
Fusarium graminearum in Wheat—Management Strategies in Central Europe


Abstract
1. Introduction
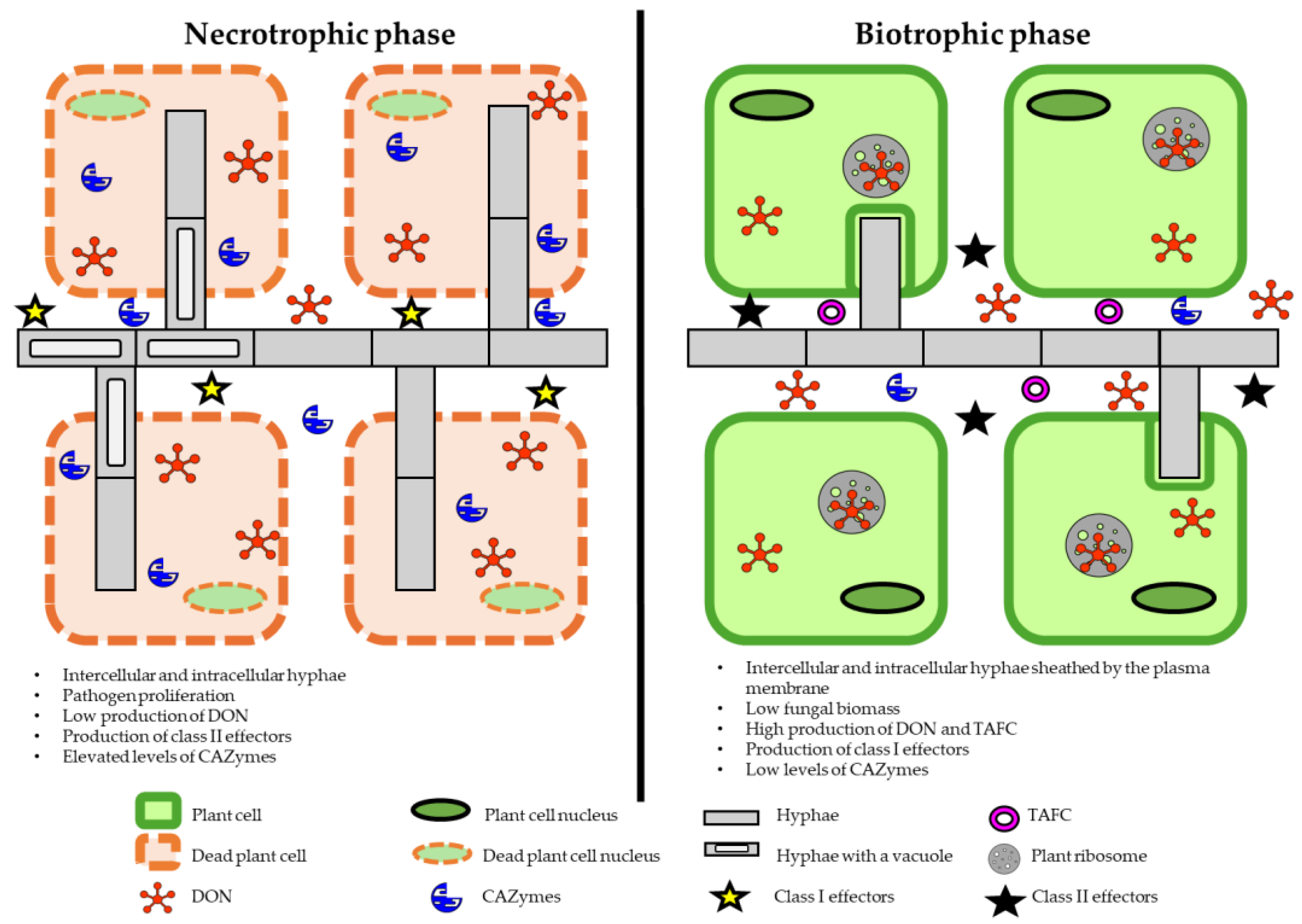
2. Spread and Infection Process of Fusarium graminearum
3. Fusarium graminearum Species Complex, Genotypes/Chemotypes, and Populations
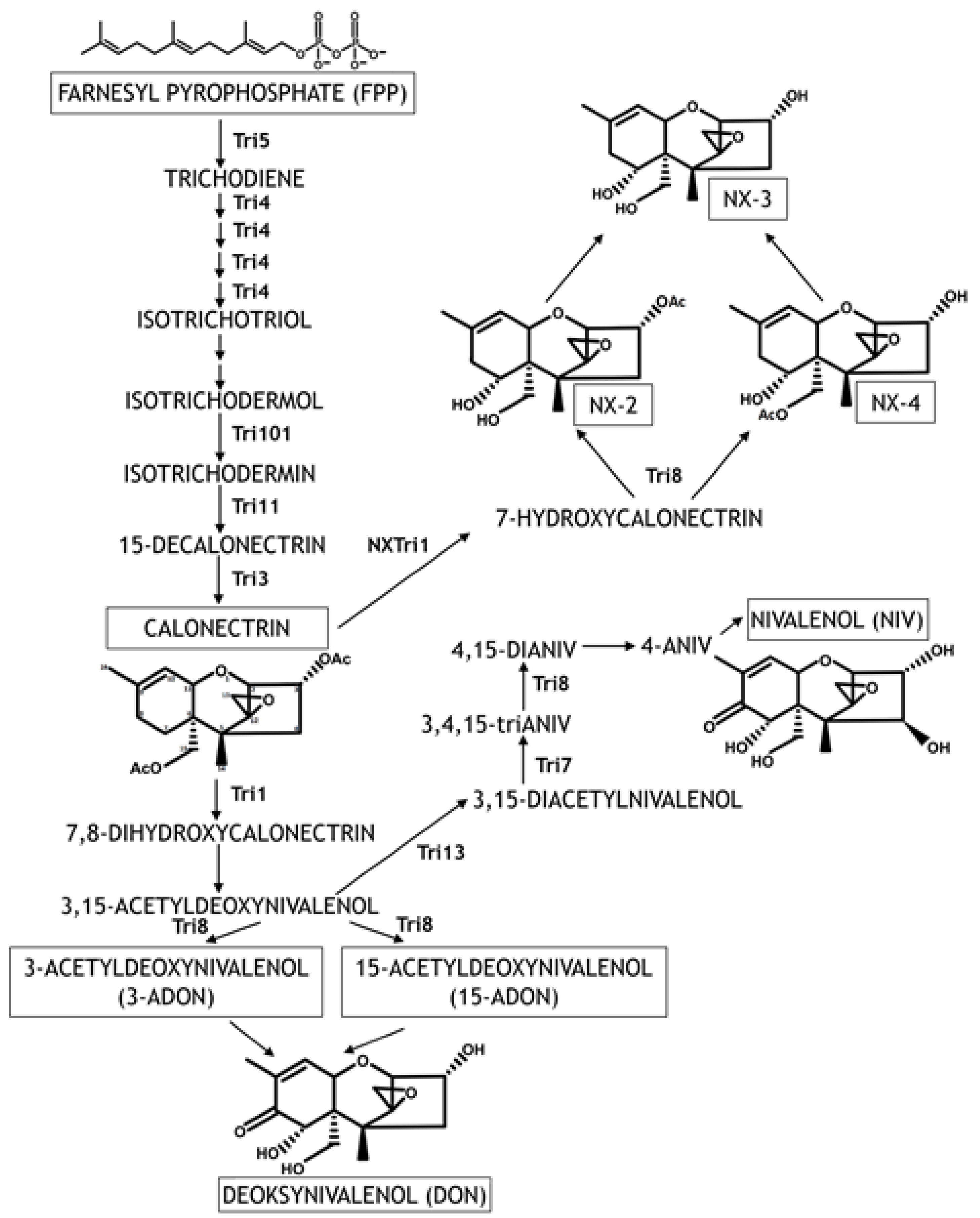
4. Trichothecene Synthesis—The Key to the Infection Process
5. The Occurrence of FHB and Mycotoxins in the Field
6. Fungicide Resistance in Fusarium graminearum and Wheat Biological Control
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kubaláková, M.; Kovárová, P.; Suchánková, P.; Cíhalíková, J.; Bartos, J.; Lucretti, S.; Watanabe, N.; Kianian, S.F.; Dolezel, J. Chromosome sorting in tetraploid wheat and its potential for genome analysis. Genetics 2005, 170, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Haile, J.K.; N’Diaye, A.; Walkowiak, S.; Nilsen, K.T.; Clarke, J.M.; Kutcher, H.R.; Steiner, B.; Buerstmayr, H.; Pozniak, C.J. Fusarium head blight in durum wheat: Recent status, breeding directions, and future research prospects. Phytopathology 2019, 109, 1664–1675. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Organization of the United Nations. Statistical Database. 2023. Available online: http://www.fao.org/faostat/en/#data (accessed on 22 July 2024).
- Tidiane, S.A.; Chiari, T.; Legesse, W.; Seid-Ahmed, K.; Ortiz, R.; van Ginkel, M.; Bassi, F.M. Durum wheat (Triticum durum Desf.): Origin, cultivation and potential expansion in Sub-Saharan Africa. Agronomy 2019, 9, 263. [Google Scholar] [CrossRef]
- Galal, A.A.; Safhi, F.A.; El-Hity, M.A.; Kamara, M.M.; Gamal El-Din, E.M.; Rehan, M.; Farid, M.; Behiry, S.I.; El-Soda, M.; Mansour, E. Molecular genetic diversity of local and exotic durum wheat genotypes and their combining ability for agronomic traits under water deficit and well-watered conditions. Life 2023, 13, 2293. [Google Scholar] [CrossRef] [PubMed]
- Ganeva, D.; Roumenina, E.; Dimitrov, P.; Gikov, A.; Bozhanova, V.; Dragov, R.; Jelev, G.; Taneva, K. Preharvest durum wheat yield, protein content, and protein yield estimation using unmanned aerial vehicle imagery and pléiades satellite data in field breeding experiments. Remote Sens. 2024, 16, 559. [Google Scholar] [CrossRef]
- Wachowska, U.; Stuper-Szablewska, K.; Perkowski, J. Yeasts isolated from wheat grain can suppress fusarium head blight and decrease trichothecene concentrations in bread wheat and durum wheat grain. Pol. J. Environ. Stud. 2020, 29, 4345–4360. [Google Scholar] [CrossRef] [PubMed]
- Rachoń, L.; Szumiło, G.; Stankowski, S. Comparison of selected technological value indicators of common wheat (Triticum aestivum spp. vulgare), durum wheat (Triticum durum) and spelt wheat (Triticum aestivum spp. spelta). Fragm. Agron. 2011, 28, 52–59. [Google Scholar]
- Wyzińska, M.; Sułek, A. Cultivation, importance and use of hard wheat. In Analysis, Production and Application of Substances and Components of Plant Origin; Maciąg, M., Maciąg, K., Eds.; Wydawnictwo Naukowe TYGIEL: Lublin, Poland, 2018; pp. 240–248. [Google Scholar]
- Alisaac, E.; Mahlein, A.-K. Fusarium head blight on wheat: Biology, modern detection and diagnosis and integrated disease management. Toxins 2023, 15, 192. [Google Scholar] [CrossRef]
- Ji, F.; He, D.; Olaniran, A.O.; Mokoena, M.P.; Xu, J.; Shi, J. Occurrence, toxicity, production and detection of Fusarium mycotoxin: A review. Food Prod. Process Nutr. 2019, 1, 6. [Google Scholar] [CrossRef]
