The Effect of Helicobacter pylori Gene Combinations of cagA, cagE, virB11, vacA, and babA on the Outcome of Gastric Disease in a Southern Moroccan Population

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Specimens
2.2. Histopathological Exams
2.3. DNA Extraction
2.4. The Detection of H. pylori and Its Virulence Genes
2.5. Statistical Analysis
3. Results
3.1. Characteristics of Infected Patients
3.2. The Distribution of H. pylori Genotypes in Infected Patients
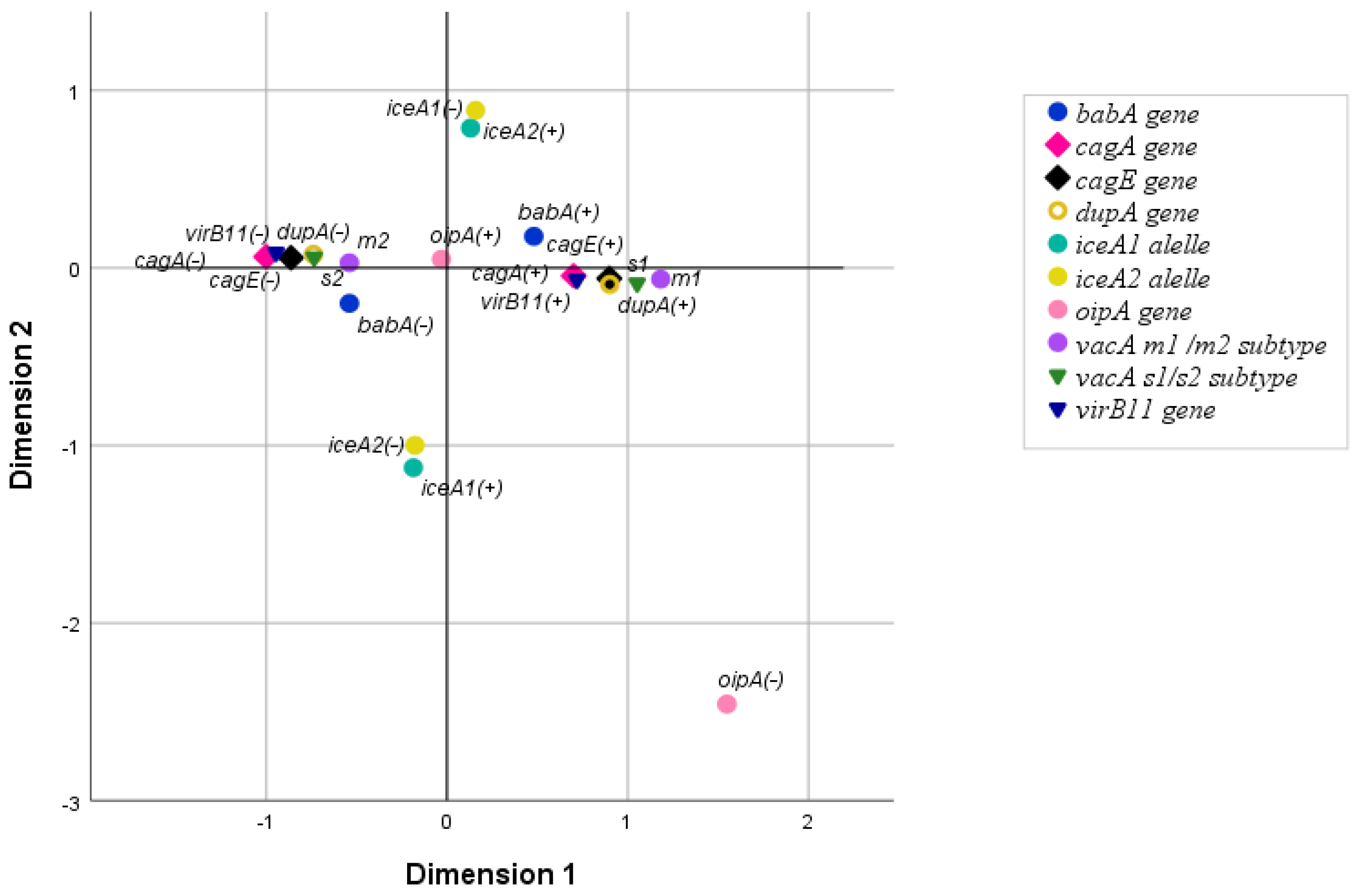
3.3. The Relationship Between H. pylori Virulence Genes
3.4. The Relationship Between Virulence Factors and Sex
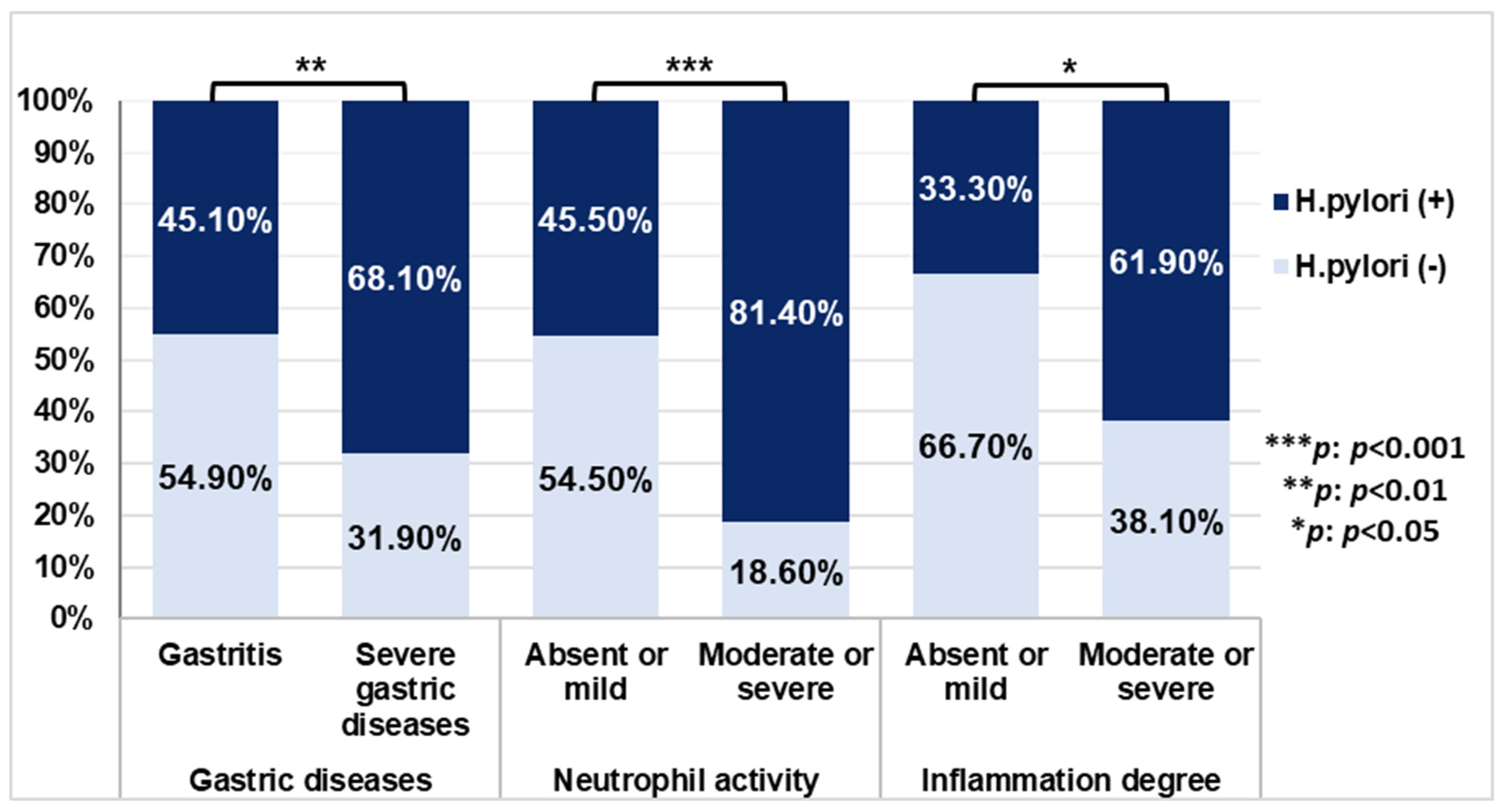
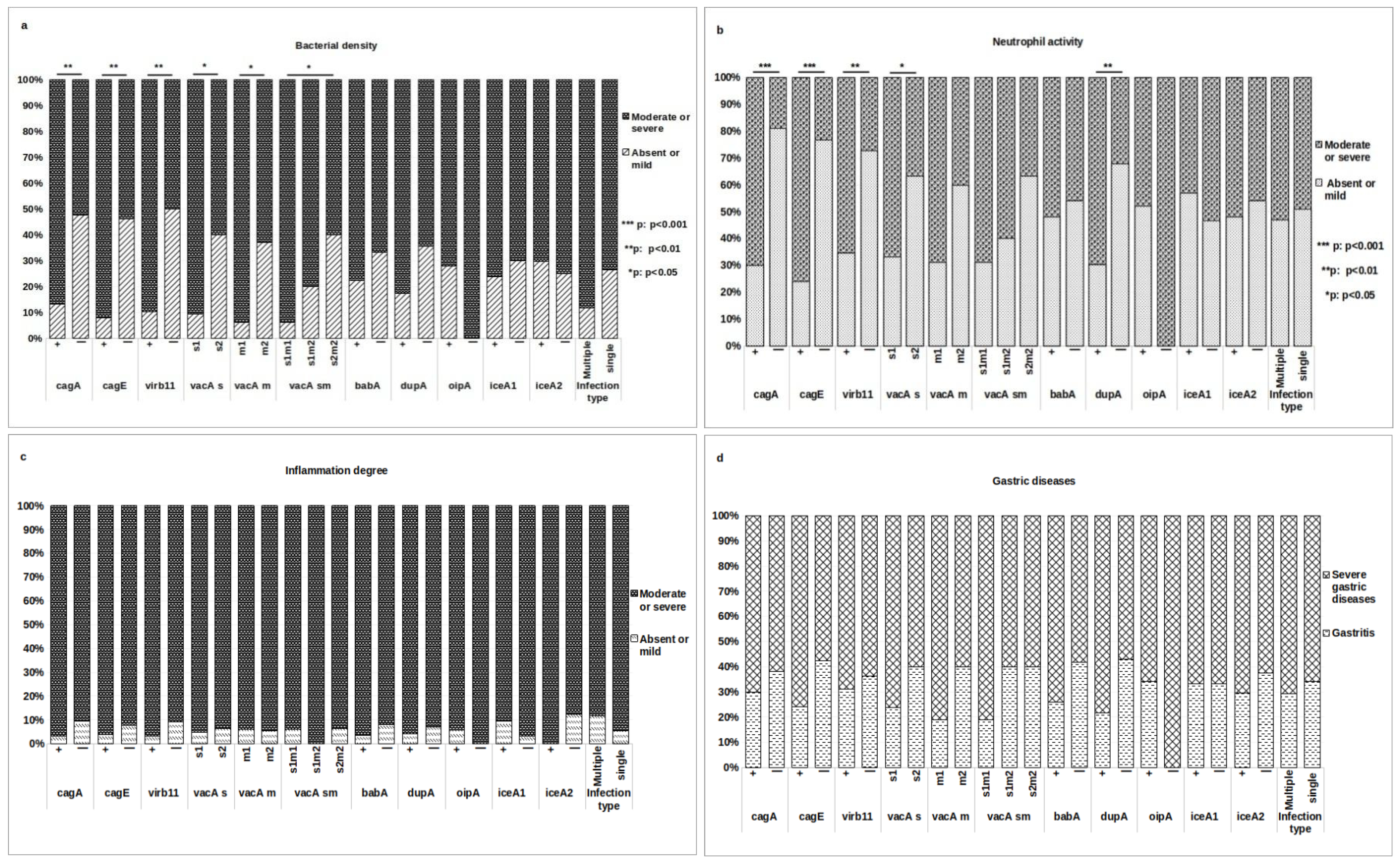
3.5. Association of H. pylori Genes with Histopathological Outcomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MCA | multiple correspondence analysis |
References
- Dixon, M.F. Pathology of Gastritis and Peptic Ulceration. In Helicobacter Pylori: Physiology and Genetics; Mobley, H.L.T., Mendz, G.L., Hazell, S.L., Eds.; ASM Press: Washington, DC, USA, 2001; pp. 457–469. [Google Scholar]
- Carlosama-Rosero, Y.H.; Bolaños-Bravo, H.; Sierra-Tórres, C.H.; Rosero, E.A. Asociación de Los Genotipos CagA, VacA e IceA de H. Pylori Con La Gastritis Crónica y Folicular En Una Población Colombiana Con Alto Riesgo de Cáncer Gástrico. Rev. Gastroenterol. Mex. 2019, 84, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Cheok, Y.Y.; Lee, C.Y.Q.; Cheong, H.C.; Vadivelu, J.; Looi, C.Y.; Abdullah, S.; Wong, W.F. An Overview of Helicobacter Pylori Survival Tactics in the Hostile Human Stomach Environment. Microorganisms 2021, 9, 2502. [Google Scholar] [CrossRef] [PubMed]
- Pereira, W.; Ferraz, M.; Zabaglia, L.; de Labio, R.; Orcini, W.; Bianchi Ximenez, J.; Neto, A.; Payão, S.L.; Rasmussen, L. Association among H. Pylori Virulence Markers DupA, CagA and VacA in Brazilian Patients. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 1. [Google Scholar] [CrossRef] [PubMed]
- Boubrik, F.; Belmouden, A.; El Kadmiri, N. Potential Non-Invasive Biomarkers of Helicobacter Pylori-Associated Gastric Cancer. J. Gastrointest. Cancer 2022, 53, 1113–1120. [Google Scholar] [CrossRef]
- Oktem-Okullu, S.; Tiftikci, A.; Saruc, M.; Cicek, B.; Vardareli, E.; Tozun, N.; Kocagoz, T.; Sezerman, U.; Yavuz, A.S.; Sayi-Yazgan, A. Multiplex-PCR-Based Screening and Computational Modeling of Virulence Factors and T-Cell Mediated Immunity in Helicobacter Pylori Infections for Accurate Clinical Diagnosis. PLoS ONE 2015, 10, e0136212. [Google Scholar] [CrossRef]
