
An Analysis of the Mycotoxins, Cytotoxicity, and Biodiversity of Airborne Molds Belonging to Aspergillus Genera Isolated from the Zoological Garden

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Method
2.3. Characterization of Meteorological Conditions
2.4. Morphological and Molecular Identification of Molds
2.5. MTT Test for Assessing Fungal Cytotoxicity
2.6. Sample Preparation and Mycotoxin Analysis
3. Results and Discussion
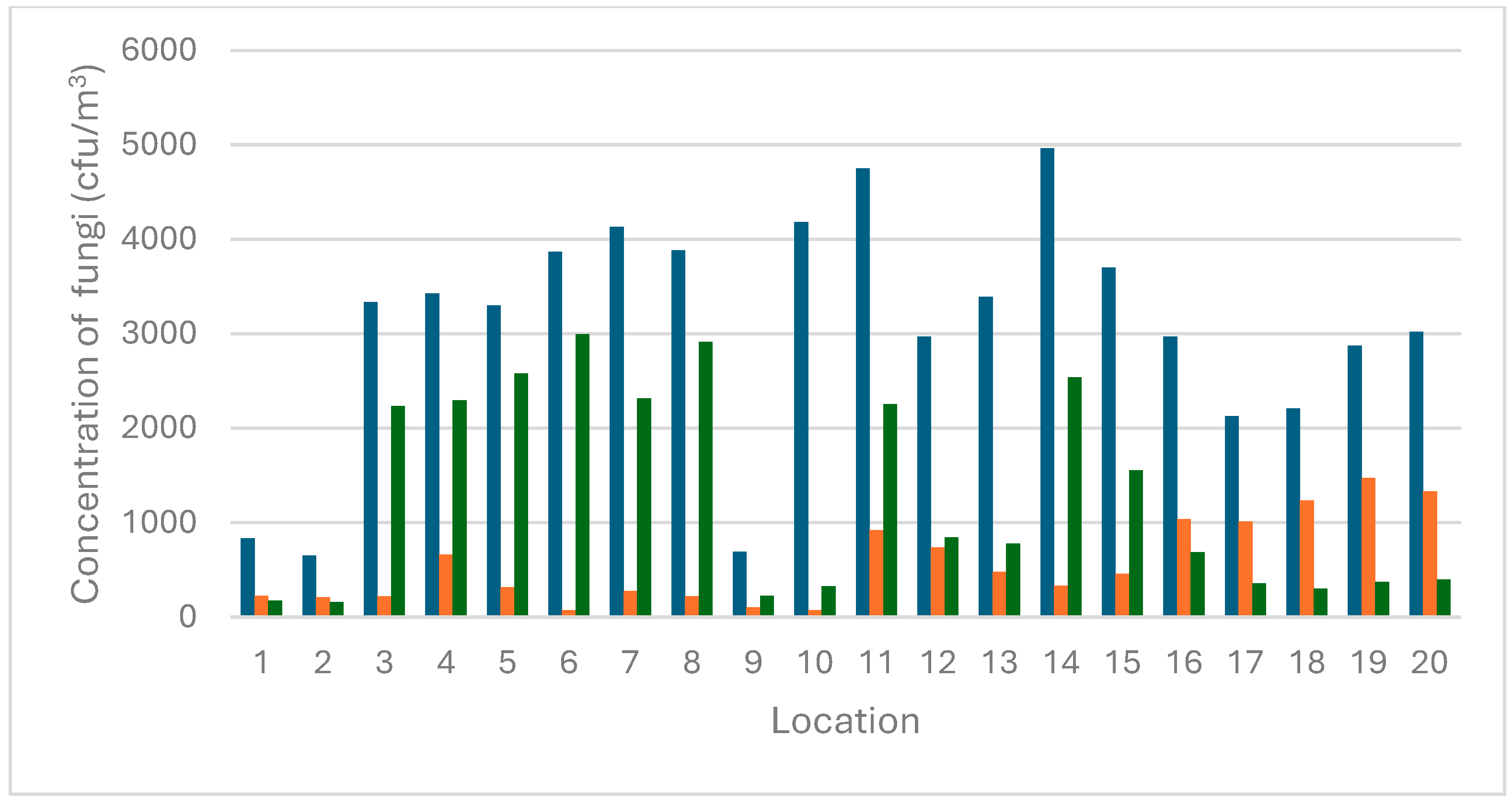
3.1. Concentration Levels of Airborne Molds
3.2. The Biodiversity of Airborne Fungi Belonging to Aspergillus Genera
3.3. Analysis of Cytotoxicity and Mycotoxin Production
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Al-Shaarani, M.; Pecoraro, L. A review of pathogenic airborne fungi and bacteria: Unveiling occurrence, sources, and profound human health implication. Front. Microbiol. 2024, 15, 1–15. [Google Scholar] [CrossRef]
- Martony, M.; Nollens, H.; Tucker, M.; Henry, L.; Schmitt, T.; Hernandez, J. Prevalence of and environmental factors associated with aerosolised Aspergillus spores at a zoological park. Vet. Rec. Open 2019, 6, e000281. [Google Scholar] [CrossRef] [PubMed]
- Grzyb, J.; Pawlak, K. Impact of bacterial aerosol, particulate matter, and microclimatic parameters on animal welfare in Chorzów (Poland) zoological garden. Environ. Sci. Pollut. Res. Int. 2021, 28, 3318–3330. [Google Scholar] [CrossRef]
- Mousavi, B.; Hedayati, M.T.; Hedayati, N.; Ilkit, M.; Syedmousavi, S. Aspergillus species in indoor environments and their possible occupational and public health hazards. Curr. Med. Mycol. 2016, 2, 36–42. [Google Scholar] [CrossRef]
- Paulussen, C.; Hallsworth, J.E.; Álvarez-Pérez, S.; Nierman, W.C.; Hamill, P.G.; Blain, D.; Rediers, H.; Lievens, B. Ecology of aspergillosis: Insights into the pathogenic potency of Aspergillus fumigatus and some other Aspergillus species. Microb. Biotechnol. 2017, 10, 296–322. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, A.; Osman, M.; Dabboussi, F.; Rafei, R.; Mallat, H.; Papon, N.; Bouchara, J.P.; Hamze, M. Recent trends in the epidemiology, diagnosis, treatment, and mechanisms of resistance in clinical Aspergillus species: A general review with a special focus on the Middle Eastern and North African region. J. Infect. Public Health 2020, 13, 1–10. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Mogensen, J.M.; Johansen, M.; Larsen, T.O.; Frisvad, J.C. Review of secondary metabolites and mycotoxins from the Aspergillus niger group. Anal. Bioanal. Chem. 2009, 395, 1225–1242. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar]