- Ji, L.; Li, Q.; Wang, Y.; Burgess, L.W.; Sun, M.; Cao, K.; Kong, L. Monitoring of Fusarium species and trichothecene genotypes associated with Fusarium head blight on wheat in Hebei province, China. Toxins 2019, 11, 243. [Google Scholar] [CrossRef]
- Bilska, K.; Stuper-Szablewska, K.; Kulik, T.; Buśko, M.; Załuski, D.; Jurczak, S.; Perkowski, J. Changes in phenylpropanoid and trichothecene production by Fusarium culmorum and F. graminearum sensu stricto via exposure to flavonoids. Toxins 2018, 10, 110. [Google Scholar] [CrossRef] [PubMed]
- Crippin, T.; Renaud, J.B.; Sumarah, M.W.; Miller, J.D. Comparing genotype and chemotype of Fusarium graminearum from cereals in Ontario, Canada. PLoS ONE 2019, 14, e0216735. [Google Scholar] [CrossRef] [PubMed]
- Summerell, B.A. Resolving Fusarium: Current status of the genus. Annu. Rev. Phytopathol. 2019, 57, 323–339. [Google Scholar] [CrossRef]
- Zhu, Y.; Xi, J.; Yao, Y.; Xu, H.; Tang, C.; Wu, L. Characterizing the dynamic linkages between environmental changes and wheat Fusarium head blight epidemics. Ecol. Inform. 2024, 80, 102524. [Google Scholar] [CrossRef]
- Cavinder, B.; Sikhakolli, U.; Fellows, K.M.; Trail, F. Sexual development and ascospore discharge in Fusarium graminearum. J. Vis. Exp. 2012, 29, 3895. [Google Scholar] [CrossRef]
- Trail, F. For blighted waves of grain: Fusarium graminearum in the postgenomics era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef]
- Yoshida, M. Studies on the control of Fusarium head blight of barley and wheat and mycotoxin levels in grains based on time of infection and toxin accumulation. J. Gen. Plant Pathol. 2012, 78, 425–426. [Google Scholar] [CrossRef]
- Beccari, G.; Arellano, C.; Covarelli, L.; Tini, F.; Sulyok, M.; Cowger, C. Effect of wheat infection timing on Fusarium head blight causal agents and secondary metabolites in grain. Int. J. Food Microbiol. 2019, 290, 214–225. [Google Scholar] [CrossRef]
- Siou, D.; Gélisse, S.; Laval, V.; Repinçay, C.; Canalès, R.; Suffert, F.; Lannou, C. Effect of wheat spike infection and mycotoxin accumulation. Plant Pathol. 2014, 63, 390–399. [Google Scholar] [CrossRef]
- Kheiri, A.; Moosawi Jorf, S.A.; Malihipour, A. Infection process and wheat response to Fusarium head blight caused by Fusarium graminearum. Eur. J. Plant Pathol. 2019, 153, 489–502. [Google Scholar] [CrossRef]
- Boenisch, M.J.; Schäfer, W. Fusarium graminearum forms mycotoxin producing infection structures on wheat. BMC Plant Biol. 2011, 11, 110. [Google Scholar] [CrossRef] [PubMed]
- Buttar, Z.A.; Cheng, M.; Wei, P.; Zhang, Z.; Lv, C.; Zhu, C.; Ali, N.F.; Kang, G.; Wang, D.; Zhang, K. Update on the basic understanding of Fusarium graminearum virulence factors in common wheat research. Plants 2024, 13, 1159. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Zhao, X.; Fang, W.; Wu, H.; Abubakar, Y.S.; Lu, G.D.; Wang, Z.; Zheng, W. Spatiotemporal nature of Fusarium graminearum-wheat coleoptile interactions. Phytopathol. Res. 2019, 1, 26. [Google Scholar] [CrossRef]
- Brown, N.A.; Evans, J.; Mead, A.; Hammond-Kosack, K.E. A spatial temporal analysis of the Fusarium graminearum transcriptome during symptomless and symptomatic wheat infection. Mol. Plant Pathol. 2017, 18, 1295–1312. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Cho, E.J.; Lee, S.; Lee, Y.S.; Yun, S.H. Functional analyses of individual mating-type transcripts at MAT loci in Fusarium graminearum and Fusarium asiaticum. FEMS Microbiol. Lett. 2012, 337, 89–96. [Google Scholar] [CrossRef]
