
Exploring Mosquito Excreta as an Alternative Sample Type for Improving Arbovirus Surveillance in Australia
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Dengue Virus Serotype 2 Culture and Isolation
2.2. Breeding, Raising, and Infecting Aedes aegypti
2.3. RNA Extraction, cDNA Generation, and qPCR
2.4. Detection of DENV2 in Mosquito Excreta Using qPCR
2.5. Mosquito Excreta Collection into Liquid Substrate
2.6. Concentration and Detection of DENV2 in Mosquito Excreta Using Magnetic Beads
3. Results
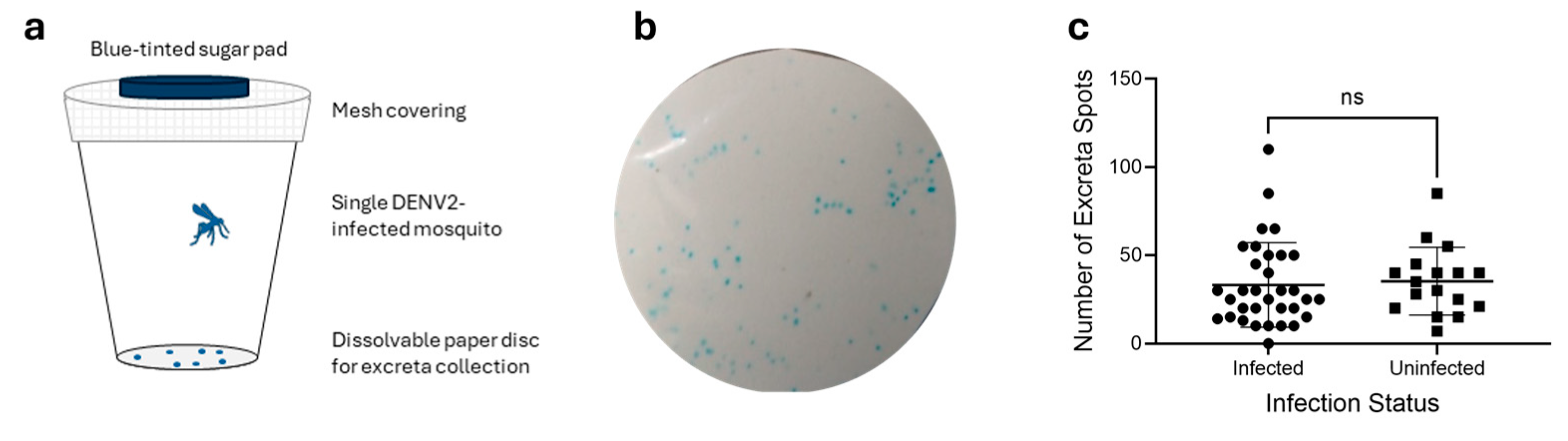
3.1. Mosquito Excretion Patterns Are Not Correlated with Infection Status
3.2. DENV2 Can Be Detected from a Single Mosquito Excreta Spot
3.3. DENV2 Detection in Excreta Is Dependent on Mosquito Viral Load
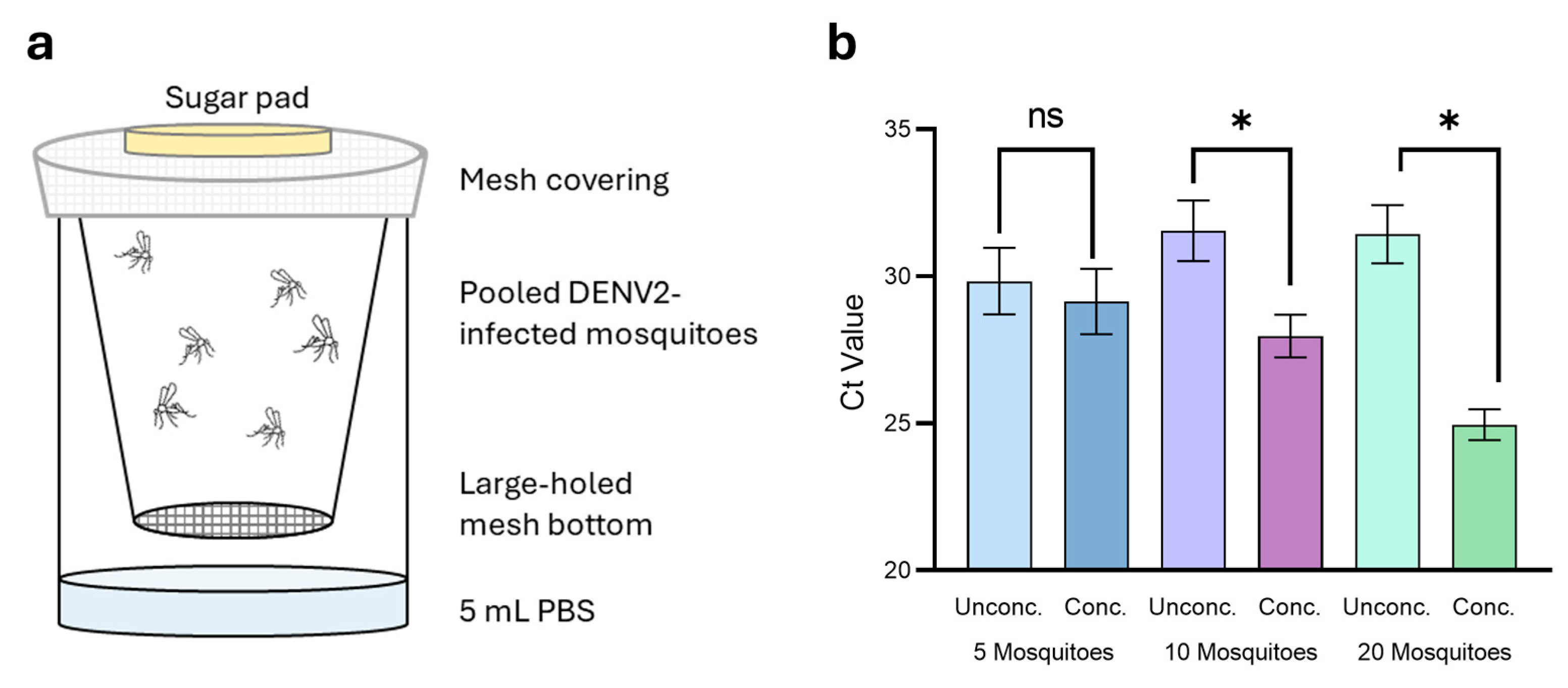
3.4. Magnetic Concentration Expands Detection Capabilities
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Arbovirus Initiative. 2024. Available online: https://www.who.int/initiatives/global-arbovirus-initiative (accessed on 16 October 2024).
- World Health Organization. Vector-Borne Diseases. 2024. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 16 October 2024).
