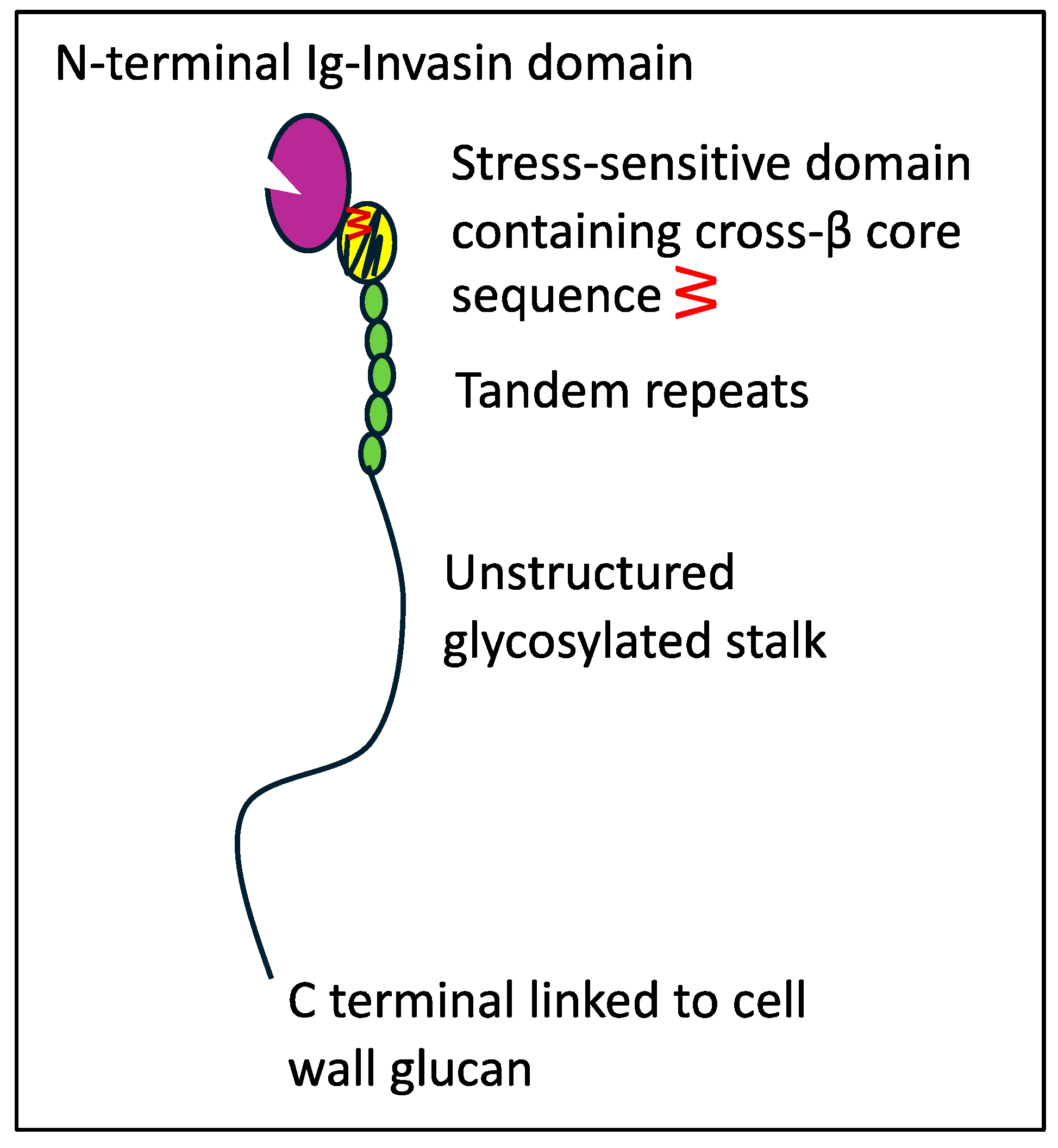
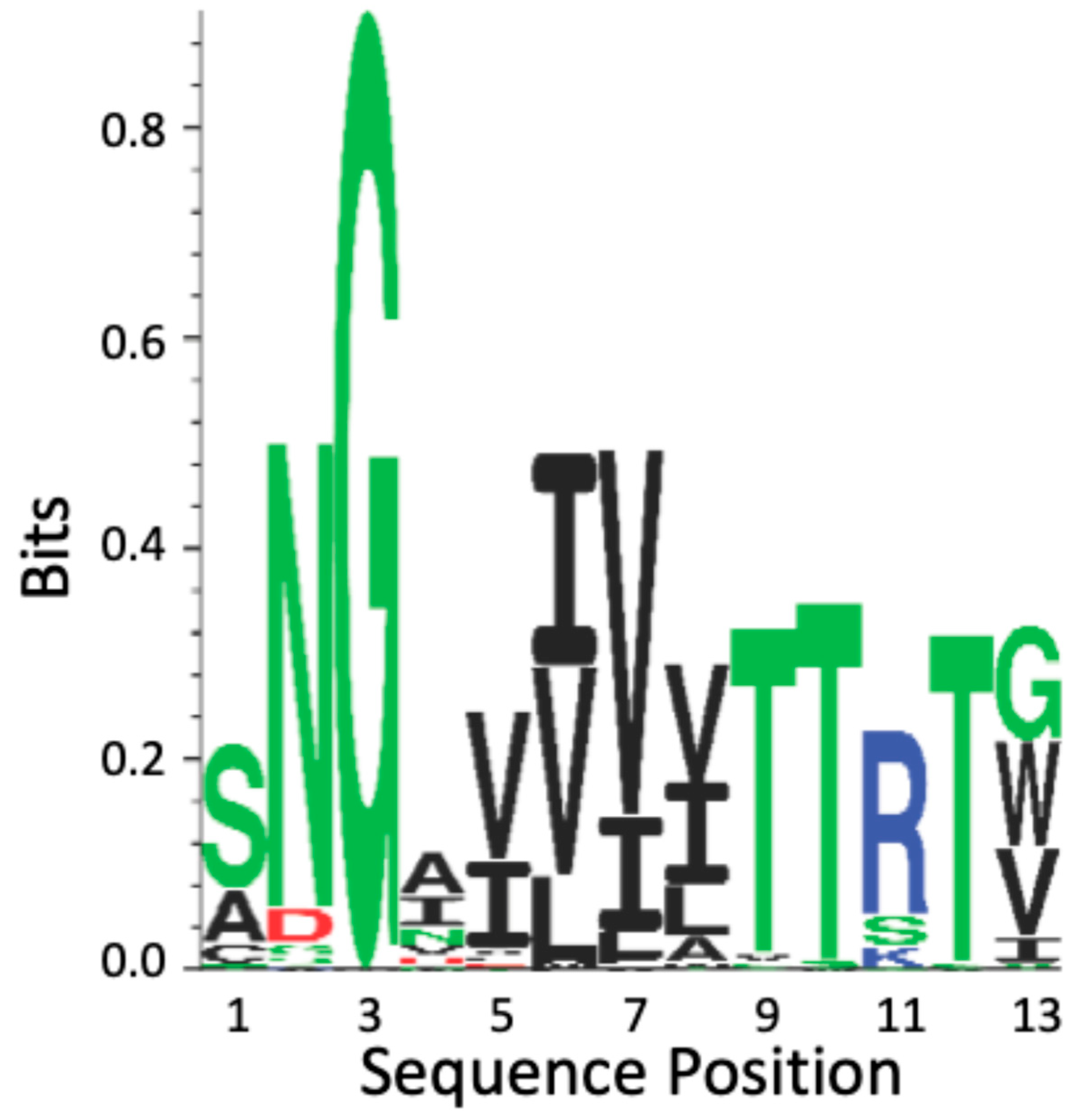
4.2. Als T Region Cross-β Core Sequences in Non-Albicans Candida
4.3. Activity of the Als-Anti-Amyloid Peptide
4.4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lass-Florl, C.; Kanj, S.S.; Govender, N.P.; Thompson, G.R., III; Ostrosky-Zeichner, L.; Govrins, M.A. Invasive candidiasis. Nat. Rev. Dis. Primers 2024, 10, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Galle, F.; Catania, M.R.; Liguori, G. Nosocomial candida infections: Epidemiology of candidaemia. J. Prev. Med. Hyg. 2006, 47, 119–126. [Google Scholar] [PubMed]
- Eggimann, P.; Que, Y.; Revelly, J.; Pagani, J. Preventing invasive Candida infections. Where could we do better? J. Hosp. Infect. 2015, 89, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Trofa, D.; Gacser, A.; Nosanchuk, J.D. Candida parapsilosis, an emerging fungal pathogen. Clin. Microbiol. Rev. 2008, 21, 606–625. [Google Scholar] [CrossRef] [PubMed]
- Lipke, P.N.; Ragonis-Bachar, P. Sticking to the Subject: Multifunctionality in Microbial Adhesins. J. Fungi 2023, 9, 419. [Google Scholar] [CrossRef] [PubMed]
- Lipke, P.N. What We Do Not Know about Fungal Cell Adhesion Molecules. J. Fungi 2018, 4, 59. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.H.; Cheng, G.; Nuessen, J.A.; Jajko, R.; Yeater, K.M.; Zhao, X.; Pujol, C.; Soll, D.R.; Hoyer, L.L. Functional specificity of Candida albicans Als3p proteins and clade specificity of ALS3 alleles discriminated by the number of copies of the tandem repeat sequence in the central domain. Microbiology 2005, 151, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Yeater, K.M.; Chandra, J.; Cheng, G.; Mukherjee, P.K.; Zhao, X.; Rodriguez-Zas, S.L.; Kwast, K.E.; Ghannoum, M.A.; Hoyer, L.L. Temporal analysis of Candida albicans gene expression during biofilm development. Microbiology 2007, 153, 2373–2385. [Google Scholar] [CrossRef] [PubMed]
- Cota, E.; Hoyer, L.L. The Candida albicans agglutinin-like sequence family of adhesins: Functional insights gained from structural analysis. Future Microbiol. 2015, 10, 1635–1648. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Smith, B.; Miller, A.N.; Staker, B.; Fields, C.; Hernandez, A.; Hoyer, L.L. Agglutinin-Like Sequence (ALS) Genes in the Candida parapsilosis Species Complex: Blurring the Boundaries Between Gene Families That Encode Cell-Wall Proteins. Front. Microbiol. 2019, 10, 781. [Google Scholar] [CrossRef] [PubMed]