- Shiota, S.; Matsunari, O.; Watada, M.; Hanada, K.; Yamaoka, Y. Systematic Review and Meta-Analysis: The Relationship between the Helicobacter Pylori DupA Gene and Clinical Outcomes. Gut Pathog. 2010, 2, 13. [Google Scholar] [CrossRef]
- Yin, L.; Liu, F.; Guo, C.; Wang, Q.; Pan, K.; Xu, L.; Xiong, Y.; Chen, Y.; Chen, Z. Analysis of Virulence Diversity of 73 Helicobacter pylori Strains Isolated in Guizhou Province, China. Mol. Med. Rep. 2018, 18, 4611–4620. [Google Scholar] [CrossRef]
- de Lima Silva, L.L.; Oliveira, A.K.S.; Gama, A.R.; Ramos, A.F.P.L.; Silva, A.M.T.C.; Blanco, A.J.V.; Vieira, J.D.G.; Rasmussem, L.T.; Carneiro, L.C.; Barbosa, M.S. Helicobacter Pylori Virulence DupA Gene: Risk Factor or Protective Factor? Braz. J. Microbiol. 2021, 52, 1921–1927. [Google Scholar] [CrossRef]
- Dabiri, H.; Jafari, F.; Baghaei, K.; Shokrzadeh, L.; Abdi, S.; Pourhoseingholi, M.A.; Mohammadzadeh, A. Prevalence of Helicobacter Pylori VacA, CagA, CagE, OipA, IceA, BabA2 and BabB Genotypes in Iranian Dyspeptic Patients. Microb. Pathog. 2017, 105, 226–230. [Google Scholar] [CrossRef]
- Banga Ndzouboukou, J.; Lei, Q.; Ullah, N.; Zhang, Y.; Hao, L.; Fan, X. Helicobacter Pylori Adhesins: HpaA a Potential Antigen in Experimental Vaccines for H. Pylori. Helicobacter 2021, 26, 12758. [Google Scholar] [CrossRef]
- Almeida, N.; Donato, M.M.; Romãozinho, J.M.; Luxo, C.; Cardoso, O.; Cipriano, M.A.; Marinho, C.; Fernandes, A.; Sofia, C. Correlation of Helicobacter Pylori Genotypes with Gastric Histopathology in the Central Region of a South-European Country. Dig. Dis. Sci. 2015, 60, 74–85. [Google Scholar] [CrossRef] [PubMed]
- El Khadir, M.; Alaoui Boukhris, S.; Benajah, D.-A.; El Rhazi, K.; Ibrahimi, S.A.; El Abkari, M.; Harmouch, T.; Nejjari, C.; Mahmoud, M.; Benlemlih, M.; et al. VacA and CagA Status as Biomarker of Two Opposite End Outcomes of Helicobacter Pylori Infection (Gastric Cancer and Duodenal Ulcer) in a Moroccan Population. PLoS ONE 2017, 12, e0170616. [Google Scholar] [CrossRef] [PubMed]
- Shariq, M.; Kumar, N.; Kumari, R.; Kumar, A.; Subbarao, N.; Mukhopadhyay, G. Biochemical Analysis of CagE: A VirB4 Homologue of Helicobacter Pylori Cag-T4SS. PLoS ONE 2015, 10, e0142606. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Khara, P.; Song, L.; Lin, A.S.; Frick-Cheng, A.E.; Harvey, M.L.; Cover, T.L.; Christie, P.J. In Situ Molecular Architecture of the Helicobacter Pylori Cag Type IV Secretion System. mBio 2019, 10, e00849-19. [Google Scholar] [CrossRef]
