
Cloning and Expression of a Truncated Form of the p72 Protein of the African Swine Fever Virus (ASFV) for Application in an Efficient Indirect ELISA System


 ,
,  , , ,
, , ,  ,
,  and
and
Abstract

1. Introduction
2. Materials and Methods
2.1. Reference Swine Serum Samples
2.2. In Silico and Bioinformatics Analysis of ASF-p72-Truncated Protein
2.3. Cloning, Subcloning, and Overexpression of ASF-p72-Truncated Protein
2.4. Antigenic Evaluation of ASF-p72-Truncated Protein in a Mouse Model
2.5. Standardization of Indirect ELISA (iELISA) of ASF-p72-Truncated Protein with Pig Serum Samples
2.6. Statistical Analysis (Sensitivity, Specificity, and Index Kappa) of iELISA
2.7. Determination of the iELISA Cut-Off Point
3. Results
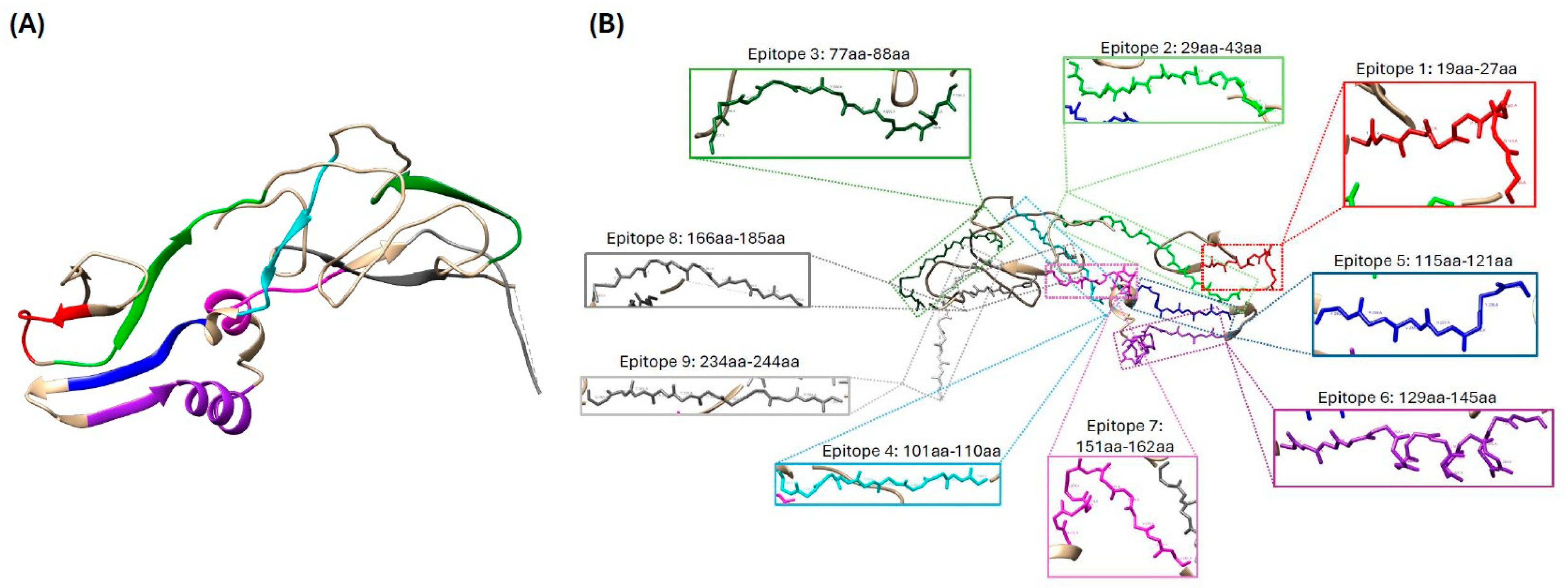
3.1. In Silico Analysis of ASF-p72-Truncated Protein
3.2. Bioinformatics Analysis of ASF-p72-Truncated Protein
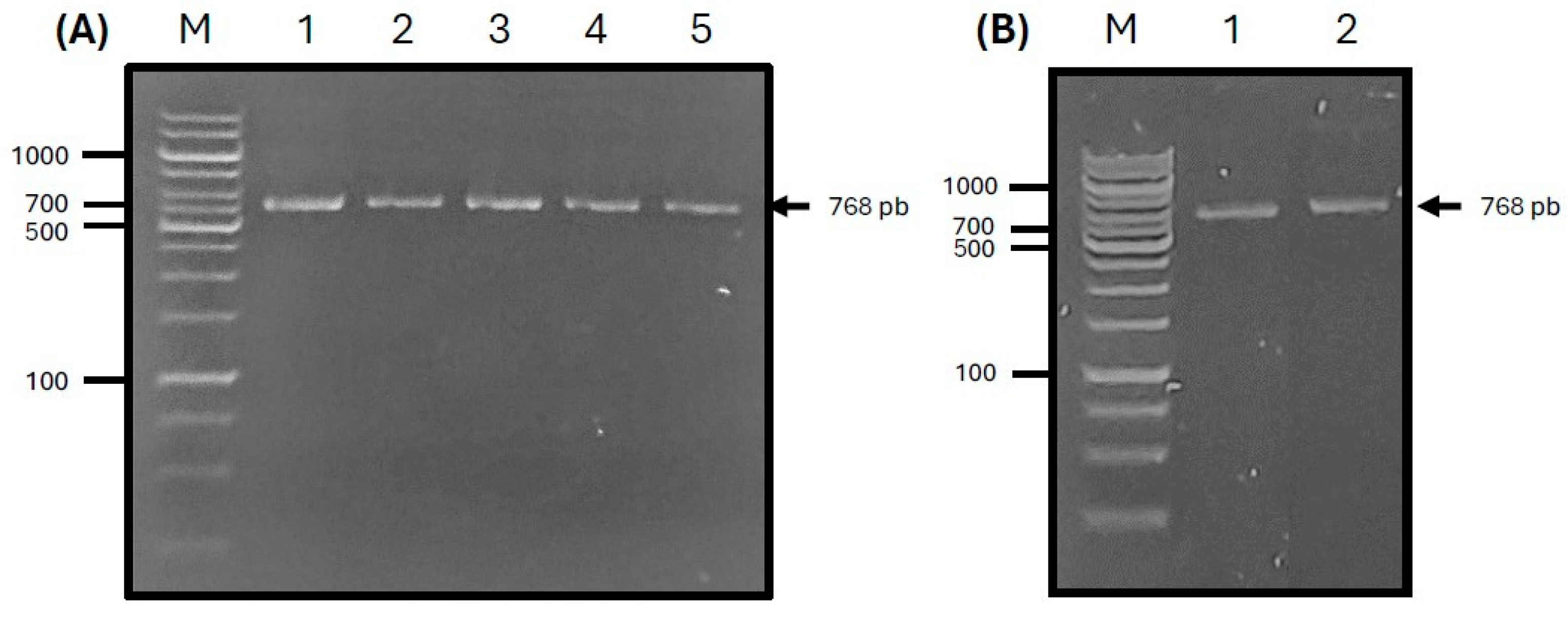
3.3. Cloning and Expression of ASF-p72-Truncated Protein
3.4. Antigenicity of ASF-p72-Truncated Protein by Mice Immunization
3.5. Standardization and Validation of Indirect ELISA (iELISA) of ASF-p72-Truncated Protein
3.6. Determination of Sensitivity, Specificity, and Concordance of iELISA
3.7. Determination of Cut-Off Values
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASF | African swine fever |
| ASFV | African swine fever virus |
| CReSA | Centre de Recerca en Sanitat Animal |
| CV | coefficient of variation |
| DP | double positive |
| ELISA | enzyme-linked immunosorbent assay |
| EURL | European Union Reference Laboratory |
| HRP | horseradish peroxidase |
| IB | immunoblotting |
| iELISA | indirect enzyme-linked immunosorbent assay |
| IIF | indirect immunofluorescence |
| IMAC | immobilized metal affinity chromatography |
| IPT | indirect immunoperoxidase test |
| IPTG | isopropyl β-D-1-thiogalactopyranoside |
| IRTA | Institute for Research and Technology in Food and Agriculture |
| MAb | monoclonal antibody |
| OIE | Organization for Animal Health |
| ORF | open reading frame |
| SC | subcutaneous |
| SD | standard deviations |
| TMB | 3,3′,5,5′-tetramethylbenzidine |
| κ | kappa |
References
- Sánchez-Cordón, P.J.; Montoya, M.; Reis, A.L.; Dixon, L.K. African swine fever: A re-emerging viral disease threatening the global pig industry. Vet. J. 2018, 233, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, M.C.; Reoyo, A.T.; Fernández-Pinero, J.; Iglesias, I.; Muñoz, M.J.; Arias, M.L. African swine fever: A global view of the current challenge. Porcine Health Manag. 2015, 1, 21. [Google Scholar] [CrossRef]