- Kapteyn, J.C.; Montijn, R.C.; Vink, E.; de la Cruz, J.; Lobell, A.; Douwes, J.E.; Shimoi, H.; Lipke, P.N.; Klis, F.M. Retention of Saccharomyces cerevisiae cell wall proteins through a phosphodiester-linked β1,3/β1,6-glucan heteropoymer. Glycobiology 1996, 6, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Kapteyn, J.C.; Montijn, R.C.; Dijkgraaf, G.J.; Van den Ende, H.; Klis, F.M. Covalent association of β-1,3-glucan with β-1,6-glucosylated mannoproteins in cell walls of Candida albicans. J. Bacteriol. 1995, 177, 3788–3792. [Google Scholar] [CrossRef] [PubMed]
- Kapteyn, J.C.; Hoyer, L.L.; Hecht, J.E.; Muller, W.H.; Andel, A.; Verkleij, A.J.; Makarow, M.; Van Den Ende, H.; Klis, F.M. The cell wall architecture of Candida albicans wild-type cells and cell wall-defective mutants. Mol. Microbiol. 2000, 35, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, L.L.; Cota, E. Candida albicans Agglutinin-Like Sequence (Als) Family Vignettes: A Review of Als Protein Structure and Function. Front. Microbiol. 2016, 7, 280. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Oh, S.H.; Jones, R.; Garnett, J.A.; Salgado, P.S.; Rusnakova, S.; Matthews, S.J.; Hoyer, L.L.; Cota, E. The peptide-binding cavity is essential for Als3-mediated adhesion of Candida albicans to human cells. J. Biol. Chem. 2014, 289, 18401–18412. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D.C.; Yeaman, M.R.; Welch, W.H.; Phan, Q.T.; Fu, Y.; Ibrahim, A.S.; Filler, S.G.; Zhang, M.; Waring, A.J.; Edwards, J.E., Jr. Functional and structural diversity in the Als protein family of Candida albicans. J. Biol. Chem. 2004, 279, 30480–30489. [Google Scholar] [CrossRef] [PubMed]
- Gaur, N.K.; Klotz, S.A.; Henderson, R.L. Overexpression of the Candida albicans ALA1 gene in Saccharomyces cerevisiae results in aggregation following attachment of yeast cells to extracellular matrix proteins, adherence properties similar to those of Candida albicans. Infect. Immun. 1999, 67, 6040–6047. [Google Scholar] [CrossRef] [PubMed]
- Frank, A.T.; Ramsook, C.B.; Otoo, H.N.; Tan, C.; Soybelman, G.; Rauceo, J.M.; Gaur, N.K.; Klotz, S.A.; Lipke, P.N. Structure and function of glycosylated tandem repeats from Candida albicans Als adhesins. Eukaryot. Cell 2010, 9, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Lipke, P.N. Not gently down the stream: Flow induces amyloid bonding in environmental and pathological fungal biofilms. mBio 2025, 16, e00203-25. [Google Scholar] [CrossRef] [PubMed]