- Dixon, M.F.; Genta, R.M.; Yardley, J.H.; Correa, P. For the International Workshop on the Histopathology of Gast Histological Classif Ication of Gastritis and Helicobacter Pylori Infection: An Agreement at Last? Helicobacter 1997, 2, 17–24. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Kwon, D.H.; Graham, D.Y. A M r 34,000 Proinflammatory Outer Membrane Protein (OipA) of Helicobacter Pylori. Proc. Natl. Acad. Sci. USA 2000, 97, 7533–7538. [Google Scholar] [CrossRef]
- Atherton, J.C.; Cover, T.L.; Twells, R.J.; Morales, M.R.; Hawkey, C.J.; Blaser, M.J. Simple and Accurate PCR-Based System for Typing Vacuolating Cytotoxin Alleles of Helicobacter Pylori. J. Clin. Microbiol. 1999, 37, 2979–2982. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Kodama, T.; Gutierrez, O.; Kim, J.G.; Kashima, K.; Graham, D.Y. Relationship between Helicobacter Pylori IceA, CagA, and VacA Status and Clinical Outcome: Studies in Four Different Countries. J. Clin. Microbiol. 1999, 37, 2274–2279. [Google Scholar] [CrossRef]
- Atherton, J.C.; Cao, P.; Peek, R.M.; Tummuru, M.K.R.; Blaser, M.J.; Cover, T.L. Mosaicism in Vacuolating Cytotoxin Alleles of Helicobacter Pylori. Association of Specific VacA Types with Cytotoxin Production and Peptic Ulceration. J. Biol. Chem. 1995, 270, 17771–17777. [Google Scholar] [CrossRef]
- Lima, V.P.; Silva-Fernandes, I.J.d.L.; Alves, M.K.S.; Rabenhorst, S.H.B. Prevalence of Helicobacter Pylori Genotypes (VacA, CagA, CagE and VirB11) in Gastric Cancer in Brazilian’s Patients: An Association with Histopathological Parameters. Cancer Epidemiol. 2011, 35, e32–e37. [Google Scholar] [CrossRef]
- Argent, R.H.; Zhang, Y.; Atherton, J.C. Simple Method for Determination of the Number of Helicobacter Pylori CagA Variable-Region EPIYA Tyrosine Phosphorylation Motifs by PCR. J. Clin. Microbiol. 2005, 43, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Homan, M.; Šterbenc, A.; Kocjan, B.J.; Luzar, B.; Zidar, N.; Orel, R.; Poljak, M. Prevalence of the Helicobacter Pylori BabA2 Gene and Correlation with the Degree of Gastritis in Infected Slovenian Children. Antonie Van Leeuwenhoek 2014, 106, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, M.; Lehn, N.; Neumayer, N.; Borén, T.; Rad, R.; Schepp, W.; Miehlke, S.; Classen, M.; Prinz, C. Clinical Relevance of the Helicobacter Pylori Gene for Blood-Group Antigen-Binding Adhesin. Proc. Natl. Acad. Sci. USA 1999, 96, 12778–12783. [Google Scholar] [CrossRef] [PubMed]
- El Khadir, M.; Boukhris Alaoui, S.; Benajah, D.; Ibrahimi, S.A.; Chbani, L.; El Abkari, M.; Bennani, B. VacA Genotypes and cagA-EPIYA-CMotifs of Helicobacter Pylori and Gastric Histopathological Lesions. Int. J. Cancer 2020, 147, 3206–3214. [Google Scholar] [CrossRef]