- Del Ponte, E.M.; Moreira, G.M.; Ward, T.J.; O’Donnell, K.; Nicolli, C.P.; Machado, F.J.; Duffeck, M.R.; Alves, K.S.; Tessmann, D.J.; Waalwijk, C.; et al. Fusarium graminearum species complex: A bibliographic analysis and web-accessible database for global mapping of species and trichothecene toxin chemotypes. Phytopathology 2022, 112, 741–751. [Google Scholar] [CrossRef]
- O’Donnell, K.; Ward, T.J.; Aberra, D.; Kistler, H.C.; Aoki, T.; Orwig, N.; Kimura, M.; Bjørnstad, Å.; Klemsdal, S.S. Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fungal. Genet. Biol. 2008, 45, 1514–1522. [Google Scholar] [CrossRef]
- Starkey, D.E.; Ward, T.J.; Aoki, T.; Gale, L.R.; Kistler, H.C.; Geiser, D.M.; Suga, H.; Tóth, B.; Varga, J.; O’donnell, K. Global molecular surveillance reveals novel Fusarium head blight species and trichothecene toxin diversity. Fungal. Genet. Biol. 2007, 44, 1191–1204. [Google Scholar] [CrossRef]
- Yli-Mattila, T.; Gagkaeva, T.; Ward, T.J.; Aoki, T.; Kistler, H.C.; O’Donnell, K. A novel Asian clade within the Fusarium graminearum species complex includes a newly discovered cereal head blight pathogen from the Russian Far East. Mycologia 2009, 101, 841–852. [Google Scholar] [CrossRef]
- Alexander, N.J.; McCormick, S.P.; Waalwijk, C.; van der Lee, T.; Proctor, R.H. The genetic basis for 3-ADON and 15-ADON trichothecene chemotypes in Fusarium. Fungal Genet. Biol. 2011, 48, 485–495. [Google Scholar] [CrossRef]
- Gil-Serna, J.; Vázquez, C.; González-Jaén, M.T.; Patiño, B. Mycotoxins: Toxicology. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C., Tortorello, M.L., Eds.; Academic Press: Amsterdam, The Netherlands, 2014; pp. 1539–1547. [Google Scholar] [CrossRef]
- Merhej, J.; Richard-Forget, F.; Barreau, C. Regulation of trichothecene biosynthesis in Fusarium: Recent advances and new insights. Appl. Microbiol. Biotechnol. 2011, 91, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Talas, F.; McDonald, B.A. Genome-wide analysis of Fusarium graminearum field populations reveals hotspots of recombination. BMC Genom. 2015, 16, 996. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewska, H.; Stępień, Ł.; Waśkiewicz, A.; Beszterda, M.; Góral, T.; Belter, J. Toxigenic Fusarium species infecting wheat heads in Poland. Cent. Eur. J. Biol. 2014, 9, 163–172. [Google Scholar] [CrossRef]
- Duba, A.; Goriewa-Duba, K.; Wachowska, U. Trichothecene genotypes analysis of Fusarium isolates from di-, tetra- and hexaploid wheat. Agronomy 2019, 9, 698. [Google Scholar] [CrossRef]
- Kulik, T.; Molcan, T.; Bilska, K.; Beyer, M.; Pasquali, M.; van Diepeningen, A.; Myszczynski, K. Two distinct Fusarium graminearum populations colonized European wheat in the past two decades. PLoS ONE 2023, 18, e0296302. [Google Scholar] [CrossRef]
- Talas, F.; Parzies, H.K.; Miedaner, T. Diversity in genetic structure and chemotype composition of Fusarium graminearum sensu stricto populations causing wheat head blight in individual fields in Germany. Eur. J. Plant Pathol. 2011, 131, 39–48. [Google Scholar] [CrossRef]
- Spolti, P.; Barros, N.C.; Gomes, L.B.; Dos Santos, J.; Del Ponte, E.M. Phenotypic and pathogenic traits of two species of the Fusarium graminearum complex possessing either 15-ADON or NIV genotype. Eur. J. Plant Pathol. 2012, 133, 621–629. [Google Scholar] [CrossRef]
- Amarasinghe, C.C.; Fernando, W.G.D. Comparative analysis of deoxynivalenol biosynthesis related gene expression among different chemotypes of Fusarium graminearum in spring wheat. Front. Microbiol. 2016, 7, 1229. [Google Scholar] [CrossRef]