- Lipke, P.N.; Klotz, S.A.; Dufrene, Y.F.; Jackson, D.N.; Garcia-Sherman, M.C. Amyloid-Like beta-Aggregates as Force-Sensitive Switches in Fungal Biofilms and Infections. Microbiol. Mol. Biol. Rev. 2017, 82, e00035-17. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.C.; Lee, J.T.; Ramsook, C.B.; Alsteens, D.; Dufrene, Y.F.; Lipke, P.N. A role for amyloid in cell aggregation and biofilm formation. PLoS ONE 2011, 6, e17632. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Isenhower, A.; Rodriguez-Bobadilla, R.; Smith, B.; Jones, J.; Hubka, V.; Fields, C.; Hernandez, A.; Hoyer, L.L. Pursuing Advances in DNA Sequencing Technology to Solve a Complex Genomic Jigsaw Puzzle: The Agglutinin-Like Sequence (ALS) Genes of Candida tropicalis. Front. Microbiol. 2021, 11, 594531. [Google Scholar] [CrossRef] [PubMed]
- Gaur, N.K.; Klotz, S.A. Expression, cloning, and characterization of a Candida albicans gene, ALA1, that confers adherence properties upon Saccharomyces cerevisiae for extracellular matrix proteins. Infect. Immun. 1997, 65, 5289–5294. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sherman, M.C.; Lysak, N.; Filonenko, A.; Richards, H.; Sobonya, R.E.; Klotz, S.A.; Lipke, P.N. Peptide detection of fungal functional amyloids in infected tissue. PLoS ONE 2014, 9, e86067. [Google Scholar] [CrossRef] [PubMed]
- Alsteens, D.; Garcia, M.C.; Lipke, P.N.; Dufrene, Y.F. Force-induced formation and propagation of adhesion nanodomains in living fungal cells. Proc. Natl. Acad. Sci. USA 2010, 107, 20744–20749. [Google Scholar] [CrossRef] [PubMed]
- Ho, V.; Herman-Bausier, P.; Shaw, C.; Conrad, K.A.; Garcia-Sherman, M.C.; Draghi, J.; Dufrene, Y.F.; Lipke, P.N.; Rauceo, J.M. An Amyloid Core Sequence in the Major Candida albicans Adhesin Als1p Mediates Cell-Cell Adhesion. mBio 2019, 10, e01766-19. [Google Scholar] [CrossRef] [PubMed]
- Mourer, T.; El Ghalid, M.; Pehau-Arnaudet, G.; Kauffmann, B.; Loquet, A.; Brûlé, S.; Cabral, V.; D’enfert, C.; Bachellier-Bassi, S. The Pga59 cell wall protein is an amyloid forming protein involved in adhesion and biofilm establishment in the pathogenic yeast Candida albicans. Npj Biofilms Microbiomes 2023, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.X.; El-Kirat-Chatel, S.; Joseph, I.G.; Jackson, D.N.; Ramsook, C.B.; Dufrene, Y.F.; Lipke, P.N. Force Sensitivity in Saccharomyces cerevisiae Flocculins. mSphere 2016, 1, e00128-16. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.X.; Lipke, P.N. Role of force-sensitive amyloid-like interactions in fungal catch bonding and biofilms. Eukaryot. Cell 2014, 13, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Golan, N.; Schwartz-Perov, S.; Landau, M.; Lipke, P.N. Structure and Conservation of Amyloid Spines from the Candida albicans Als5 Adhesin. Front. Mol. Biosci. 2022, 9, 926959. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Sherman, M.C.; Lundberg, T.; Sobonya, R.E.; Lipke, P.N.; Klotz, S.A. A unique biofilm in human deep mycoses: Fungal amyloid is bound by host serum amyloid P component. NPJ Biofilms Microbiomes 2015, 1, 15009. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, K.B.; Garcia, M.C.; Sobonya, R.; Lipke, P.N.; Klotz, S.A. New Features of Invasive Candidiasis in Humans: Amyloid Formation by Fungi and Deposition of Serum Amyloid P Component by the Host. J. Infect. Dis. 2012, 206, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Thorn, J.L.; Gilchrist, K.B.; Sobonya, R.E.; Gaur, N.K.; Lipke, P.N.; Klotz, S.A. Postmortem candidaemia: Marker of disseminated disease. J. Clin. Pathol. 2010, 63, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Golconda, U.; Sobonya, R.E.; Klotz, S.A. Do Pentraxins Bind to Fungi in Invasive Human Gastrointestinal Candidiasis? J. Fungi 2018, 4, 111. [Google Scholar] [CrossRef] [PubMed]
- Klotz, S.A.; Sobonya, R.E.; Lipke, P.N.; Garcia-Sherman, M.C. Serum Amyloid P Component and Systemic Fungal Infection: Does It Protect the Host or Is It a Trojan Horse? Open Forum Infect. Dis. 2016, 3, ofw166. [Google Scholar] [CrossRef] [PubMed]
- Pepys, M.B. Invasive Candidiasis: New Insights Presaging New Therapeutic Approaches? J. Infect. Dis. 2012, 206, 1339–1341. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.F.; Finan, C.; Chopade, S.; Ellmerich, S.; Rossor, M.N.; Hingorani, A.D.; Pepys, M.B. Genetic evidence for serum amyloid P component as a drug target in neurodegenerative disorders. Open Biol. 2024, 14, 230419. [Google Scholar] [CrossRef] [PubMed]
- Richards, D.B.; Cookson, L.M.; Berges, A.C.; Barton, S.V.; Lane, T.; Ritter, J.M.; Fontana, M.; Moon, J.C.; Pinzani, M.; Gillmore, J.D.; et al. Therapeutic Clearance of Amyloid by Antibodies to Serum Amyloid P Component. N. Engl. J. Med. 2015, 373, 1106–1114. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, L.L.; Fundyga, R.; Hecht, J.E.; Kapteyn, J.C.; Klis, F.M.; Arnold, J. Characterization of agglutinin-like sequence genes from non-albicans Candida and phylogenetic analysis of the ALS family. Genetics 2001, 157, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.H.; Schliep, K.; Isenhower, A.; Rodriguez-Bobadilla, R.; Vuong, V.M.; Fields, C.J.; Hernandez, A.G.; Hoyer, L.L. Using Genomics to Shape the Definition of the Agglutinin-Like Sequence (ALS) Family in the Saccharomycetales. Front. Cell. Infect. Microbiol. 2021, 11, 794529. [Google Scholar] [CrossRef] [PubMed]