- Roiz, D.; Pontifes, P.A.; Jourdain, F.; Diagne, C.; Leroy, B.; Vaissière, A.-C.; Tolsá-García, M.J.; Salles, J.-M.; Simard, F.; Courchamp, F. The rising global economic costs of invasive Aedes mosquitoes and Aedes-borne diseases. Sci. Total Environ. 2024, 933, 173054. [Google Scholar] [CrossRef] [PubMed]
- Van Den Hurk, A.F.; Hall-Mendelin, S.; Johansen, C.A.; Warrilow, D.; Ritchie, S.A. Evolution of mosquito-based arbovirus surveillance systems in Australia. J. Biomed. Biotechnol. 2012, 2012, 325659. [Google Scholar] [CrossRef]
- Ramírez, A.L.; Van Den Hurk, A.F.; Meyer, D.B.; Ritchie, S.A. Searching for the proverbial needle in a haystack: Advances in mosquito-borne arbovirus surveillance. Parasites Vectors 2018, 11, 320. [Google Scholar] [CrossRef] [PubMed]
- Government of Wester Australia Department of Health. Arbovirus Surveillance Program. 2024. Available online: https://www.health.wa.gov.au/articles/a_e/arbovirus-surveillance-program#:~:text=Regular%20monitoring%20of%20MVEV%20and,northern%20mosquito%20surveillance (accessed on 16 October 2024).