- Hanelt, M.; Gareis, M.; Kollarczik, B. Cytotoxicity of mycotoxins evaluated by the MTT cell culture assay. Mycopathologia 1994, 128, 164–174. [Google Scholar] [CrossRef]
- Viegas, C.; Gomes, B.; Cervantes, R.; Moreira, S.; Dias, M.; Pena, P.; Carolino, E.; Twarużek, M.; Kosicki, R.; Soszczyńska, E.; et al. Microbial contamination in grocery stores from Portugal and Spain—The neglected indoor environment to be tackled in the scope of the One Health approach. Sci. Total Environ. 2023, 875, 162602. [Google Scholar] [CrossRef] [PubMed]
- Plewa-Tutaj, K.; Twarużek, M.; Kosicki, R.; Soszczyńska, E. Analysis of mycotoxins and cytotoxicity of airborne molds isolated from the zoological garden—Screening research. Pathogens 2024, 13, 294. [Google Scholar] [CrossRef]
- Grzyb, J.; Lenart-Boroń, A. Size distribution and concentration of fungal aerosol in animal premises of a zoological garden. Aerobiologia 2020, 36, 233–248. [Google Scholar]
- Wei, Y.; Liu, T.; Zhang, W.; Zhang, W. The toxic mechanism of gliotoxins and biosynthetic strategies for toxicity prevention. Int. J. Mol. Sci. 2021, 22, 13510. [Google Scholar] [CrossRef]
- Melo, A.M.; Stevens, D.A.; Tell, L.A.; Veríssimo, C.; Sabino, R.; Xavier, M.O. Aspergillosis, Avian Species and the One Health Perspective: The Possible Importance of Birds in Azole Resistance. Microorganisms 2020, 8, 2037. [Google Scholar] [CrossRef] [PubMed]
- de Jong, J.E.; Heuvelink, A.E.; Dieste Pérez, L.; Holstege, M.M.C. Aspergillus spp., aspergillosis and azole usage in animal species in Europe: Results from a multisectoral survey and review of recent literature. Med. Mycol. 2025, 25, 63. [Google Scholar] [CrossRef]
- Itoh, M.; Matsumoto, N.; Hagino, K.; Sawayama, N.; Kuwayama, M.; Yamada, K.; Toyotome, T. Nationwide survey about the occurrence of aspergillosis in captive penguins in zoos and aquariums in Japan. Animals 2023, 13, 1913. [Google Scholar] [CrossRef]
- Skóra, J.; Sulyok, M.; Nowak, A.; Otlewska, A.; Gutarowska, B. Toxinogenicity and cytotoxicity of Alternaria, Aspergillus and Penicillium moulds isolated from working environments. Int. J. Environ. Sci. Technol. 2017, 14, 595–608. [Google Scholar]
- Twarużek, M.; Zastempowska, E.; Soszczyńska, E.; Ałtyn, I. The use of in vitro assays for the assessment of cytotoxicity on the example of MTT test. Acta Univ. Lodz. Folia Biol. Oecologica 2018, 14, 23–32. [Google Scholar] [CrossRef]