- Ge, S.; Li, J.; Fan, X.; Liu, F.; Li, L.; Wang, Q.; Ren, W.; Bao, J.; Liu, C.; Wang, H.; et al. Molecular characterization of African swine fever virus, China, 2018. Emerg. Infect. Dis. 2018, 24, 2131–2133. [Google Scholar] [CrossRef] [PubMed]
- Global African Swine Fever Research Alliance. Available online: https://www.ars.usda.gov/GARA/reports.htm (accessed on 21 April 2025).
- Gonzales, W.; Moreno, C.; Duran, U.; Henao, N.; Bencosme, M.; Lora, P.; Reyes, R.; Núñez, R.; De Gracia, A.; Perez, A.M. African swine fever in the Dominican Republic. Transbound. Emerg. Dis. 2021, 68, 3018–3019. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.K.; Escribano, J.M.; Martins, C.; Rock, D.L.; Salas, M.L.; Wilkinson, P.J. Asfarviridae. In Virus Taxonomy, VIIIth Report of the ICTV; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Elsevier/Academic Press: London, UK, 2005; pp. 135–143. [Google Scholar]
- Anderson, E.C.; Hutchings, G.H.; Mukarati, N.; Wilkinson, P.J. African swine fever virus infection of the bushpig (Potamochoerus porcus) and its significance in the epidemiology of the disease. Vet. Microbiol. 1998, 62, 1–15. [Google Scholar] [CrossRef]
- Kleiboeker, S.B.; Scoles, G.A.; Burrage, T.G.; Sur, J. African swine fever virus replication in the midgut epithelium is required for infection of ornithodoros ticks. J. Virol. 1999, 73, 8587–8598. [Google Scholar] [CrossRef]
- Duan, X.; Ru, Y.; Yang, W.; Ren, J.; Hao, R.; Qin, X.; Li, D.; Zheng, H. Research progress on the proteins involved in african swine fever virus infection and replication. Front. Immunol. 2022, 13, 947180. [Google Scholar] [CrossRef]
- Alejo, A.; Matamoros, T.; Guerra, M.; Andres, G. A proteomic atlas of the african swine fever virus particle. J. Virol. 2018, 92, e01293-18. [Google Scholar] [CrossRef]
- Spinard, E.; Dinhobl, M.; Tesler, N.; Birtley, H.; Signore, A.V.; Ambagala, A.; Masembe, C.; Borca, M.V.; Gladue, D.P. A Re-Evaluation of African Swine Fever Genotypes Based on P72 Sequences Reveals the Existence of Only Six Distinct P72 Groups. Viruses 2023, 15, 2246. [Google Scholar] [CrossRef]
