
Diagnosis and Management of Infections in Patients with Mendelian Susceptibility to Mycobacterial Disease


Abstract
1. Introduction
2. Pathophysiology and Clinical Manifestations of MSMD
3. Infectious Spectrum of MSMD

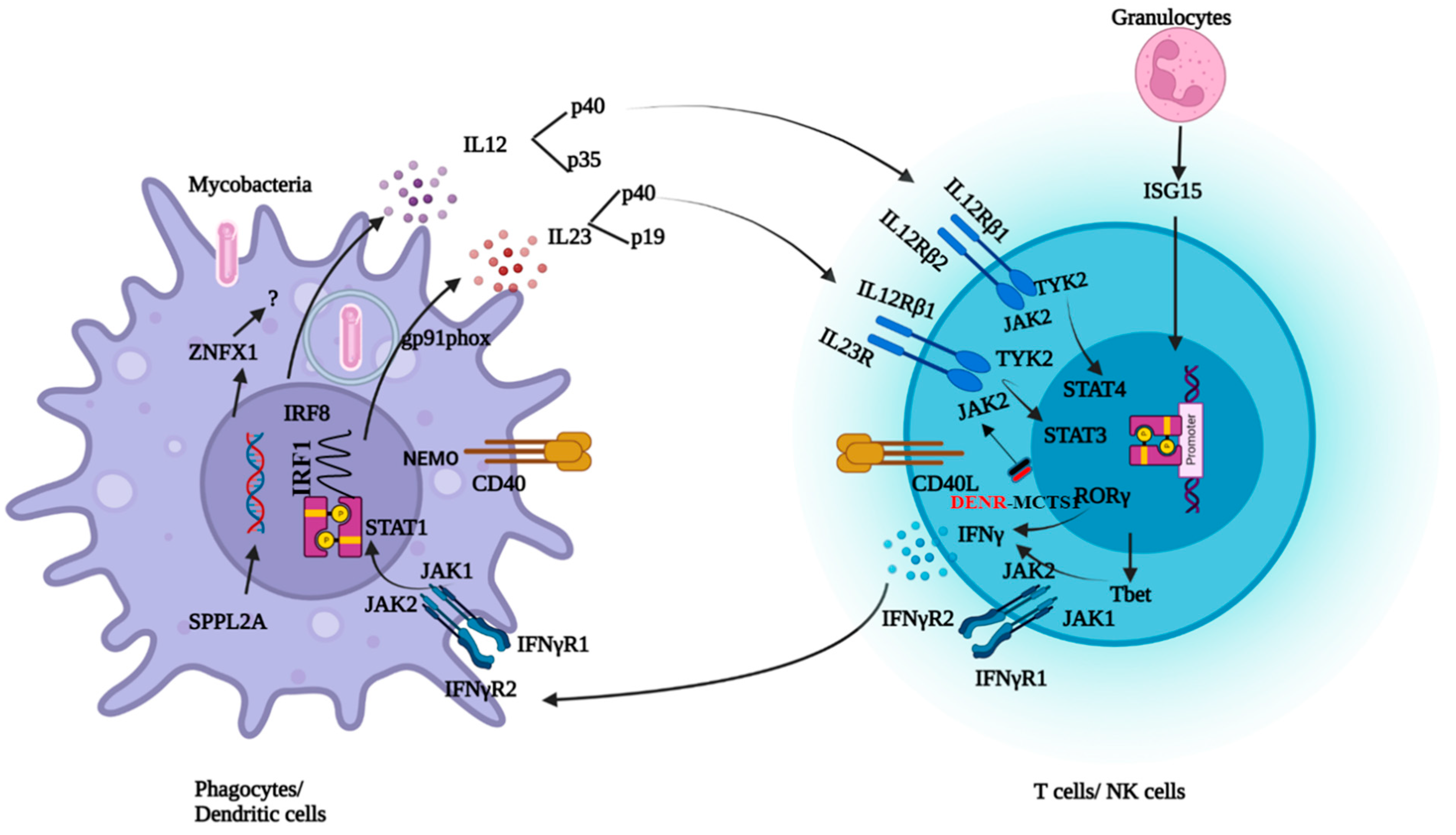{kind=link}
{kind=link}
{kind=link}
| Study | Year | Origin | No. of MSMD Patients | % of Patients Presenting with BCG Complications | BCG Adenitis | BCGosis |
|---|---|---|---|---|---|---|
| 2022 | China | 65 | 92% | 29% | 63% |
| 2022 | Mexico | 22 | 63% | 36% | 27% |
| 2022 | Iran | 24 | 100% | 33% | 67% |
| 2021 | India | 55 | 84% | - | 84% |
| 2021 | South Africa | 22 | 64% | 9% | 55% |
| 2021 | Turkey | 12 | 50% | 17% | 33% |
| 2021 | Mexico | 11 | 36% | - | 36% |
| 2012 | Egypt | 9 | 78% | - | 78% |
4. Challenges Associated with a Diagnosis of MSMD
4.1. Microbiological Diagnosis
4.2. Immunological Evaluation
4.3. Genetic Diagnosis
5. Management of Patients with MSMD
6. Hematopoietic Stem Cell Transplantation
7. Gene Therapy
8. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- van de Vosse, E.; Ottenhoff, T.H.; de Paus, R.A.; Verhard, E.M.; de Boer, T.; van Dissel, J.T.; Kuijpers, T.W. Mycobacterium bovis BCG-itis and cervical lymphadenitis due to Salmonella enteritidis in a patient with complete interleukin-12/-23 receptor beta1 deficiency. Infection 2010, 38, 128–130. [Google Scholar] [CrossRef] [PubMed]
- Bousfiha, A.; Moundir, A.; Tangye, S.G.; Picard, C.; Jeddane, L.; Al-Herz, W.; Rundles, C.C.; Franco, J.L.; Holland, S.M.; Klein, C.; et al. The 2022 Update of IUIS Phenotypical Classification for Human Inborn Errors of Immunity. J. Clin. Immunol. 2022, 42, 1508–1520. [Google Scholar] [CrossRef]
- Costagliola, G.; Peroni, D.G.; Consolini, R. Beyond Infections: New Warning Signs for Inborn Errors of Immunity in Children. Front. Pediatr. 2022, 10, 855445. [Google Scholar] [CrossRef]
- Casanova, J.L.; Abel, L. Genetic dissection of immunity to mycobacteria: The human model. Annu. Rev. Immunol. 2002, 20, 581–620. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.; Newport, M.J.; D’Souza, S.; Kalabalikis, P.; Brown, I.N.; Lenicker, H.M.; Agius, P.V.; Davies, E.G.; Thrasher, A.; Klein, N.; et al. Familial disseminated atypical mycobacterial infection in childhood: A human mycobacterial susceptibility gene? Lancet 1995, 345, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Casanova, J.L.; Jouanguy, E.; Lamhamedi, S.; Blanche, S.; Fischer, A. Immunological conditions of children with BCG disseminated infection. Lancet 1995, 346, 581. [Google Scholar] [CrossRef] [PubMed]