- Subsomwong, P.; Miftahussurur, M.; Uchida, T.; Vilaichone, R.; Ratanachu-ek, T.; Mahachai, V.; Yamaoka, Y. Prevalence, Risk Factors, and Virulence Genes of Helicobacter Pylori among Dyspeptic Patients in Two Different Gastric Cancer Risk Regions of Thailand. PLoS ONE 2017, 12, e0187113. [Google Scholar] [CrossRef]
- Lee, D.-H.; Ha, J.-H.; Shin, J.-I.; Kim, K.-M.; Choi, J.; Park, S.; Park, J.-S.; Seo, J.-H.; Park, J.-S.; Shin, M.-K.; et al. Increased Risk of Severe Gastric Symptoms by Virulence Factors VacAs1c, AlpA, BabA2, and Hop Z in Helicobacter Pylori Infection. J. Microbiol. Biotechnol. 2021, 31, 368–379. [Google Scholar] [CrossRef]
- Molina-Castro, S.; Garita-Cambronero, J.; Malespín-Bendaña, W.; Une, C.; Ramírez, V. Virulence Factor Genotyping of Helicobacter Pylori Isolated from Costa Rican Dyspeptic Patients. Microb. Pathog. 2019, 128, 276–280. [Google Scholar] [CrossRef]
- Ben Mansour, K.; Fendri, C.; Zribi, M.; Masmoudi, A.; Labbene, M.; Fillali, A.; Ben Mami, N.; Najjar, T.; Meherzi, A.; Sfar, T.; et al. Prevalence of Helicobacter Pylori VacA, CagA, IceA and OipA Genotypes in Tunisian Patients. Ann. Clin. Microbiol. Antimicrob. 2010, 9, 10. [Google Scholar] [CrossRef]
- Erzin, Y.; Koksal, V.; Altun, S.; Dobrucali, A.; Aslan, M.; Erdamar, S.; Dirican, A.; Kocazeybek, B. Prevalence of Helicobacter Pylori VacA, CagA, CagE, IceA, BabA2 Genotypes and Correlation with Clinical Outcome in Turkish Patients with Dyspepsia. Helicobacter 2006, 11, 574–580. [Google Scholar] [CrossRef]
- Jalilian, S.; Alvandi, A.; Jouybari, T.A.; Pajavand, H.; Abiri, R. Lack of Association between the Presence of DupA and BabA 2 Genes in Helicobacter Pylori and Gastroduodenal Disorders. Mol. Genet. Microbiol. Virol. 2017, 32, 55–61. [Google Scholar] [CrossRef]
- Chiurillo, M.A.; Moran, Y.; Cañas, M.; Valderrama, E.; Granda, N.; Sayegh, M.; Ramírez, J.L. Genotyping of Helicobacter Pylori Virulence-Associated Genes Shows High Diversity of Strains Infecting Patients in Western Venezuela. Int. J. Infect. Dis. 2013, 17, e750–e756. [Google Scholar] [CrossRef] [PubMed]
- Akeel, M.; Shehata, A.; Elhafey, A.; Elmakki, E.; Aboshouk, T.; Ageely, H.; Mahfouz, M. Helicobacter Pylori VacA, CagA and IceA Genotypes in Dyspeptic Patients from Southwestern Region, Saudi Arabia: Distribution and Association with Clinical Outcomes and Histopathological Changes. BMC Gastroenterol. 2019, 19, 16. [Google Scholar] [CrossRef] [PubMed]