- Victoria Health. Mosquito-Borne Diseases. 2024. Available online: https://www.health.vic.gov.au/infectious-diseases/mosquito-borne-diseases (accessed on 16 October 2024).
- South Australia Health. Arbovirus and Mosquito Monitoring Reports. 2024. Available online: https://www.sahealth.sa.gov.au/wps/wcm/connect/public+content/sa+health+internet/public+health/pest+management/mosquitoes/arbovirus+and+mosquito+monitoring+reports#Sentinelchickensurveillance (accessed on 16 October 2024).
- New South Wales Health. Mosquito-Borne Disease Surveillance. 2023. Available online: https://www.health.nsw.gov.au/Infectious/mosquito-borne/Pages/surveillance.aspx#mosquitomonitoring (accessed on 16 October 2024).
- Knope, K.; Doggett, S.L.; Jansen, C.C.; Johansen, C.A.; Kurucz, N.; Feldman, R.; Lynch, S.E.; Hobby, M.P.; Sly, A.; Jardine, A.; et al. Arboviral diseases and malaria in Australia, 2014–15: Annual report of the National Arbovirus and Malaria Advisory Committee. Commun. Dis. Intell. 2019, 43, 1–69. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.R.; Webb, C.E.; Higgs, S.; Van Den Hurk, A.F. Japanese Encephalitis Virus Emergence in Australia: Public Health Importance and Implications for Future Surveillance. Vector-Borne Zoonotic Dis. 2022, 22, 529–534. [Google Scholar] [CrossRef]
- Kurucz, N.; McMahon, J.L.; Warchot, A.; Hewitson, G.; Barcelon, J.; Moore, F.; Moran, J.; Harrison, J.J.; Colmant, A.M.G.; Staunton, K.M.; et al. Nucleic Acid Preservation Card Surveillance Is Effective for Monitoring Arbovirus Transmission on Crocodile Farms and Provides a One Health Benefit to Northern Australia. Viruses 2022, 14, 1342. [Google Scholar] [CrossRef] [PubMed]
- Kurucz, N.; Minney-Smith, C.A.; Johansen, C.A. Arbovirus surveillance using FTATM cards in modified CO2-baited encephalitis virus surveillance traps in the Northern Territory, Australia. J. Vector Ecol. 2019, 44, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Hall-Mendelin, S.; Ritchie, S.A.; Johansen, C.A.; Zborowski, P.; Cortis, G.; Dandridge, S.; Hall, R.A.; van den Hurk, A.F. Exploiting mosquito sugar feeding to detect mosquito-borne pathogens. Proc. Natl. Acad. Sci. USA 2010, 107, 11255–11259. [Google Scholar] [CrossRef] [PubMed]
- van den Hurk, A.F.; Hall-Mendelin, S.; Townsend, M.; Kurucz, N.; Edwards, J.; Ehlers, G.; Rodwell, C.; Moore, F.A.; McMahon, J.L.; Northill, J.A.; et al. Applications of a sugar-based surveillance system to track arboviruses in wild mosquito populations. Vector-Borne Zoonotic Dis. 2014, 14, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Flies, E.J.; Toi, C.; Weinstein, P.; Doggett, S.L.; Williams, C.R. Converting Mosquito Surveillance to Arbovirus Surveillance with Honey-Baited Nucleic Acid Preservation Cards. Vector-Borne Zoonotic Dis. 2015, 15, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.J.; Kerlin, T.; Hall-Mendelin, S.; van den Hurk, A.F.; Cortis, G.; Doggett, S.L.; Toi, C.; Fall, K.; McMahon, J.L.; Townsend, M.; et al. Development and field evaluation of the sentinel mosquito arbovirus capture kit (SMACK). Parasit Vectors 2015, 8, 509. [Google Scholar] [CrossRef]