- Poyhonen, L.; Bustamante, J.; Casanova, J.L.; Jouanguy, E.; Zhang, Q. Life-threatening infections due to live-attenuated vaccines: Early manifestations of inborn errors of immunity. J. Clin. Immunol. 2019, 39, 376–390. [Google Scholar] [CrossRef]
- Rosain, J.; Kong, X.; Martinez-Barricarte, R.; Oleaga-Quintas, C.; Ramirez-Alejo, N.; Markle, J.; Okada, S.; Boisson-Dupuis, S.; Casanova, J.; Bustamante, J. Mendelian susceptibility to mycobacterial disease: 2014–2018 update. Immunol. Cell Biol. 2019, 97, 360–367. [Google Scholar] [CrossRef]
- Bustamante, J.; Boisson-Dupuis, S.; Abel, L.; Casanova, J.L. Mendelian susceptibility to mycobacterial disease: Genetic, immunological, and clinical features of inborn errors of IFN-gamma immunity. Semin. Immunol. 2014, 26, 454–470. [Google Scholar] [CrossRef]
- Boisson-Dupuis, S.; Bustamante, J.; El-Baghdadi, J.; Camcioglu, Y.; Parvaneh, N.; El Azbaoui, S.; Agader, A.; Hassani, A.; El Hafidi, N.; Mrani, N.A.; et al. Inherited and acquired immunodeficiencies underlying tuberculosis in childhood. Immunol. Rev. 2015, 264, 103–112. [Google Scholar] [CrossRef]
- Shabani, M.; Aleyasin, S.; Kashef, S.; Zoghi, S.; Deswarte, C.; Casanova, J.L.; Bustamante, J.; Rezaei, N. A novel recessive mutation of interferon-gamma receptor 1 in a patient with mycobacterium tuberculosis in bone marrow aspirate. J. Clin. Immunol. 2019, 39, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, J. Mendelian susceptibility to mycobacterial disease: Recent discoveries. Hum. Genet. 2020, 139, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Kerner, G.; Rosain, J.; Guérin, A.; Al-Khabaz, A.; Oleaga-Quintas, C.; Rapaport, F.; Massaad, M.J.; Ding, J.-Y.; Khan, T.; Ali, F.A.; et al. Inherited Human IFN-γ Deficiency Underlies Mycobacterial Disease. J. Clin. Investig. 2020, 130, 3158–3171. [Google Scholar] [CrossRef]
- Yang, R.; Mele, F.; Worley, L.; Langlais, D.; Rosain, J.; Benhsaien, I.; Elarabi, H.; Croft, C.A.; Doisne, J.-M.; Zhang, P.; et al. Human T-Bet Governs Innate and Innate-like Adaptive IFN-γ Immunity against Mycobacteria. Cell 2020, 183, 1826–1847.e31. [Google Scholar] [CrossRef]
- Voyer, T.L.; Neehus, A.-L.; Yang, R.; Ogishi, M.; Rosain, J.; Alroqi, F.; Alshalan, M.; Blumental, S.; Ali, F.A.; Khan, T.; et al. Inherited Deficiency of Stress Granule ZNFX1 in Patients with Monocytosis and Mycobacterial Disease. Proc. Natl. Acad. Sci. USA 2021, 118, e2102804118. [Google Scholar] [CrossRef]