- Idowu, A.; Mzukwa, A.; Harrison, U.; Palamides, P.; Haas, R.; Mbao, M.; Mamdoo, R.; Bolon, J.; Jolaiya, T.; Smith, S.; et al. Detection of Helicobacter Pylori and Its Virulence Genes (CagA, DupA, and VacA) among Patients with Gastroduodenal Diseases in Chris Hani Baragwanath Academic Hospital, South Africa. BMC Gastroenterol. 2019, 19, 73. [Google Scholar] [CrossRef] [PubMed]
- Elnosh, M.M.; Hamedelnil, Y.F.; Elshareef, W.A.; Abugrain, A.Y.; Osman, E.H.; Albasha, A.M.; Aseel, K.H.; Ali, F.Y.; Abdalla, W.M.; AbdAlla, A.B.; et al. The CagA, CagE, VacA, DupA and IceA1 Genes of Helicobacter Pylori in Sudanese Gastritis Patients: Distribution and Relationship with Clinical Outcomes and Histological Alterations. Malays. J. Microbiol. 2022, 18, 261–270. [Google Scholar] [CrossRef]
- El-Shenawy, A.; Diab, M.; Shemis, M.; El-Ghannam, M.; Salem, D.; Abdelnasser, M.; Shahin, M.; Abdel-Hady, M.; El-Sherbini, E.; Saber, M. Detection of Helicobacter Pylori VacA, CagA and IceA1 Virulence Genes Associated with Gastric Diseases in Egyptian Patients. Egypt. J. Med. Hum. Genet. 2017, 18, 365–371. [Google Scholar] [CrossRef]
- Lai, C.-H.; Huang, J.-C.; Chiang-Ni, C.; Li, J.-P.; Wu, L.-T.; Wu, H.-S.; Sun, Y.-C.; Lin, M.-L.; Lee, J.-F.; Lin, H.-J. Mixed Infections of Helicobacter Pylori Isolated from Patients with Gastrointestinal Diseases in Taiwan. Gastroenterol. Res. Pract. 2016, 2016, 7521913. [Google Scholar] [CrossRef]
- Boonyanugomol, W.; Kongkasame, W.; Palittapongarnpim, P.; Baik, S.-C.; Jung, M.; Shin, M.-K.; Kang, H.-L.; Lee, W.-K. Genetic Variation in the Cag Pathogenicity Island of Helicobacter Pylori Strains Detected from Gastroduodenal Patients in Thailand. Braz. J. Microbiol. 2020, 51, 1093–1101. [Google Scholar] [CrossRef]
- Šterbenc, A.; Jarc, E.; Poljak, M.; Homan, M. Helicobacter Pylori Virulence Genes. World J. Gastroenterol. 2019, 25, 4870–4884. [Google Scholar] [CrossRef]
- García-Gómez, E.; González-Pedrajo, B.; Camacho-Arroyo, I. Role of Sex Steroid Hormones in Bacterial-Host Interactions. Biomed. Res. Int. 2013, 2013, 928290. [Google Scholar] [CrossRef]
- Hosoda, K.; Shimomura, H.; Hayashi, S.; Yokota, K.; Hirai, Y. Steroid Hormones as Bactericidal Agents to Helicobacter Pylori. FEMS Microbiol Lett 2011, 318, 68–75. [Google Scholar] [CrossRef]
- Morgan, E.; Arnold, M.; Camargo, M.C.; Gini, A.; Kunzmann, A.T.; Matsuda, T.; Meheus, F.; Verhoeven, R.H.A.; Vignat, J.; Laversanne, M.; et al. The Current and Future Incidence and Mortality of Gastric Cancer in 185 Countries, 2020–40: A Population-Based Modelling Study. EClinicalMedicine 2022, 47, 101404. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Lacy, D.B.; Ohi, M.D. The Helicobacter Pylori Cag Type IV Secretion System. Trends Microbiol. 