- Devine, T.L.; Venard, C.E.; Myser, W.C. Measurement of salivation by Aedes aegypti (L.) feeding on a living host. J. Insect Physiol. 1965, 11, 347–353. [Google Scholar] [CrossRef]
- Fontaine, A.; Jiolle, D.; Moltini-Conclois, I.; Lequime, S.; Lambrechts, L. Excretion of dengue virus RNA by Aedes aegypti allows non-destructive monitoring of viral dissemination in individual mosquitoes. Sci. Rep. 2016, 6, 24885. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.C.; Cime-Castillo, J.; Argotte-Ramos, R.; Vargas, V.; Correa-Morales, F.; Sánchez-Tejeda, G.; Lanz-Mendoza, H. Detection of NS1 protein from dengue virus in excreta and homogenates of wild-caught Aedes aegypti mosquitoes using monoclonal antibodies. Pathog. Dis. 2022, 80, ftac002. [Google Scholar] [CrossRef]
- Ramírez, A.L.; Hall-Mendelin, S.; Doggett, S.L.; Hewitson, G.R.; McMahon, J.L.; Ritchie, S.A.; van den Hurk, A.F. Mosquito excreta: A sample type with many potential applications for the investigation of Ross River virus and West Nile virus ecology. PLoS Negl. Trop. Dis. 2018, 12, e0006771. [Google Scholar] [CrossRef] [PubMed]
- Körsten, C.; Vasić, A.; Al-Hosary, A.A.; Tews, B.A.; Răileanu, C.; Silaghi, C.; Schäfer, M. Excretion Dynamics of Arboviruses in Mosquitoes and the Potential Use in Vector Competence Studies and Arbovirus Surveillance. Trop. Med. Infect. Dis. 2023, 8, 410. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.B.; Ramirez, A.L.; Van Den Hurk, A.F.; Kurucz, N.; Ritchie, S.A. Development and Field Evaluation of a System to Collect Mosquito Excreta for the Detection of Arboviruses. J. Med. Entomol. 2019, 56, 1116–1121. [Google Scholar] [CrossRef]
- Gooding, R.H.; Cheung, A.C.; Rolseth, B.M. The digestive processes of haematophagous insects: III. inhibition of trypsin by honey and the possible functions of the Oesophageal diverticula of mosquitoes (diptera). Can. Entomol. 1973, 105, 433–436. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Watts, D.M.; Burke, D.S.; Harrison, B.A.; Whitmire, R.E.; Nisalak, A. Effect of temperature on the vector efficiency of aedes aegypti for dengue 2 virus. Am. I. Trop. Med. Hyg. 1987, 36, 143–152. [Google Scholar] [CrossRef]
- Sakuma, C.; Kanuka, H. A simple and affordable method for estimating the fluid volume a mosquito sucks using food dyes. Trop. Med. Health 2021, 49, 13. [Google Scholar] [CrossRef] [PubMed]
- Novelo, M.; Hall, M.D.; Pak, D.; Young, P.R.; Holmes, E.C.; McGraw, E.A. Intra-host growth kinetics of dengue virus in the mosquito Aedes aegypti. PLoS Pathog. 2019, 15, e1008218. [Google Scholar] [CrossRef] [PubMed]