- Wang, Y.; Kang, W.; Yang, W.; Zhang, J.; Li, D.; Zheng, H. Structure of African Swine Fever Virus and Associated Molecular Mechanisms Underlying Infection and Immunosuppression: A Review. Front. Immunol. 2021, 12, 715582. [Google Scholar] [CrossRef]
- Liu, Q.; Ma, B.; Qian, N.; Zhang, F.; Tan, X.; Lei, J.; Xiang, Y. Structure of the African swine fever virus major capsid protein p72. Cell Res. 2019, 29, 953–955. [Google Scholar] [CrossRef]
- Oura, C.A.L.; Denyer, M.S.; Takamatsu, H.; Parkhouse, R.M.E. In vivo depletion of CD8+ T lymphocytes abrogates protective immunity to African swine fever virus. J. Gen. Virol. 2005, 86, 2445–2450. [Google Scholar] [CrossRef]
- Montoya, M.; Franzoni, G.; Pérez-Nuñez, D.; Revilla, Y.; Galindo, I.; Alonso, C.; Netherton, C.L.; Blohm, U. 3. Immune responses against African swine fever virus infection. In Understanding and Combatting African Swine Fever; Brill: Leiden, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Gladue, D.P.; Borca, M.V. Recombinant ASF Live Attenuated Virus Strains as Experimental Vaccine Candidates. Viruses 2022, 14, 878. [Google Scholar] [CrossRef]
- Revilla, Y.; Pérez-Núñez, D.; Richt, J.A. African Swine Fever Virus Biology and Vaccine Approaches. Adv. Virus Res. 2018, 100, 41–74. [Google Scholar] [CrossRef] [PubMed]
- Oura, C.A.; Edwards, L.; Batten, C.A. Virological diagnosis of African swine fever--comparative study of available tests. Virus Res. 2013, 173, 150–158. [Google Scholar] [CrossRef] [PubMed]
- VALIDATION GUIDELINE 3.6.2.: Development and Optimisation of Antigen Detection Assays. Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/aahm/current/GUIDELINE_3.6.2_ANTIGEN_DETECT.pdf (accessed on 21 April 2025).
- Muzykina, L.; Barrado-Gil, L.; Gonzalez-Bulnes, A.; Crespo-Piazuelo, D.; Cerón, J.J.; Alonso, C.; Montoya, M. Overview of Modern Commercial Kits for Laboratory Diagnosis of African Swine Fever and Swine Influenza A Viruses. Viruses 2024, 16, 505. [Google Scholar] [CrossRef] [PubMed]
- African Swine Fever (Infection with African Swine Fever Virus). Available online: https://www.gob.mx/cms/uploads/attachment/file/870460/e-bsc-23_anexo16.pdf (accessed on 21 April 2025).
- Onisk, D.V.; Borca, M.V.; Kutish, G.; Kramer, E.; Irusta, P.; Rock, D.L. Passively transferred African swine fever virus antibodies protect swine against lethal infection. Virology 1994, 198, 350–354. [Google Scholar] [CrossRef]