2020, 28, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Shahini Shams Abadi, M.; Ashrafi-Dehkordi, K.; Ahmadi, R.; Rahimian, G.; Mirzaei, Y.; Fereidani, R.; Shohan, M.; Azadegan-Dehkordi, F. Frequency of Virulence-Associated Genotypes of Helicobacter Pylori and Their Correlation with Clinical Outcome and Histological Parameters in Infected Patients. Heliyon 2021, 7, e07610. [Google Scholar] [CrossRef] [PubMed]
- Miernyk, K.M.; Bruden, D.; Rudolph, K.M.; Hurlburt, D.A.; Sacco, F.; McMahon, B.J.; Bruce, M.G. Presence of CagPAI Genes and Characterization of VacA s, i and m Regions in Helicobacter Pylori Isolated from Alaskans and Their Association with Clinical Pathologies. J. Med. Microbiol. 2020, 69, 218–227. [Google Scholar] [CrossRef]
- Chen, R.; Li, Y.; Chen, X.; Chen, J.; Song, J.; Yang, X.; Ye, L.; Wu, Z.; Xie, P.; Zhong, Q.; et al. DupA+H. Pylori Reduces Diversity of Gastric Microbiome and Increases Risk of Erosive Gastritis. Front. Cell. Infect. Microbiol. 2023, 13, 1103909. [Google Scholar] [CrossRef]
- Toller, I.M.; Neelsen, K.J.; Steger, M.; Hartung, M.L.; Hottiger, M.O.; Stucki, M.; Kalali, B.; Gerhard, M.; Sartori, A.A.; Lopes, M.; et al. Carcinogenic Bacterial Pathogen Helicobacter Pylori Triggers DNA Double-Strand Breaks and a DNA Damage Response in Its Host Cells. Proc. Natl. Acad. Sci. USA 2011, 108, 14944–14949. [Google Scholar] [CrossRef]
- Murata-Kamiya, N.; Hatakeyama, M. Helicobacter Pylori-induced DNA Double-stranded Break in the Development of Gastric Cancer. Cancer Sci. 2022, 113, 1909–1918. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Bacterial Density n (%) | Neutrophil Activity n (%) | Inflammation Degree n (%) | Gastric Diseases n (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Combinations | Absent or Mild | Moderate or Severe | Absent or Mild | Moderate or Severe | Absent or Mild | Moderate or Severe | Gastritis | Severe Gastric Diseases | |
| cagA (+), cagE (+), virB11 (+), vacA s1m1, babA (+) | (+) | 1/13 (7.7) | 12/13 (92.3) | 4/13 (30.8) | 9/13 (69.2) | 1/13 (7.7) | 12/13 (92.3) | 1/13 (7.7) | 12/13 (92.3) |
| (−) | 13/38 (34.2) | 25/38 (65.8) | 22/38 (57.9) | 16/38 (42.1) | 2/38 (5.3) | 36/38 (94.7) | 16/38 (42.1) | 22/38 (57.9) | |
| p-value | 0.082 | 0.091 | 1 | 0.038 * | |||||
| ORᵘ (CI 95%) | - | - | - | 8.73 (1.03–74.12) | |||||
| cagA (−), cagE (−) virB11 (−), vacA s2m2, babA (−) | (+) | 5/11 (45.5) | 6/11 (54.5) | 9/11 (81.8) | 2/11 (18.2) | 2/11 (18.2) | 9/11 (81.8) | 5/11 (45.5) | 6/11 (54.5) |