- Gniadek, A.; Krzyściak, P.; Twarużek, M.; Macura, A.B. Occurrence of fungi and cytotoxicity of the species: Aspergillus ochraceus, Aspergillus niger and Aspergillus flavus isolated from the air of hospital wards. Int. J. Occup. Med. Environ. Health 2020, 30, 231–239. [Google Scholar] [CrossRef]
- Scharf, D.H.; Heinekamp, T.; Remme, N.; Hortschansky, P.; Brakhage, A.A.; Hertweck, C. Biosynthesis and function of gliotoxin in Aspergillus fumigatus. Appl. Microbiol. Biotechnol. 2012, 93, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Rai, R.V.; Karim, A.A. Mycotoxins in Food and Feed: Present Status and Future Concerns. Compr. Rev. Food Sci. Food Saf. 2009, 17, 692–712. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2023, 16, 497–516. [Google Scholar] [CrossRef]
- Speijers, G.J.; Speijers, M.H. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef]
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Location | Average T (°C) | Average H (%) | Total cfu/m3 | cfu/m3 Aspergillus | cfu/m3 Penicillium | cfu/m3 Different Species |
|---|---|---|---|---|---|---|
| 1 | 22.13 | 41.28 | 837 | 225 | 175 | 437 |
| 2 | 21.83 | 37.38 | 650 | 211 | 162 | 275 |
| 3 | 16.48 | 68.05 | 3335 | 221 | 2236 | 878 |
| 4 | 17.13 | 70.45 | 3425 | 662 | 2295 | 468 |
| 5 | 16.83 | 67.05 | 3300 | 319 | 2581 | 400 |
| 6 | 19.35 | 56.43 | 3869 | 75 | 2994 | 800 |
| 7 | 17.38 | 51.78 | 4134 | 275 | 2318 | 1550 |
| 8 | 18.65 | 64.03 | 3886 | 221 | 2914 | 750 |
| 9 | 19.88 | 42.60 | 694 | 106 | 225 | 362 |
| 10 | 15.5 | 75.98 | 4181 | 75 | 325 | 3781 |
| 11 | 21.16 | 61.50 | 4750 | 922 | 2257 | 1571 |
| 12 | 20.80 | 62.25 | 2972 | 736 | 843 | 1393 |
| 13 | 45798 | 63.63 | 3393 | 479 | 779 | 2137 |
| 14 | 20.93 | 61.65 | 4963 | 331 | 2538 | 2094 |
| 15 | 21.13 | 60.80 | 3700 | 457 | 1557 | 1686 |
| 16 | 22.53 | 70.93 | 2969 | 1037 | 687 | 1243 |
| 17 | 24.38 | 58.80 | 2128 | 1014 | 357 | 757 |
| 18 | 24.60 | 59.78 | 2212 | 1237 | 300 | 675 |
| 19 | 24.80 | 58.28 | 2875 | 1475 | 375 | 1025 |
| 20 | 24.62 | 61.38 | 3020 | 1330 | 400 | 1290 |
| No | Species | Zoological Garden | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | ||
| 1 | A.ochraceus | + | + | + | + | + | + | ||||||||||||||
| 2 | A.flavus | + | + | + | + | ||||||||||||||||
| 3 | A.fumigatus | + | + | + | + | + | + | + | |||||||||||||
| 4 | A.steynii | + | + | + | + | ||||||||||||||||
| 5 | A.tamarri | + | + | + | |||||||||||||||||
| 6 | A.pseudoglaucus | + | |||||||||||||||||||
| 7 | A.niger | + | + | + | + | + | + | + | |||||||||||||