- Jameson, B.A.; Wolf, H. The antigenic index: A novel algorithm for predicting antigenic determinants. Comput. Appl. Biosci. CABIOS 1988, 4, 181–186. [Google Scholar] [CrossRef]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Emini, E.A.; Hughes, J.V.; Perlow, D.S.; Boger, J. Induction of hepatitis A virus neutralizing antibody by a virus-specific synthetic peptide. J. Virol. 1985, 55, 836–839. [Google Scholar] [CrossRef]
- Gupta, R.; Brunak, S. Prediction of glycosylation across the human proteome and the correlation to protein function. Pac. Symp. Biocomput. 2001, 310–322. [Google Scholar] [CrossRef]
- Lara-Romero, R.; Cerriteño-Sánchez, J.L.; Mendoza-Elvira, S.; García-Cambrón, J.B.; Castañeda-Montes, M.A.; Pérez-Aguilar, J.M.; Cuevas-Romero, J.S. Development of Novel Recombinant Antigens of Nucleoprotein and Matrix Proteins of Porcine orthorubulavirus: Antigenicity and Structural Prediction. Viruses 2022, 14, 1946. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Montes, M.A.; Cuevas-Romero, J.S.; Cerriteño-Sánchez, J.L.; de María Ávila-De la Vega, L.; García-Cambrón, J.B.; Ramírez-Álvarez, H. Small ruminant lentivirus capsid protein (SRLV-p25) antigenic structural prediction and immunogenicity to recombinant SRLV-rp25-coupled to immunostimulatory complexes based on glycyrrhizinic acid. Biosci. Biotechnol. Biochem. 2023, 87, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-Romero, J.S.; Rivera-Benítez, J.F.; Hernández-Baumgarten, E.; Hernández-Jaúregui, P.; Vega, M.; Blomström, A.L.; Berg, M.; Baule, C. Cloning, expression and characterization of potential immunogenic recombinant hemagglutinin-neuraminidase protein of Porcine rubulavirus. Protein Expr. Purif. 2016, 128, 1–7. [Google Scholar] [CrossRef]
- Norma Oficial Mexicana NOM-062-ZOO-1999. Especificaciones Técnicas Para la Producción, Cuidado y Uso de Los Animales de Laboratorio. Available online: https://www.gob.mx/cms/uploads/attachment/file/203498/NOM-062-ZOO-1999_220801.pdf (accessed on 21 April 2025).
- Crowther, J. Enzyme Linked Immunosorbent Assay (ELISA). In Molecular Biomethods Handbook. Springer Protocols Handbooks; Walker, J.M., Rapley, R., Eds.; Humana Press: Totowa, NJ, USA, 2008. [Google Scholar] [CrossRef]
- Calculating Inter- and Intra-Assay Coefficients of Variability. Available online: https://salimetrics.com/calculating-inter-and-intra-assay-coefficients-of-variability/ (accessed on 21 April 2025).