| (−) | 9/40 (22.5) | 31/40 (77.5) | 17/40 (42.5) | 23/40 (57.5) | 1/40 (2.5) | 39/40 (97.5) | 12/40 (30) | 28/40 (70) | |
| p-value | 0.148 | 0.021 * | 0.114 | 0.472 | |||||
| ORᵘ (CI 95%) | - | 0.16 (0.03–0.86) | - | - | |||||
| cagA (−), cagE (−), virB11 (−), vacA s2m2, babA (+) | (+) | 3/7 (42.9) | 4/7 (57.1) | 5/7 (71.4) | 2/7 (28.6) | 0/7 (0.0) | 7/7 (100) | 2/7 (28.6) | 5/7 (71.4) |
| (−) | 11/44 (25) | 33/44 (75) | 21/44 (47.7) | 23/44 (52.3) | 3/44 (6.8) | 41/44 (93.2) | 15/44 (34.1) | 29/44 (65.9) | |
| p-value | 0.376 | 0.419 | 1 | 1 | |||||
| ORᵘ (CI 95%) | - | - | - | - | |||||
| Follicular Gastritis n (%) | Gastric Atrophy n (%) | Intestinal Metaplasia n (%) | Gastric Cancer n (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Combinations | (+) | (−) | (+) | (−) | (+) | (−) | (+) | (−) | |
| cagA (+), cagE (+), virB11 (+), vacA s1m1, babA (+) | (+) | 6/13 (46.2) | 7/13 (53.8) | 0/13 (0.0) | 13/13 (100) | 2/13 (15.4) | 11/13 (84.6) | 3/13 (23.1) | 10/13 (76.9) |
| (−) | 11/38 (28.9) | 27/38 (71.1) | 6/38 (15.8) | 32/38 (84.2) | 1/38 (2.6) | 37/38 (97.4) | 1/38 (2.6) | 37/38 (97.4) | |
| p-value | 0.315 | 0.318 | 0.156 | 0.046 * | |||||
| ORᵘ (CI 95%) | - | - | - | 11.10 (1.04–118.57) | |||||
| cagA (−), cagE (−), virB11 (−), vacA s2m2, babA (−) | (+) | 4/11 (36.4) | 7/11 (63.6) | 1/11 (9.1) | 10/11 (90.9) | 0/11 (0.0) | 11/11 (100) | 0/11 (0.0) | 11/11 (100) |
| (−) | 13/40 (32.5) | 27/40 (67.5) | 5/40 (12.5) | 35/40 (87.5) | 3/40 (7.5) | 37/40 (92.5) | 4/40 (10) | 36/40 (90) | |
| p-value | 1 | 1 | 1 | 0.565 | |||||
| ORᵘ (CI 95%) | - | - | - | - | |||||
| cagA (−), cagE (−), virB11 (−), vacA s2m2, babA (+) | (+) | 2/7 (28.6) | 5/7 (71.4) | 3/7 (42.9) | 4/7 (57.1) | 0/7 (0.0) | 7/7 (100) | 0/7 (0.0) | 7/7 (100) |
| (−) | 15/44 (34.1) | 29/44 (65.9) | 3/44 (6.8) | 41/44 (93.2) | 3/44 (6.8) | 41/44 (93.2) | 4/44 (9.1) | 40/44 (90.9) | |
| p-value | 1 | 0.028 * | 1 | 1 | |||||
| ORᵘ (CI 95%) | - | 10.25 (1.53–68.62) | - | - | |||||
| Histopathological Outcomes | Bivariate Analysis | Multivariate Analysis | |||||
|---|---|---|---|---|---|---|---|
| Risk Factors | p-Value | ORᵘ (CI 95%) | p-Value | ORª (CI 95%) | |||
| Neutrophil activity n (%) | |||||||
| Absent or mild | Moderate or severe | ||||||
| cagE | (+) | 6 (24) | 19 (76) | p < 0.001 * | 10.56 (2.89–38.50) | 0.03 * | 4.99 (1.18–21.09) |
| (−) | 20 (76.9) | 6 (23.1) | ref | ref | |||
| Gastric diseases n (%) | |||||||
| Gastritis | Severe gastric diseases | ||||||
| cagA (+), cagE (+), virB11 (+), vacA s1m1, babA (+) | (+) | 1 (7.7) | 12 (92.3) | 0.038 * | 8.73 (1.03–74.12) | 0.047 * | 8.73 (1.03–74.12) |