- Munira, S.; Yuki, R.; Saito, Y.; Nakayama, Y. ALK Inhibitors-Induced M Phase Delay Contributes to the Suppression of Cell Proliferation. Cancers 2020, 12, 1054. [Google Scholar] [CrossRef] [PubMed]
- Reijneveld, J.F.; Ocampo, T.A.; Shahine, A.; Gully, B.S.; Vantourout, P.; Hayday, A.C.; Rossjohn, J.; Moody, D.B.; Van Rhijn, I. Human γδ T cells recognize CD1b by two distinct mechanisms. Proc. Natl. Acad. Sci. USA 2020, 117, 22944–22952. [Google Scholar] [CrossRef]
- Lee, H.-N.; Manangeeswaran, M.; Lewkowicz, A.P.; Engel, K.; Chowdhury, M.; Garige, M.; Eckhaus, M.A.; Sourbier, C.; Ireland, D.D.; Verthelyi, D. NK cells require immune checkpoint receptor LILRB4/gp49B to control neurotropic Zika virus infections in mice. JCI Insight 2022, 7, e151420. [Google Scholar] [CrossRef] [PubMed]
- Galasso, B.; Sharma, S.; Knollmann-Ritschel, B.; Sharma, A. Magnetic bead purification of enveloped alphavirus and flavivirus. MethodsX 2021, 8, 101549. [Google Scholar] [CrossRef]
- OZ Biosciences. Viral Applications: Mag4C-Lv. 2021. Available online: www.ozbiosciences.com (accessed on 16 October 2024).
- Mukhopadhyay, S.; Kuhn, R.J.; Rossmann, M.G. A structural perspective of the Flavivirus life cycle. Nat. Rev. Microbiol. 2005, 3, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Montgomery, B.L.; Adamczyk, R.; Ehlers, G.; van den Hurk, A.F.; Warrilow, D. A LAMP-based colorimetric assay to expedite field surveillance of the invasive mosquito species aedes aegypti and aedes albopictus. PLoS Negl. Trop. Dis. 2020, 14, e0008130. [Google Scholar] [CrossRef]
- Gaber, M.; Ahmad, A.A.; El-Kady, A.M.; Tolba, M.; Suzuki, Y.; Mohammed, S.M.; Elossily, N.A. Dengue fever as a reemerging disease in upper Egypt: Diagnosis, vector surveillance and genetic diversity using RT-LAMP assay. PLoS ONE 2022, 17, e0265760. [Google Scholar] [CrossRef] [PubMed]
- Khodaparast, M.; Sharley, D.; Best, N.; Marshall, S.; Beddoe, T. In-field LAMP assay for rapid detection of human faecal contamination in environmental water. Environ. Sci. 2022, 8, 2641–2651. [Google Scholar] [CrossRef]
- Amaral, C.; Antunes, W.; Moe, E.; Duarte, A.G.; Lima, L.M.P.; Santos, C.; Gomes, I.L.; Afonso, G.S.; Vieira, R.; Teles, H.S.S.; et al. A molecular test based on RT-LAMP for rapid, sensitive and inexpensive colorimetric detection of SARS-CoV-2 in clinical samples. Sci. Rep. 2021, 11, 16430. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Kaur, S.; Kayabasi, A.; Ranjbaran, M.; Rath, I.; Benschikovski, I.; Raut, B.; Ra, K.; Rafiq, N.; Verma, M.S. A portable, easy-to-use paper-based biosensor for rapid in-field detection of fecal contamination on fresh produce farms. Biosens. Bioelectron. 2024, 259, 116374. [Google Scholar] [CrossRef] [PubMed]