- Tighe, P.J.; Ryder, R.R.; Todd, I.; Fairclough, L.C. ELISA in the multiplex era: Potentials and pitfalls. Proteomics. Clin. Appl. 2015, 9, 406–422. [Google Scholar] [CrossRef]
- Abraira, V. El Índice Kappa. Medicina de Familia. Semergen 2001, 27, 247–249. [Google Scholar] [CrossRef]
- Dierksheide, C.W. Medical decisions: Interpreting clinical test; in clinical practice, the predictive value helps reduce uncertainties when tests are used in diagnosis. ASM News 1987, 53, 677–680. [Google Scholar]
- Power, M.; Fell, G.; Wright, M. Principles for high-quality, high-value testing. Evid.-Based Med. 2013, 18, 5–10. [Google Scholar] [CrossRef]
- López-Ratón, M.; Rodríguez-Álvarez, M.X.; Cadarso-Suárez, C.; Gude-Sampedro, F. Optimal Cutpoints: An R Package for Selecting Optimal Cutpoints in Diagnostic Tests. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar] [CrossRef]
- Spinard, E.; O’Donnell, V.; Vuono, E.; Rai, A.; Davis, C.; Ramirez-Medina, E.; Espinoza, N.; Valladares, A.; Borca, M.V.; Gladue, D.P. Full genome sequence for the African swine fever virus outbreak in the Dominican Republic in 1980. Sci. Rep. 2023, 13, 1024. [Google Scholar] [CrossRef]
- Gallardo, C.; Fernández-Pinero, J.; Pelayo, V.; Gazaev, I.; Markowska-Daniel, I.; Pridotkas, G.; Nieto, R.; Fernández-Pacheco, P.; Bokhan, S.; Nevolko, O.; et al. Genetic variation among African swine fever genotype II viruses, eastern and central Europe. Emerg. Infect. Dis. 2014, 20, 1544–1547. [Google Scholar] [CrossRef]
- Qu, H.; Ge, S.; Zhang, Y.; Wu, X.; Wang, Z. A systematic review of genotypes and serogroups of African swine fever virus. Virus Genes 2022, 58, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Yang, S.; Shao, J.; Zhou, G.; Ma, Y.; Wen, S.; Hou, Z.; Peng, D.; Guo, H.; Liu, W.; et al. Identification of p72 epitopes of African swine fever virus and preliminary application. Front. Microbiol. 2023, 14, 1126794. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhao, D.; Wang, J.; Zhang, Y.; Wang, M.; Gao, Y.; Li, F.; Wang, J.; Bu, Z.; Rao, Z.; et al. Architecture of African swine fever virus and implications for viral assembly. Science 2019, 366, 640–644. [Google Scholar] [CrossRef] [PubMed]
- OIE Terrestrial Manual. Chapter, 2019, 3(8):1. Available online: https://www.woah.org/fileadmin/Home/fr/Health_standards/tahm/3.08.01_ASF.pdf (accessed on 21 April 2025).