| (−) | 16 (42.1) | 22 (57.9) | ref | ref | |||
| Follicular gastritis n (%) | |||||||
| (+) | (−) | ||||||
| Age | ≤40 | 12 (48) | 13 (52) | 0.029 * | 3.88 (1.11–13.55) | 0.013 * | 5.87 (1.46–23.58) |
| >40 | 5 (19.2) | 21 (80.8) | ref | ref | |||
| Bacterial density | Moderate or severe | 16 (43.2) | 21 (56.8) | 0.019 * | 9.90 (1.17–83.80) | 0.017 * | 15.38 (1.64–144.40) |
| Absent or mild | 1 (7.1) | 13 (92.9) | ref | ref | |||
| Gastric atrophy n (%) | |||||||
| (+) | (−) | ||||||
| cagA (−), cagE (−), virB11 (−), vacA s2m2, babA (+) | (+) | 3 (42.9) | 4 (57.1) | 0.028 * | 10.25 (1.53–68.62) | 0.016 * | 10.25 (1.53–68.62) |
| (−) | 3 (6.8) | 41 (93.2) | ref | ref | |||
| Gastric cancer n (%) | |||||||
| (+) | (−) | ||||||
| cagA (+), cagE (+), virB11 (+), vacA s1m1, babA (+) | (+) | 3 (23.1) | 10 (76.9) | 0.046 * | 11.10 (1.04–118.57) | 0.046 * | 11.10 (1.04–118.57) |
| (−) | 1 (2.6) | 37 (97.4) | ref | ref | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barhoine, M.; Moustaoui, F.; Hammani, O.; Aghrouch, M.; Lemkhente, Z.; Belhabib, Z.; Bajaddoub, Z.; Touyar, A.; Aqoudad, N.; Rherissi, B.; et al. The Effect of Helicobacter pylori Gene Combinations of cagA, cagE, virB11, vacA, and babA on the Outcome of Gastric Disease in a Southern Moroccan Population. Pathogens 2025, 14, 279. https://doi.org/10.3390/pathogens14030279
Barhoine M, Moustaoui F, Hammani O, Aghrouch M, Lemkhente Z, Belhabib Z, Bajaddoub Z, Touyar A, Aqoudad N, Rherissi B, et al. The Effect of Helicobacter pylori Gene Combinations of cagA, cagE, virB11, vacA, and babA on the Outcome of Gastric Disease in a Southern Moroccan Population. Pathogens. 2025; 14(3):279. https://doi.org/10.3390/pathogens14030279
Chicago/Turabian StyleBarhoine, Mariama, Fatima Moustaoui, Omayma Hammani, Mohamed Aghrouch, Zohra Lemkhente, Zineb Belhabib, Zineb Bajaddoub, Anass Touyar, Nourdin Aqoudad, Bouchra Rherissi, and et al. 2025. "The Effect of Helicobacter pylori Gene Combinations of cagA, cagE, virB11, vacA, and babA on the Outcome of Gastric Disease in a Southern Moroccan Population" Pathogens 14, no. 3: 279. https://doi.org/10.3390/pathogens14030279
APA StyleBarhoine, M., Moustaoui, F., Hammani, O., Aghrouch, M., Lemkhente, Z., Belhabib, Z., Bajaddoub, Z., Touyar, A., Aqoudad, N., Rherissi, B., El Kadmiri, N., Idaghdour, Y., Boubrik, F., & Belmouden, A. (2025). The Effect of Helicobacter pylori Gene Combinations of cagA, cagE, virB11, vacA, and babA on the Outcome of Gastric Disease in a Southern Moroccan Population. Pathogens, 14(3), 279. https://doi.org/10.3390/pathogens14030279