- Kelly, A.C.; Ward, T.J. Population genomics of Fusarium graminearum reveals signatures of divergent evolution within a major cereal pathogen. PLoS ONE 2018, 13, e0194616. [Google Scholar] [CrossRef]
- Hao, G.; McCormick, S.; Tiley, H.; Gutiérrez, S.; Yulfo-Soto, G.; Vaughan, M.M.; Ward, T.J. NX trichothecenes are required for Fusarium graminearum infection of wheat. Mol. Plant Microbe. Interact. 2023, 36, 294–304. [Google Scholar] [CrossRef]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, J.; Wang, H.; Yang, X.; Zhang, C.; Zhao, Z.; Wang, J. NX toxins: New threat posed by Fusarium graminearum species complex. Trends Food Sci. Technol. 2022, 119, 79–191. [Google Scholar] [CrossRef]
- Varga, E.; Wiesenberger, G.; Hametner, C.; Ward, T.J.; Dong, Y.; Schöfbeck, D.; McCormick, S.; Broz, K.; Stückler, R.; Schuhmacher, R.; et al. New tricks of an old enemy: Isolates of Fusarium graminearum produce a type A trichothecene mycotoxin. Environ. Microbiol. 2015, 17, 2588–2600. [Google Scholar] [CrossRef] [PubMed]
- Gale, L.R.; Bryant, J.; Calvo, S.; Giese, H.; Katan, T.; O Donnell, K.; Suga, H.; Taga, M.; Usgaard, T.R.; Ward, T.J.; et al. Chromosome complement of the fungal plant pathogen Fusarium graminearum based on genetic and physical mapping and cytological observations. Genetics 2005, 171, 985–1001. [Google Scholar] [CrossRef]
- Gil-Serna, J.; Vázquez, C.; Patiño, B. Genetic regulation of aflatoxin, ochratoxin A, trichothecene, and fumonisin biosynthesis: A review. Int. J. Microbiol. 2020, 23, 89–96. [Google Scholar] [CrossRef]
- Proctor, R.H.; McCormick, S.P.; Alexander, N.J.; Desjardins, A.E. Evidence that a secondary metabolic biosynthetic gene cluster has grown by gene relocation during evolution of the filamentous fungus Fusarium. Mol. Microbiol. 2009, 74, 1128–1142. [Google Scholar] [CrossRef]
- Huang, P.; Yu, X.; Liu, H.; Ding, M.; Wang, Z.; Xu, J.-R.; Jiang, C. Regulation of TRI5 expression and deoxynivalenol biosynthesis by a long non-coding RNA in Fusarium graminearum. Nat. Commun. 2024, 15, 1216. [Google Scholar] [CrossRef]
- Hohn, T.M.; Beremand, P.D. Isolation and nucleotide sequence of a sesquiterpene cyclase gene from the trichothecene-producing fungus Fusarium sporotrichioides. Gene 1989, 79, 131–138. [Google Scholar] [CrossRef]
- Bai, G.H.; Desjardins, A.E.; Plattner, R.D. Deoxynivalenol-nonproducing Fusarium graminearum causes initial infection, but does not cause disease spread in wheat spikes. Mycopathologia 2002, 153, 91–98. [Google Scholar] [CrossRef]
- McCormick, S.P.; Alexander, N.J.; Proctor, R.H. Gang, D., Ed.; Trichothecene triangle: Toxins, genes, and plant disease. In Phytochemicals, Plant Growth, and the Environment; Recent Advances in Phytochemistry; Springer: New York, NY, USA, 2013; Volume 42. [Google Scholar] [CrossRef]
- Liang, J.M.; Xayamongkhon, H.; Broz, K.; Dong, Y.; McCormick, S.P.; Abramova, S.; Ward, T.J.; Ma, Z.H.; Kistler, H.C. Temporal dynamics and population genetic structure of Fusarium graminearum in the upper Midwestern United States. Fungal Genet. Biol. 2014, 73, 83–92. [Google Scholar] [CrossRef]
- Alexander, N.J.; Proctor, R.H.; McCormick, S.P. Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in Fusarium. Toxin Rev. 2009, 28, 198–215. [Google Scholar] [CrossRef]
- Kimura, M.; Tokai, T.; O’Donnell, K.; Ward, T.J.; Fujimura, M.; Hamamoto, H.; Shibata, T.; Yamaguchi, I. The trichothecene biosynthesis gene cluster of Fusarium graminearum F15 contains a limited number of essential pathway genes and expressed non-essential genes. FEBS Lett. 2003, 539, 105–110. [Google Scholar] [CrossRef]