| 8 | A.nomius | + | |||||||||||||||||||
| 9 | A.westerdijikiae | + | + | + | + | ||||||||||||||||
| 10 | A.ostianus | + | + | + | + | ||||||||||||||||
| 11 | A.tubingensis | + | + | + | |||||||||||||||||
| 12 | A. sydowii | + | + | ||||||||||||||||||
| 13 | A.elegans | + | + | ||||||||||||||||||
| 14 | Aspergillus section Nigri | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |||||
| 15 | Aspergillus section Circumdati | + | + | + | + | + | + | + | + | + | + | + | |||||||||
| 16 | Aspergillus section Fumigati | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 17 | Aspergillus section Flavi | + | + | + | + | + | + | + | + | + | + | + | + | + | + | ||||||
| 18 | Aspergillus section Nidulantes | + | |||||||||||||||||||
| Species | IC50 [cm2/mL] |
|---|---|
| Section Flavi 4cz | 3.64 |
| Section Flavi 5bz | 3.9 |
| Section Flavi 14bz | 5.67 |
| A. flavus 15w | 2.07 |
| A. fumigatus 1aj | 0.09 |
| A. fumigatus 5aj | 0.17 |
| A. fumigatus 5cj | 0.17 |
| A. fumigatus 7l | 0.3 |
| A. fumigatus 8w | 0.21 |
| A. fumigatus 9cl | 0.36 |
| A. fumigatus 10z | 4.37 |
| A. fumigatus 19l | 0.14 |
| A. fumigatus 26l | 0.41 |
| A. tamarii 1l | 18.36 |
| A. tamarii 14al | 10.24 |
| A. tamarii 6w | 9.73 |
| A. tamarii 12z | 8.77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plewa-Tutaj, K.; Chmielewska, Z.; Twarużek, M.; Kosicki, R.; Soszczyńska, E. An Analysis of the Mycotoxins, Cytotoxicity, and Biodiversity of Airborne Molds Belonging to Aspergillus Genera Isolated from the Zoological Garden. Pathogens 2025, 14, 332. https://doi.org/10.3390/pathogens14040332
Plewa-Tutaj K, Chmielewska Z, Twarużek M, Kosicki R, Soszczyńska E. An Analysis of the Mycotoxins, Cytotoxicity, and Biodiversity of Airborne Molds Belonging to Aspergillus Genera Isolated from the Zoological Garden. Pathogens. 2025; 14(4):332. https://doi.org/10.3390/pathogens14040332
Chicago/Turabian StylePlewa-Tutaj, Kinga, Zuzanna Chmielewska, Magdalena Twarużek, Robert Kosicki, and Ewelina Soszczyńska. 2025. "An Analysis of the Mycotoxins, Cytotoxicity, and Biodiversity of Airborne Molds Belonging to Aspergillus Genera Isolated from the Zoological Garden" Pathogens 14, no. 4: 332. https://doi.org/10.3390/pathogens14040332
APA StylePlewa-Tutaj, K., Chmielewska, Z., Twarużek, M., Kosicki, R., & Soszczyńska, E. (2025). An Analysis of the Mycotoxins, Cytotoxicity, and Biodiversity of Airborne Molds Belonging to Aspergillus Genera Isolated from the Zoological Garden. Pathogens, 14(4), 332. https://doi.org/10.3390/pathogens14040332