- Rosain, J.; Neehus, A.-L.; Manry, J.; Yang, R.; Le Pen, J.; Daher, W.; Liu, Z.; Chan, Y.-H.; Tahuil, N.; Türel, Ö.; et al. Human IRF1 Governs Macrophagic IFN-γ Immunity to Mycobacteria. Cell 2023, 186, 621–645.e33. [Google Scholar] [CrossRef]
- Bohlen, J.; Zhou, Q.; Philippot, Q.; Ogishi, M.; Rinchai, D.; Nieminen, T.; Seyedpour, S.; Parvaneh, N.; Rezaei, N.; Yazdanpanah, N.; et al. Human MCTS1-Dependent Translation of JAK2 Is Essential for IFN-γ Immunity to Mycobacteria. Cell 2023, 186, 5114–5134.e27. [Google Scholar] [CrossRef]
- Martin-Fernandez, M.; Buta, S.; Le Voyer, T.; Li, Z.; Dynesen, L.T.; Vuillier, F.; Franklin, L.; Ailal, F.; Muglia Amancio, A.; Malle, L.; et al. A partial form of inherited human USP18 deficiency underlies infection and inflammation. J. Exp. Med. 2022, 219, e20211273. [Google Scholar] [CrossRef]
- de Jong, R.; Altare, F.; Haagen, I.A.; Elferink, D.G.; Boer, T.; van Breda Vriesman, P.J.; Kabel, P.J.; Draaisma, J.M.; van Dissel, J.T.; Kroon, F.P.; et al. Severe mycobacterial and Salmonella infections in interleukin-12 receptor-deficient patients. Science 1998, 280, 1435–1438. [Google Scholar] [CrossRef]
- de Beaucoudrey, L.; Samarina, A.; Bustamante, J.; Cobat, A.; Boisson-Dupuis, S.; Feinberg, J.; Al-Muhsen, S.; Janniere, L.; Rose, Y.; de Suremain, M.; et al. Revisiting human IL-12Rbeta1 deficiency: A survey of 141 patients from 30 countries. Medicine 2010, 89, 381–402. [Google Scholar] [CrossRef] [PubMed]
- Philippot, Q.; Ogishi, M.; Bohlen, J.; Puchan, J.; Arias, A.A.; Nguyen, T.; Martin-Fernandez, M.; Conil, C.; Rinchai, D.; Momenilandi, M.; et al. Human IL-23 Is Essential for IFN-γ–Dependent Immunity to Mycobacteria. Sci. Immunol. 2023, 8, eabq5204. [Google Scholar] [CrossRef]
- Sivanesan, D.; Beauchamp, C.; Quinou, C.; Lee, J.; Lesage, S.; Chemtob, S.; Rioux, J.D.; Michnick, S.W. IL23R (Interleukin 23 receptor) variants protective against inflammatory bowel diseases (IBD) display loss of function due to impaired protein stability and intracellular trafficking. J. Biol. Chem. 2016, 291, 8673–8685. [Google Scholar] [CrossRef]
- de Beaucoudrey, L.; Puel, A.; Filipe-Santos, O.; Cobat, A.; Ghandil, P.; Chrabieh, M.; Feinberg, J.; von Bernuth, H.; Samarina, A.; Jannière, L.; et al. Mutations in STAT3 and IL12RB1 impair the development of human IL-17-producing T cells. J. Exp. Meditor 2008, 205, 1543–1550. [Google Scholar] [CrossRef]
- Kempski, J.; Brockmann, L.; Gagliani, N.; Huber, S. T(H)17 cell and epithelial cell crosstalk during inflammatory bowel disease and carcinogenesis. Front. Immunol. 2017, 8, 1373. [Google Scholar] [CrossRef]