- McCormick, S.P.; Alexander, N.J. Fusarium Tri8 encodes a trichothecene C-3 esterase. Appl. Environ. Microbiol. 2002, 68, 2959–2964. [Google Scholar] [CrossRef]
- Villafana, R.T.; Ramdass, A.C.; Rampersad, S.N. Selection of Fusarium trichothecene toxin genes for molecular detection depends on TRI gene cluster organization and gene function. Toxins 2019, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Wachowska, U.; Sulyok, M.; Wiwart, M.; Suchowilska, E.; Giedrojć, W.; Gontarz, D.; Kandler, W.; Krska, R. Secondary metabolites of pathogenic fungi in Triticum durum grain protected with Debaryomyces hansenii in two different locations in Poland. Agronomy 2023, 13, 721. [Google Scholar] [CrossRef]
- Chełkowski, J.; Gromadzka, K.; Stępień, Ł.; Lenc, L.; Kostecki, M.; Berthiller, F. Fusarium species, zearalenone and deoxynivalenol content in preharvest scabby wheat heads from Poland. World Mycotoxin J. 2012, 5, 133–141. [Google Scholar] [CrossRef]
- Góral, T.; Łukanowski, A.; Małuszyńska, E.; Stuper-Szablewska, K.; Buśko, M.; Perkowski, J. Performance of winter wheat cultivars grown organically and conventionally with focus on Fusarium head blight and Fusarium trichothecene toxins. Microorganisms 2019, 7, 439. [Google Scholar] [CrossRef]
- Lenc, L.; Czecholiński, G.; Wyczling, D.; Turów, T.; Kaźmierczak, A. Fusarium head blight (FHB) and Fusarium spp. on grain of spring wheat cultivars grown in Poland. J. Plant Prot. Res. 2015, 55, 266–277. [Google Scholar] [CrossRef]
- Sadowski, C.; Lenc, L.; Kuś, J. Fusarium head blight and Fusarium spp. on grain of winter wheat, a mixture of cultivars and spelt grown in organic system. J. Res. App. Agric. Eng. 2010, 55, 79–83. [Google Scholar]
- Gorczyca, A.; Oleksy, A.; Gala-Czekaj, D.; Urbaniak, M.; Laskowska, M.; Waśkiewicz, A.; Stępień, Ł. Fusarium head blight incidence and mycotoxin accumulation in three durum wheat cultivars in relation to sowing date and density. Sci. Nat. 2018, 105, 2. [Google Scholar] [CrossRef]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Mañes, J. Fusarium species, chemotype characterisation and trichothecene contamination of durum and soft wheat in an area of central Italy. J. Sci. Food Agric. 2015, 95, 540–551. [Google Scholar] [CrossRef]
- Polišenská, I.; Jirsa, O.; Salava, J.; Sedláčková, I.; Frydrych, J. Fusarium mycotoxin content and Fusarium species presence in Czech organic and conventional wheat. World Mycotoxin J. 2021, 14, 201–211. [Google Scholar] [CrossRef]
- Suproniene, S.; Sakalauskas, S.; Stumbriene, K.; Zvirdauskiene, R.; Svegzda, P. Variances in trichothecene chemotype distribution in Lithuanian wheat grain and within pure culture Fusarium graminearum isolated from the same grain samples. Eur. J. Plant Pathol. 2016, 144, 371–381. [Google Scholar] [CrossRef]
- Sakalauskas, S.; Stumbriene, K.; Suproniene, S.; Svegzda, P. Changes in Fusarium link species composition from Lithuanian wheat grain in years 2005–2007 to 2011–2013. Rural Sustain. Res. 2014, 32, 45–50. [Google Scholar] [CrossRef]
- Tima, H.; Brückner, A.; Mohácsi-Farkas, C.; Kiskó, G. Fusarium mycotoxins in cereals harvested from Hungarian fields. Food Addit. Contam. Part B Surveill. 2016, 9, 127–131. [Google Scholar] [CrossRef]
- Commission Regulation (EU) 2024/1022 of 8 April 2024 Amending Regulation (EU) 2023/915 as Regards Maximum Levels of Deoxynivalenol in Food. Available online: https://eur-lex.europa.eu/eli/reg/2024/1022/oj/eng (accessed on 12 October 2024).