- Batovska, J.; Lynch, S.E.; Rodoni, B.C.; Sawbridge, T.I.; Cogan, N.O. Metagenomic arbovirus detection using MinION nanopore sequencing. J. Virol. Methods 2017, 249, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Inglis, T.J.J.; Bradbury, R.S.; McInnes, R.L.; Frances, S.P.; Merritt, A.J.; Levy, A.; Nicholson, J.; Neville, P.J.; Lindsay, M.; Smith, D.W. Deployable molecular detection of arboviruses in the Australian outback. Am. J. Trop. Med. Hyg. 2016, 95, 633–638. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Mosquito ID | Positive | Equivocal | Negative | Average Ct ± SEM | Positive Ct Range |
|---|---|---|---|---|---|
| Mosquito 5 (Ct = 17.03) | 10 | 0 | 0 | 30.29 ± 0.80 | 27.45–34.79 |
| Mosquito 44 (Ct = 17.21) | 6 | 0 | 4 | 32.39 ± 1.08 | 27.10–34.22 |
| Mosquito 42 (Ct = 17.59) | 2 | 3 | 5 | 33.25 ± 01.19 | 31.57–34.94 |
| Mosquito 1 (Ct = 17.62) | 1 | 0 | 9 | 35.30 ± 0 | 35.30 |
| Mosquito 28 (Ct = 17.72) | 7 | 3 | 0 | 32.44 ± 0.78 | 28.99–35.39 |
| Mosquito ID | No. Spots per Pool | Positive Pools | Equivocal Pools | Negative Pools | Average Ct ± SEM | Positive Ct Range |
|---|---|---|---|---|---|---|
| Mosquito 4 (Ct = 16.72) | 5 10 | 3 0 | 0 1 | 0 2 | 30.96 ± 1.02 ND | 29.15–32.70 ND |
| Mosquito 21 (Ct = 17.52) | 5 10 | 1 3 | 1 0 | 1 0 | 34.28 ± 0 31.15 ± 0.12 | 34.28 30.91–31.11 |
| Mosquito 19 (Ct = 17.87) | 5 10 | 3 3 | 0 0 | 0 0 | 29.25 ± 1.74 29.54 ± 1.49 | 26.31–32.34 26.78–31.88 |
| Mosquito 32 (Ct = 19.34) | 5 10 | 1 0 | 0 2 | 2 1 | 31.86 ± 0 ND | 31.86 ND |
| Mosquito 9 (Ct = 20.89) | 5 10 | 0 0 | 0 1 | 3 2 | ND ND | ND ND |
| Mosquito 17 (Ct = 22.87) | 5 10 | 1 0 | 0 1 | 2 2 | 33.63 ± 0 ND | 33.63 ND |
| No. of Spots | Replicate | Pool 1 Ct | No. of Spots | Replicate | Pool 2 Ct |
|---|---|---|---|---|---|
| 20 | 1 | 28.56 | 50 | 1 | 32.18 |
| 2 | 34.61 * | 2 | 32.10 | ||
| 3 | 31.96 | 3 | 32.37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malcolm, T.R.; Klein, M.J.; Petkovic, K.; Smith, I.; Blasdell, K.R. Exploring Mosquito Excreta as an Alternative Sample Type for Improving Arbovirus Surveillance in Australia. Pathogens 2025, 14, 42. https://doi.org/10.3390/pathogens14010042
Malcolm TR, Klein MJ, Petkovic K, Smith I, Blasdell KR. Exploring Mosquito Excreta as an Alternative Sample Type for Improving Arbovirus Surveillance in Australia. Pathogens. 2025; 14(1):42. https://doi.org/10.3390/pathogens14010042
Chicago/Turabian StyleMalcolm, Tess R., Melissa J. Klein, Karolina Petkovic, Ina Smith, and Kim R. Blasdell. 2025. "Exploring Mosquito Excreta as an Alternative Sample Type for Improving Arbovirus Surveillance in Australia" Pathogens 14, no. 1: 42. https://doi.org/10.3390/pathogens14010042
APA StyleMalcolm, T. R., Klein, M. J., Petkovic, K., Smith, I., & Blasdell, K. R. (2025). Exploring Mosquito Excreta as an Alternative Sample Type for Improving Arbovirus Surveillance in Australia. Pathogens, 14(1), 42. https://doi.org/10.3390/pathogens14010042