- Gallardo, C.; Soler, A.; Nieto, R.; Carrascosa, A.L.; De Mia, G.M.; Bishop, R.P.; Martins, C.; Fasina, F.O.; Couacy-Hymman, E.; Heath, L.; et al. Comparative evaluation of novel African swine fever virus (ASF) antibody detection techniques derived from specific ASF viral genotypes with the OIE internationally prescribed serological tests. Vet. Microbiol. 2013, 162, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jiao, J.; Liu, N.; Ren, S.; Zeng, H.; Peng, J.; Zhang, Y.; Guo, L.; Liu, F.; Lv, T.; et al. Novel p22 and p30 dual-proteins combination based indirect ELISA for detecting antibodies against African swine fever virus. Front. Vet. Sci. 2023, 10, 1093440. [Google Scholar] [CrossRef]
- Zhou, L.; Song, J.; Wang, M.; Sun, Z.; Sun, J.; Tian, P.; Zhuang, G.; Zhang, A.; Wu, Y.; Zhang, G. Establishment of a Dual-Antigen Indirect ELISA Based on p30 and pB602L to Detect Antibodies against African Swine Fever Virus. Viruses 2023, 15, 1845. [Google Scholar] [CrossRef]
- Gallardo, C.; Nieto, R.; Soler, A.; Pelayo, V.; Fernández-Pinero, J.; Markowska-Daniel, I.; Pridotkas, G.; Nurmoja, I.; Granta, R.; Simón, A.; et al. Assessment of African Swine Fever Diagnostic Techniques as a Response to the Epidemic Outbreaks in Eastern European Union Countries: How To Improve Surveillance and Control Programs. J. Clin. Microbiol. 2015, 53, 2555–2565. [Google Scholar] [CrossRef]
- Onyilagha, C.; Quizon, K.; Zhmendak, D.; El Kanoa, I.; Truong, T.; Ambagala, T.; Clavijo, A.; Le, V.P.; Babiuk, S.; Ambagala, A. Development and Validation of an Indirect and Blocking ELISA for the Serological Diagnosis of African Swine Fever. Pathogens 2024, 13, 981. [Google Scholar] [CrossRef]
- Weeks, S.D.; Drinker, M.; Loll, P.J. Ligation independent cloning vectors for expression of SUMO fusions. Protein Expr. Purif. 2007, 53, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Stertman, L.; Palm, A.-K.E.; Zarnegar, B.; Carow, B.; Lunderius Andersson, C.; Magnusson, S.E.; Carnrot, C.; Shinde, V.; Smith, G.; Glenn, G.; et al. The Matrix-MTM adjuvant: A critical component of vaccines for the 21st century. Hum. Vaccines Immunother. 2023, 19, 2189885. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Liu, Q.; Li, L.; Zhang, R.; Chang, Y.; Zhao, J.; Liu, S.; Zhao, X.; Chen, X.; Sun, Y.; et al. Nanobodies against African swine fever virus p72 and CD2v proteins as reagents for developing two cELISAs to detect viral antibodies. Virol. Sin. 2024, 39, 478–489. [Google Scholar] [CrossRef] [PubMed]
- WOAH. African Swine Fever: WOAH Warns Veterinary Authorities and Pig Industry of Risk from Use of Sub-Standard Vaccines. Available online: https://www.woah.org/en/african-swine-fever-woah-warns-veterinary-authorities-and-pig-industry-of-risk-from-use-of-sub-standard-vaccines/ (accessed on 21 April 2025).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strong-Positive ASF Serum (C++) IPT Value 1:1280 | Weak-Positive ASF Serum (C+) IPT Value 1:180 | Negative ASF Serum (C−) C− |