- Mesterhazy, A. What Is Fusarium Head Blight (FHB) Resistance and what are its food safety risks in wheat? Problems and solutions—A review. Toxins 2024, 16, 31. [Google Scholar] [CrossRef]
- Amarasinghe, C.C.; Simsek, S.; Brûlé-Babel, A.; Fernando, W.D. Analysis of deoxynivalenol and deoxynivalenol-3-glucosides content in Canadian spring wheat cultivars inoculated with Fusarium graminearum. Food Addit. Contamin. Part A 2016, 33, 1254–1264. [Google Scholar] [CrossRef]
- Wenda-Piesik, A.; Lemańczyk, G.; Twarużek, M.; Błajet-Kosicka, A.; Kazek, M.; Grajewski, J. Fusarium head blight incidence and detection of Fusarium toxins in wheat in relation to agronomic factors. Eur. J. Plant Pathol. 2017, 149, 515–531. [Google Scholar] [CrossRef]
- Baturo-Cieśniewska, A.; Lukanowski, A.; Kolenda, M. Effect of fungicide application on wheat head blight, occurrence of Fusarium spp. and mycotoxin production. Plant Breed. Seed Sci. 2011, 63, 29–38. [Google Scholar] [CrossRef]
- Kulik, T.; Łojko, M.; Jestoi, M.; Perkowski, J. Sublethal concentrations of azoles induce tri transcript levels and trichothecene production in Fusarium graminearum. FEMS Microbiol. Lett. 2012, 335, 58–67. [Google Scholar] [CrossRef]
- Duan, Y.; Lu, F.; Zhou, Z.; Zhao, H.; Zhang, J.; Mao, Y.; Li, M.; Wang, J.; Zhou, M. Quinone outside inhibitors affect DON biosynthesis, mitochondrial structure and toxisome formation in Fusarium graminearum. J. Hazard. Mater. 2020, 398, 122908. [Google Scholar] [CrossRef]
- Klocke, B.; Sommerfeldt, N.; Wagner, C.; Schwarz, J.; Baumecker, M.; Ellmer, F.; Jacobi, A.; Matschiner, K.; Petersen, J.; Wehling, P.; et al. Disease threshold-based fungicide applications: Potential of multi-disease resistance in winter wheat cultivars in Germany. Eur. J. Plant Pathol. 2023, 165, 363–383. [Google Scholar] [CrossRef]
- Parliament European Commission. Directive 2009/128/EC of the European Parliament and of the Council of 21 October 2009 establishing a framework for Community action to achieve the sustainable use of pesticides. Off J Eur Union. 2009, L309, 71–86. [Google Scholar]
- Ministry of Agriculture and Rural Development. Republic of Poland. 2024. Available online: https://www.gov.pl/web/piorin/wyszukiwarka-sor-mrirw (accessed on 1 December 2024).
- FRAC. Fungicide Resistance Action Committee. 2024. Available online: https://www.frac.info (accessed on 10 October 2024).
- Anderson, N.R.; Freije, A.N.; Bergstrom, G.C.; Bradley, C.A.; Cowger, C.; Faske, T.; Hollier, C.; Kleczewski, N.; Padgett, G.B.; Paul, P.; et al. Sensitivity of Fusarium graminearum to metconazole and tebuconazole fungicides before and after widespread use in wheat in the United States. Plant Health Prog. 2020, 21, 85–90. [Google Scholar] [CrossRef]
- Miao, J.; Li, Y.; Hu, S.; Li, G.; Gao, X.; Dai, T.; Liu, X. Resistance risk, resistance mechanism and the effect on DON production of a new SDHI fungicide cyclobutrifluram in Fusarium graminearum. Pestic. Biochem. Physiol. 2024, 199, 105795. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Du, J.; Chi, M.; Sun, X.; Liang, W.; Huang, J.; Li, B. The Y137H mutation in the cytochrome P450 FgCYP51B protein confers reduced sensitivity to tebuconazole in Fusarium graminearum. Pest Manag. Sci. 2018, 74, 1472–1477. [Google Scholar] [CrossRef]
- Rekanović, E.; Mihajlović, M.; Potočnik, I. In vitro sensitivity of Fusarium graminearum (Schwabe) to difenoconazole, prothioconazole and thiophanate-mythyl. Pestic. Fitomed. 2010, 25, 325–333. [Google Scholar] [CrossRef]
- Duffeck, M.R.; dos Santos Alves, K.; Machado, F.J.; Esker, P.D.; Del Ponte, E.M. Modeling yield losses and fungicide profitability for managing Fusarium head blight in Brazilian spring wheat. Phytopathology 2020, 110, 370–378. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, J.; Shao, J.; Yin, Y.; Ma, Z. Gene transcription profiling of Fusarium graminearum treated with an azole fungicide tebuconazole. Appl. Microbiol. Biotechnol. 2020, 85, 1105–1114. [Google Scholar] [CrossRef]
- Qi, P.F.; Zhang, Y.Z.; Liu, C.H.; Zhu, J.; Chen, Q.; Guo, Z.R.; Wang, Y.; Xu, B.J.; Zheng, T.; Jiang, Y.F.; et al. Fusarium graminearum ATP-Binding cassette transporter gene FgABCC9 is required for its transportation of salicylic acid, fungicide resistance, mycelial growth and pathogenicity towards wheat. Int. J. Mol. Sci. 2018, 19, 2351. [Google Scholar] [CrossRef]
- Ivić, D.; Sever, Z.; Kuzmanovska, B. In vitro Sensitivity of Fusarium graminearum, f. avenaceum and F. verticillioides to carbendazim, tebuconazole, flutriafol, metconazole and prochloraz. Pestic. Fitomed. 2011, 26, 35–42. [Google Scholar] [CrossRef]