- Dorman, S.E.; Picard, C.; Lammas, D.; Heyne, K.; van Dissel, J.T.; Baretto, R.; Rosenzweig, S.D.; Newport, M.; Levin, M.; Roesler, J.; et al. Clinical features of dominant and recessive interferon gamma receptor 1 deficiencies. Lancet 2004, 364, 2113–2121. [Google Scholar] [CrossRef] [PubMed]
- Camcioglu, Y.; Picard, C.; Lacoste, V.; Dupuis, S.; Akcakaya, N.; Cokura, H.; Kaner, G.; Demirkesen, C.; Plancoulaine, S.; Emile, J.F.; et al. HHV-8-associated Kaposi sarcoma in a child with IFNgammaR1 deficiency. J. Pediatr. 2004, 144, 519–523. [Google Scholar] [CrossRef]
- Jouanguy, E.; Dupuis, S.; Pallier, A.; Doffinger, R.; Fondaneche, M.C.; Fieschi, C.; Lamhamedi-Cherradi, S.; Altare, F.; Emile, J.F.; Lutz, P.; et al. In a novel form of IFN-gamma receptor 1 deficiency, cell surface receptors fail to bind IFN-gamma. J. Clin. Investig. 2000, 105, 1429–1436. [Google Scholar] [CrossRef]
- Chantrain, C.F.; Bruwier, A.; Brichard, B.; Largent, V.; Chapgier, A.; Feinberg, J.; Casanova, J.L.; Stalens, J.P.; Vermylen, C. Successful hematopoietic stem cell transplantation in a child with active disseminated Mycobacterium fortuitum infection and interferon-gamma receptor 1 deficiency. Bone Marrow Transpl. 2006, 38, 75–76. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sologuren, I.; Boisson-Dupuis, S.; Pestano, J.; Vincent, Q.B.; Fernandez-Perez, L.; Chapgier, A.; Cardenes, M.; Feinberg, J.; Garcia-Laorden, M.I.; Picard, C.; et al. Partial recessive IFN-gammaR1 deficiency: Genetic, immunological, and clinical features of 14 patients from 11 kindreds. Hum. Mol. Genet. 2011, 20, 1509–1523. [Google Scholar] [CrossRef]
- Jouanguy, E.; Lamhamedi-Cherradi, S.; Lammas, D.; Dorman, S.E.; Fondaneche, M.C.; Dupuis, S.; Doffinger, R.; Altare, F.; Girdlestone, J.; Emile, J.F.; et al. A human IFNGR1 small deletion hotspot associated with dominant susceptibility to mycobacterial infection. Nat. Genet. 1999, 21, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, M.; Miki, M.; Mizoguchi, Y.; Hirata, O.; Nishimura, S.; Tamaura, M.; Kagawa, R.; Hayakawa, S.; Kobayashi, M.; Okada, S. Enhanced Osteoclastogenesis in Patients with MSMD Due to Impaired Response to IFN-γ. J. Allergy Clin. Immunol. 2022, 149, 252–261.e6. [Google Scholar] [CrossRef]