|---|---|---|
| 1.143 ± 0.178 (1 SD) | 0.506 ± 0.089 (1 SD) | 0.147 ± 0.030 (1 SD) |
| 1.143 ± 0.357 (2 SD) | 0.506 ± 0.179 (2 SD) | 0.147 ± 0.060 (2 SD) |
| Range C++ 0.786 to 1.50 OD | Range C+ 0.326 to 0.596 OD | Range C− 0.08 to 0.208 OD |
| CV 17% | CV 3% |
| Laboratory “A” | Laboratory “B” | ||||||
|---|---|---|---|---|---|---|---|
| ID | Description | ELISA*-Ingezim PPA Compac Corta | ELISA*-Ingezim PPA Compac Larga | ELISA* Indirect % Bloqueo Compet | ELISA* Indirect Bloqueo Compet | OD of ASF-p72t Protein iELISA | ASF-p72t Protein iELISA |
| 2019.1 | Pig serum experimentally infected with PPA strain NH/68. Bleeding 30 dpi. | Neg/DUD | Pos | 65.97 | pos | 0.883 | Pos |
| 2019.2 | Pig serum (C2) infected and experimentally re-infected with ASFV (NHV/68 strain). Bleeding 63 dpi/28 dpi. | Pos | Pos | 94.04 | Pos | 0.73 | Pos |
| 2019.8 | Uninfected pork serum. Undiluted | Neg | Neg | 0.27 | Neg | 0.12 | Neg |
| 2019.10 | Uninfected pork serum. Undiluted | Neg | Neg | −2.07 | Neg | 0.179 | Neg |
| 2020.1 | Sow Filtered Serum 3161/NEG) LCV Animal Facility | Neg | Neg | 18.6 | Neg | 0.122 | Neg |
| 2020.3 | Sow Filtered Serum 3161/NEG) LCV Animal Facility | Neg | Neg | 9.9 | Neg | 0.168 | Neg |
| 2020.4 | 27 February 2020 and lyophilized on 6 January 2020. Proceeds from infection (and re-infection) experimentally with VPPA strain NHV/68. Indentation 63 dpi -28 dpi. | Pos | Pos | 102.8 | Pos | 0.558 | Pos |
| 2020.5 | 7 April 2019 and lyophilized on 7 September 2019. Proceeds of an infection (and re-infection) experimentally with the VPPA strain NHV/68. Bleeding 63 dpi -28 dpi. | Pos | Pos | 97.9 | Pos | 0.597 | Pos |
| 2020.6 | 27 February 2020 and lyophilized on 1 June 2020. Proceeds of an infection (and re-infection) experimentally with VPPA strain NHV/68. Bleeding 63 dpi -28 dpi. | DUD | Pos | 77.7 | Pos | 0.506 | Pos |
| 2020.8 | 27 February 2020 and lyophilized on 1 June 2020. Proceeds from infection (and re-infection) experimentally with VPPA strain NHV/68. Bleeding 63 dpi -28 dpi. | NEG/DUD | Pos | 63.2 | Pos | 0.337 | Pos |
| 2020.9 | 7 April 2019 and lyophilized on 7 September 2019. It is from an experimentally infected with VPPA strain NHV/68. Bleeding 30 dpi. | Pos | Pos | 96.9 | Pos | 0.614 | Pos |
| 2021.1 | 7 May 2021 and lyophilized on 19 July 2021. It is from an infection (and re-infection) experimentally with VPPA strain NHV/68. Bleeding 63 dpi -28 dpi. | Pos | Pos | 89.9 | Pos | 0.862 | Pos |
| 2021.2 | 27 February 2020 and lyophilized on 2 June 2020. It is from an infection (and re-infection) experimentally with VPPA strain NHV/68. Bleeding 63 dpi -28 dpi. | Neg/DUD | Pos | 75.2 | Pos | 0.97 | Pos |