- Satala, D.; Karkowska-Kuleta, J.; Bras, G.; Rapala-Kozik, M.; Kozik, A. Candida parapsilosis cell wall proteins-CPAR2_404800 and CPAR2_404780-Are adhesins that bind to human epithelial and endothelial cells and extracellular matrix proteins. Yeast 2023, 40, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Kozik, A.; Karkowska-Kuleta, J.; Zajac, D.; Bochenska, O.; Kedracka-Krok, S.; Jankowska, U.; Rapala-Kozik, M. Fibronectin-, vitronectin- and laminin-binding proteins at the cell walls of Candida parapsilosis and Candida tropicalis pathogenic yeasts. BMC Microbiol. 2015, 15, 197. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Li, W.; Liu, X.; Che, J.; Wu, Y.; Lu, J. Distinct Expression Levels of ALS, LIP, and SAP Genes in Candida tropicalis with Diverse Virulent Activities. Front. Microbiol. 2016, 7, 1175. [Google Scholar] [CrossRef] [PubMed]
- Neale, M.N.; Glass, K.A.; Longley, S.J.; Kim, D.J.; Laforce-Nesbitt, S.S.; Wortzel, J.D.; Shaw, S.K.; Bliss, J.M. Role of the inducible adhesin, CpAls7, in binding of Candida parapsilosis to extracellular matrix under fluid shear. Infect. Immun. 2018, 86, e00892-17. [Google Scholar] [CrossRef] [PubMed]
- Zoppo, M.; Di Luca, M.; Franco, M.; Rizzato, C.; Lupetti, A.; Stringaro, A.; De Bernardis, F.; Schaudinn, C.; Barrasa, M.I.; Bottai, D.; et al. CpALS4770 and CpALS4780 contribution to the virulence of Candida parapsilosis. Microbiol. Res. 2020, 231, 126351. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, L.; Zoppo, M.; Rizzato, C.; Bottai, D.; Hernandez, A.G.; Hoyer, L.L.; Tavanti, A. Characterization of the Candida orthopsilosis agglutinin-like sequence (ALS) genes. PLoS ONE 2019, 14, e0215912. [Google Scholar] [CrossRef] [PubMed]
- Bertini, A.; Zoppo, M.; Lombardi, L.; Rizzato, C.; De Carolis, E.; Vella, A.; Torelli, R.; Sanguinetti, M.; Tavanti, A. Targeted gene disruption in Candida parapsilosis demonstrates a role for CPAR2_404800 in adhesion to a biotic surface and in a murine model of ascending urinary tract infection. Virulence 2016, 7, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Galan-Ladero, M.A.; Blanco-Blanco, M.T.; Fernandez-Calderon, M.C.; Lucio, L.; Gutierrez-Martin, Y.; Blanco, M.T.; Perez-Giraldo, C. Candida tropicalis biofilm formation and expression levels of the CTRG ALS-like genes in sessile cells. Yeast 2019, 36, 107–115. [Google Scholar] [CrossRef] [PubMed]
- de Souza, C.M.; Dos Santos, M.M.; Furlaneto-Maia, L.; Furlaneto, M.C. Adhesion and biofilm formation by the opportunistic pathogen Candida tropicalis: What do we know? Can. J. Microbiol. 2023, 69, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Schikora-Tamarit, M.A.; Gabaldon, T. Recent gene selection and drug resistance underscore clinical adaptation across Candida species. Nat. Microbiol. 2024, 9, 284–307. [Google Scholar] [CrossRef] [PubMed]