- Matelionienė, N.; Žvirdauskienė, R.; Kadžienė, G.; Zavtrikovienė, E.; Supronienė, S. In Vitro sensitivity test of Fusarium species from weeds and non-gramineous plants to triazole fungicides. Pathogens 2024, 13, 160. [Google Scholar] [CrossRef] [PubMed]
- Machado, F.J.; Silva, C.N.; Paiva, G.F.; Feksa, H.R.; Tessmann, D.J.; Mizubuti, E.S.G.; Del Ponte, E.M. Sensitivity to tebuconazole and carbendazim in Fusarium graminearum species complex populations causing wheat head blight in southern Brazil. Trop. Plant Pathol. 2024, 49, 157–167. [Google Scholar] [CrossRef]
- Andrade, S.M.; Augusti, G.R.; Paiva, G.F.; Feksa, H.R.; Tessmann, D.J.; Machado, F.J.; Mizubuti, E.S.G.; Del Ponte, E.M. Phenotypic and molecular characterization of the resistance to azoxystrobin and pyraclostrobin in Fusarium graminearum populations from Brazil. Plant Pathol. 2022, 71, 1152–1163. [Google Scholar] [CrossRef]
- Luan, S.; Chen, Y.; Wang, X.; Yan, D.; Xu, J.; Cui, H.; Huang, Q. Synergy of cystamine and pyraclostrobin against Fusarium graminearum involves membrane permeability mitigation and autophagy enhancement. Pestic. Biochem. Physiol. 2022, 188, 105287. [Google Scholar] [CrossRef]
- Xu, C.; Li, M.; Zhou, Z.; Li, J.; Chen, D.; Duan, Y.; Zhou, M. Impact of five succinate dehydrogenase inhibitors on don biosynthesis of Fusarium asiaticum, causing Fusarium head blight in wheat. Toxins 2019, 11, 272. [Google Scholar] [CrossRef]
- Fernández-Ortuño, D.; Torés, J.A.; de Vicente, A.; Pérez-García, A. Field resistance to QoI fungicides in Podosphaera fusca is not supported by typical mutations in the mitochondrial cytochrome b gene. Pest. Manag. Sci. 2008, 64, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Qiu, H.; Gao, X.; Li, Y.; Zheng, X.; Cai, Y.; Sheng, G.; Shen, Y.; Wang, J.; Zhou, M.; et al. Resistance risk and molecular mechanism of tomato wilt pathogen Fusarium oxysporum f. sp. lycopersici to pyraclostrobin. J. Agric. Food Chem. 2024, 72, 3998–4007. [Google Scholar] [CrossRef]
- Dubos, T.; Pasquali, M.; Pogoda, F.; Hoffmann, L.; Beyer, M. Evidence for natural resistance towards trifloxystrobin in Fusarium graminearum. Eur. J. Plant Pathol. 2022, 130, 239–248. [Google Scholar] [CrossRef]
- Tsukamoto, M.; Nakamura, T.; Kimura, H.; Nakayama, H. Synthesis and application of trifluoromethyl pyridines as a key structural motif in active agrochemical and pharmaceutical ingredients. J. Pestic. Sci. 2021, 46, 125–142. [Google Scholar] [CrossRef]
- Syngenta. 2024. Available online: https://www.syngenta.com/ (accessed on 14 October 2024).
- Shao, W.; Wang, J.; Wang, H.; Wen, Z.; Liu, C.; Zhang, Y.; Zhao, Y.; Ma, Z. Fusarium graminearum FgSdhC1 point mutation A78V confers resistance to the succinate dehydrogenase inhibitor pydiflumetofen. Pest Manag. Sci. 2022, 78, 1780–1788. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Chu, Y.; Wen, H.; Li, B.; Dong, X.; Sun, M.; Meng, D.; Wang, M.; Gao, T.; Duan, J. Stereoselective bioactivity, toxicity and degradation of novel fungicide sedaxane with four enantiomers under rice-wheat rotation mode. Ecotoxic. Environ. Saf. 2022, 241, 113784. [Google Scholar] [CrossRef] [PubMed]
- Risoli, S.; Cotrozzi, L.; Pisuttu, C.; Nali, C. Biocontrol agents of fusarium head blight in wheat: A meta-analytic approach to elucidate their strengths and weaknesses. Phytopathology 2024, 114, 521–537. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.A.; Pernica, M.; Yap, J.; Belakova, S.; Vaculova, K.; Branyik, T. Biocontrol effect of Pythium oligandrum on artificial Fusarium culmorum infection during malting of wheat. J. Cereal Sci. 2021, 100, 103258. [Google Scholar] [CrossRef]
- Pellan, L.; Dieye, C.A.T.; Durand, N.; Fontana, A.; Schorr-Galindo, S.; Strub, C. Biocontrol agents reduce progression and mycotoxin production of Fusarium graminearum in spikelets and straws of wheat. Toxins 2021, 13, 597. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Country | Year | Plant Species | Fusarium graminearum | Fusarium avenaceum | Fusarium poae | Fusarium sporotrichioides | Fusarium tricinctum | Fusarium culmorum | Deoxynivalenol | Nivalenol | Zearalenone | Moniliformin | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Percentage of Colonized Kernels | μg kg−1 | ||||||||||||
| Poland | 2007 | Triticum aestivum | 1 | 1–8.8 | 4.0–14.0 | 1–3.0 | 0.8–5.8 | 0 | nb | nb | nb | nb | [63] |
| 2009 | 6.2–38.5 | 4.4–71.4 | 0 | 0 | 0.2–4.00 | 7.2–81.2 | 2040–76,700 | nb | 20–680 | 70–120 | [36] | ||
| 2009 | 21.3–55.1 | 15.8–20.5 | 0 | 0 | 0 | 5.4–42.9 | 4010–69,150 | nb | 4–490 | nb | [60] | ||
| 2011–2013 | 1–4.5 | 2.5–11.0 | 1.0–8.0 | 0.3–2.8 | 1.0–6.0 | 1.10.0 | nb | nb | nb | nb | [62] | ||
| 2014 | 0.7–79.5 * | 0–106.7 * | 9.4–132.4 * | 0–113.3 * | 0 | 0–346.2 * | 5.8–444.4 | 0–19.0 | 0 | 0 | [61] | ||