| 2021.3 | 5 July 2021 and lyophilized on 19 July 2021. It is from an infection (and re-infection) experimentally with the VPPA strain NHV/68. Bleeding 63 dpi -28 dpi. | Pos | Pos | 93.4 | Pos | 0.543 | Pos |
| 2021.5 | 27 February 2020 and lyophilized on 2 June 2020. Proceeds from infection (and re-infection) experimentally with VPPA strain NHV/68. Bleeding 63 dpi -28 dpi. | Pos | Pos | 97.3 | Pos | 0.562 | Pos |
| 2021.7 | 27 June 2020 and lyophilized on 5 June 2020. Proceeds from infection (and re-infection) experimentally with VPPA strain NHV/68. Bleeding 30 dpi. | Pos | Pos | 99.2 | Pos | 0.476 | Pos |
| 2022.1 | Uninfected pork serum. Undiluted. | Neg | Neg | 0 | Neg | 0.147 | Neg |
| 2022.5 | 7 April 2022 and lyophilized on 11 May 2022. It is derived from an experimental infection (and re-infection) with the VPPA strain NHV/68. | Pos | Pos | 99 | Pos | 0.938 | Pos |
| IPT-Positive Serum | IPT-Negative Serum | Total | |
|---|---|---|---|
| Positive serums | a (22) | b (1) | r = a + b (23) |
| Negative serums | c (3) | d (43) | s = c + d (46) |
| Total | t = a + b (25) | u = b + d (44) | N = a + b + c + d (69) |
| Sensitivity | |||
| Specificity | |||
| Kappa index | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuevas-Romero, J.S.; Zavala-Ocampo, P.L.; Pina-Pedrero, S.; Ganges, L.; Muñoz-Aguilera, A.; García-Cambrón, J.B.; Rodriguez, F.; Ambagala, A.; Cerriteño-Sánchez, J.L. Cloning and Expression of a Truncated Form of the p72 Protein of the African Swine Fever Virus (ASFV) for Application in an Efficient Indirect ELISA System. Pathogens 2025, 14, 542. https://doi.org/10.3390/pathogens14060542
Cuevas-Romero JS, Zavala-Ocampo PL, Pina-Pedrero S, Ganges L, Muñoz-Aguilera A, García-Cambrón JB, Rodriguez F, Ambagala A, Cerriteño-Sánchez JL. Cloning and Expression of a Truncated Form of the p72 Protein of the African Swine Fever Virus (ASFV) for Application in an Efficient Indirect ELISA System. Pathogens. 2025; 14(6):542. https://doi.org/10.3390/pathogens14060542
Chicago/Turabian StyleCuevas-Romero, Julieta Sandra, Perla Lucero Zavala-Ocampo, Sonia Pina-Pedrero, Llilianne Ganges, Adriana Muñoz-Aguilera, José Bryan García-Cambrón, Fernando Rodriguez, Aruna Ambagala, and José Luis Cerriteño-Sánchez. 2025. "Cloning and Expression of a Truncated Form of the p72 Protein of the African Swine Fever Virus (ASFV) for Application in an Efficient Indirect ELISA System" Pathogens 14, no. 6: 542. https://doi.org/10.3390/pathogens14060542
APA StyleCuevas-Romero, J. S., Zavala-Ocampo, P. L., Pina-Pedrero, S., Ganges, L., Muñoz-Aguilera, A., García-Cambrón, J. B., Rodriguez, F., Ambagala, A., & Cerriteño-Sánchez, J. L. (2025). Cloning and Expression of a Truncated Form of the p72 Protein of the African Swine Fever Virus (ASFV) for Application in an Efficient Indirect ELISA System. Pathogens, 14(6), 542. https://doi.org/10.3390/pathogens14060542