- Karkowska-Kuleta, J.; Zajac, D.; Bras, G.; Bochenska, O.; Rapala-Kozik, M.; Kozik, A. Binding of human plasminogen and high-molecular-mass kininogen by cell surface-exposed proteins of Candida parapsilosis. Acta Biochim. Pol. 2017, 64, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Escamilla, A.M.; Rousseau, F.; Schymkowitz, J.; Serrano, L. Prediction of sequence-dependent and mutational effects on the aggregation of peptides and proteins. Nat. Biotechnol. 2004, 22, 1302–1306. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Myers, C.L.; Sheppard, D.C.; Phan, Q.T.; Sanchez, A.A.; E Edwards, J.; Filler, S.G. Role of the fungal Ras-protein kinase A pathway in governing epithelial cell interactions during oropharyngeal candidiasis. Cell Microbiol. 2005, 7, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Klotz, S.A.; Gaur, N.K.; De Armond, R.; Sheppard, D.; Khardori, N.; Edwards, J.E.; Lipke, P.; El-Azizi, M. Candida albicans Als proteins mediate aggregation with bacteria and yeasts. Med. Mycol. Off. Publ. Int. Soc. Hum. Anim. Mycol. 2007, 45, 363–370. [Google Scholar] [CrossRef]
- Ramsook, C.B.; Tan, C.; Garcia, M.C.; Fung, R.; Soybelman, G.; Henry, R.; Litewka, A.; O’Meally, S.; Otoo, H.N.; Khalaf, R.A.; et al. Yeast cell adhesion molecules have functional amyloid-forming sequences. Eukaryot. Cell 2010, 9, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Nunez-Beltran, A.; Lopez-Romero, E.; Cuellar-Cruz, M. Identification of proteins involved in the adhesion of Candida species to different medical devices. Microb. Pathog. 2017, 107, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Otoo, H.N.; Lee, K.G.; Qiu, W.; Lipke, P.N. Candida albicans Als adhesins have conserved amyloid-forming sequences. Eukaryot. Cell 2008, 7, 776–782. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Sherman, M.C.; Hamid, S.A.; Jackson, D.N.; Thomas, J.; Lipke, P.N. Functional Amyloids in Adhesion of Non-albicans Candida Species. Pathogens 2025, 14, 723. https://doi.org/10.3390/pathogens14080723
Garcia-Sherman MC, Hamid SA, Jackson DN, Thomas J, Lipke PN. Functional Amyloids in Adhesion of Non-albicans Candida Species. Pathogens. 2025; 14(8):723. https://doi.org/10.3390/pathogens14080723
Chicago/Turabian StyleGarcia-Sherman, Melissa C., Safraz A. Hamid, Desmond N. Jackson, James Thomas, and Peter N. Lipke. 2025. "Functional Amyloids in Adhesion of Non-albicans Candida Species" Pathogens 14, no. 8: 723. https://doi.org/10.3390/pathogens14080723
APA StyleGarcia-Sherman, M. C., Hamid, S. A., Jackson, D. N., Thomas, J., & Lipke, P. N. (2025). Functional Amyloids in Adhesion of Non-albicans Candida Species. Pathogens, 14(8), 723. https://doi.org/10.3390/pathogens14080723