| 2015 | 1.5–4.9 | 0 | 69–76 | 0 | 0 | 16–26 | 106–1099 | 25–453 | nb | nb | [7] | ||
| 2016–2017 | 46.07–100 | 3.03–45.45 | 9.09 | 0 | 0 | 1.75–13.33 | nb | nb | nb | nb | [13] | ||
| 2011–2013 | Triticum durum | 0 | 8 | 2.0 | 0 | 2.0 | 9.0 | nb | nb | nb | nb | [62] | |
| 2012 | 12 | 24 | 24 | 0 | 12 | 4 | 254–1030 | <LOD | 2.0–27 | <LOD | [64] | ||
| 2013 | 48 | 39 | 0 | 5 | 1 | 5 | 10–10,879 | <LOD | 41–307 | 9–42 | |||
| 2014 | 43 | 36 | 1 | 13 | 0 | 3 | 424–3988 | 88–155 | 2.8–21 | 40–580 | |||
| 2018 | 2.1–9.0 | 3 | 0–24 | 0 | 0 | 4.0–20.1 | 72–2083 | 13.9–318.9 | 0.95–64.4 | 0 | [59] | ||
| Italy | 2009 | Triticum durum | 27 | 22 | 0 | 0 | 0 | 13 | 10.7–597.3 | 41.0–1648.6 | nb | nb | [65] |
| 2010 | 69 | 6 | 1 | 1 | 0 | 7 | 32.9–512.3 | 135.6–965.4 | nb | nb | |||
| 2009 | Triticum aestivum | 16 | 37 | 16 | 0 | 0 | 0 | 9.7–198.1 | 122–963.6 | nb | nb | ||
| 2010 | 60 | 15 | 3 | 0 | 0 | 4 | 26.3–261.3 | 21.1–621.5 | nb | nb | |||
| Czech Republic | 2015 | Triticum aestivum | 38 | 0 | 9 | 0 | 0 | 8 | 20 | nb | 2 | nb | [66] |
| 2016 | 77 | 13 | 53 | 0 | 0 | 2 | 148 | nb | 6 | nb | |||
| 2017 | 21 | 6 | 75 | 32 | 0 | 0 | 75 | nb | 2 | nb | |||
| Lithuania | 2013–2014 | Triticum aestivum | nb | nb | nb | nb | nb | nb | nb | nb | nb | nb | [67] |
| 2005–2007 | 0 | 17.6 | 4.4 | 4.47 | 0.6 | 5.7 | nb | nb | nb | nb | [68] | ||
| 2011–2013 | 19.1 | 18.7 | 4.9 | 19.1 | 1 | 19.4 | nb | nb | nb | nb | |||
| Germany | 2006–2007 | Triticum aestivum | 64.9 | 0 | 3 | 0 | 3 | 26.1 | nb | nb | nb | nb | [39] |
| Hungary | 2015 | Triticum aestivum | nb | nb | nb | nb | nb | nb | 230–1880 | nb | 50–98 | nb | [69] |
| MOA | Group Name | Chemical or Biological Group | Active Ingredient | Target Site and Code | Number of Fungicides |
|---|---|---|---|---|---|
| C. respiration | QoI-fungicides, (quinone outside inhibitors) | Methoxy-acrylates | azoxystrobin | C3 complex III: cytochrome bc1 (ubiquinol oxidase) at Qo site (cyt b gene) | **** |
| fluoxastrobin | * | ||||
| Oxime acetates | trifloxystrobin | * | |||
| SDHI-fungicides (succinate-dehydrogenase inhibitors) | Pyrazole-4-carboxamides | bixafen | C2 complex II: succinate-dehydrogenase | * | |
| sedaxane | * | ||||
| benzovindiflupyr | * | ||||
| Pyridinyl-ethyl-benzamides | fluopyram | * | |||
| Pyridine-carboxamides | boscalid | * | |||
| G: sterol biosynthesis in membranes | DMI-fungicides (demethylation inhibitors) | Triazolinthiones | prothioconazole | G1 C14-demethylase in sterol biosynthesis (erg11/cyp51) | **** |
| Triazoles | metconazole | *** | |||
| (SBI: Class I) | tebuconazole | *** | |||
| bromuconazole | ** | ||||
| Microbial | Pythium oligandrum | * | |||
| Fungicide | Country of Origin | Year of Isolation | Number of Isolates | Mean EC50 (μg/mL) | EC min (μg/mL) | EC max (μg/mL) | Reference |
|---|---|---|---|---|---|---|---|
| Triazoles | |||||||
| Propiconazole | Germany | 2008 | 197 | 22.2 | 5.4 | 62.2 | [35] |
| Tebuconazole | Brazil | 2011–2020 | 22 | 0.57 | 0.06 | 3.15 | [90] |
| 11 | 0.05 | 0.00 | 0.19 | ||||
| Metconazole | Lithuania | 2021–2022 | 13 | 0.789 | 0.010 | 2.967 | [89] |
| Tebuconazole | 10.877 | 1.197 | 25.623 | ||||
| Prothioconazole | 8.751 | 2.201 | 22.909 | ||||
| Tebuconazole | Serbia | 13 | 1.282 | 0.910 | 2.570 | [88] | |
| Metconazole | 2009 | 0.479 | 0.090 | 1.660 | |||
| Carbendazim | 0.687 | 0.390 | 1.410 | ||||
| Strobilurin | |||||||
| Azoxystrobin | China | 2017 | 32 | 0.966 | 0.274 | 1.240 | [76] |
| Fluoxastrobin | 0.841 | 0.268 | 1.775 | ||||
| Azoxystrobin | Brazil | 2007–2020 | 225 | 28.06 | 0.260 | 329.7 | [91] |
| Pyraclostrobin | 0.330 | 0.030 | 1.130 | ||||
| Pyraclostrobin | China | nd * | 7.083 | 4.185 | 13.399 | [92] | |
| SDHI | |||||||
| Boscalid | China | 2008–2020 | 6 | 2.265 | 1.280 | 2.410 | [93] |
| Benzovindiflupyr | 2.568 | 2.030 | 2.980 | ||||
| Fluopyram | 0.597 | 0.520 | 0.690 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giedrojć, W.; Pluskota, W.E.; Wachowska, U. Fusarium graminearum in Wheat—Management Strategies in Central Europe. Pathogens 2025, 14, 265. https://doi.org/10.3390/pathogens14030265
Giedrojć W, Pluskota WE, Wachowska U. Fusarium graminearum in Wheat—Management Strategies in Central Europe. Pathogens. 2025; 14(3):265. https://doi.org/10.3390/pathogens14030265
Chicago/Turabian StyleGiedrojć, Weronika, Wioletta E. Pluskota, and Urszula Wachowska. 2025. "Fusarium graminearum in Wheat—Management Strategies in Central Europe" Pathogens 14, no. 3: 265. https://doi.org/10.3390/pathogens14030265
APA StyleGiedrojć, W., Pluskota, W. E., & Wachowska, U. (2025). Fusarium graminearum in Wheat—Management Strategies in Central Europe. Pathogens, 14(3), 265. https://doi.org/10.3390/pathogens14030265