- Prando, C.; Samarina, A.; Bustamante, J.; Boisson-Dupuis, S.; Cobat, A.; Picard, C.; AlSum, Z.; Al-Jumaah, S.; Al-Hajjar, S.; Frayha, H.; et al. Inherited IL-12p40 deficiency: Genetic, immunologic, and clinical features of 49 patients from 30 kindreds. Medicine 2013, 92, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Vogt, G.; Bustamante, J.; Chapgier, A.; Feinberg, J.; Boisson Dupuis, S.; Picard, C.; Mahlaoui, N.; Gineau, L.; Alcais, A.; Lamaze, C.; et al. Complementation of a pathogenic IFNGR2 misfolding mutation with modifiers of N-glycosylation. J. Exp. Med. 2008, 205, 1729–1737. [Google Scholar] [CrossRef]
- Vogt, G.; Chapgier, A.; Yang, K.; Chuzhanova, N.; Feinberg, J.; Fieschi, C.; Boisson-Dupuis, S.; Alcais, A.; Filipe-Santos, O.; Bustamante, J.; et al. Gains of glycosylation comprise an unexpectedly large group of pathogenic mutations. Nat. Genet. 2005, 37, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Chapgier, A.; Wynn, R.F.; Jouanguy, E.; Filipe-Santos, O.; Zhang, S.; Feinberg, J.; Hawkins, K.; Casanova, J.L.; Arkwright, P.D. Human complete Stat-1 deficiency is associated with defective type I and II IFN responses in vitro but immunity to some low virulence viruses in vivo. J. Immunol. 2006, 176, 5078–5083. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, S.; Jouanguy, E.; Al-Hajjar, S.; Fieschi, C.; Al-Mohsen, I.Z.; Al-Jumaah, S.; Yang, K.; Chapgier, A.; Eidenschenk, C.; Eid, P.; et al. Impaired response to interferon-alpha/beta and lethal viral disease in human STAT1 deficiency. Nat. Genet. 2003, 33, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Vairo, D.; Tassone, L.; Tabellini, G.; Tamassia, N.; Gasperini, S.; Bazzoni, F.; Plebani, A.; Porta, F.; Notarangelo, L.D.; Parolini, S.; et al. Severe impairment of IFN-gamma and IFN-alpha responses in cells of a patient with a novel STAT1 splicing mutation. Blood 2011, 118, 1806–1817. [Google Scholar] [CrossRef] [PubMed]
- Chapgier, A.; Kong, X.F.; Boisson-Dupuis, S.; Jouanguy, E.; Averbuch, D.; Feinberg, J.; Zhang, S.Y.; Bustamante, J.; Vogt, G.; Lejeune, J.; et al. A partial form of recessive STAT1 deficiency in humans. J. Clin. Investig. 2009, 119, 1502–1514. [Google Scholar] [CrossRef]
- Averbuch, D.; Chapgier, A.; Boisson-Dupuis, S.; Casanova, J.L.; Engelhard, D. The clinical spectrum of patients with deficiency of Signal Transducer and Activator of Transcription-1. Pediatr. Infect. Dis. J. 2011, 30, 352–355. [Google Scholar] [CrossRef]
- Kong, X.F.; Ciancanelli, M.; Al-Hajjar, S.; Alsina, L.; Zumwalt, T.; Bustamante, J.; Feinberg, J.; Audry, M.; Prando, C.; Bryant, V.; et al. A novel form of human STAT1 deficiency impairing early but not late responses to interferons. Blood 2010, 116, 5895–5906. [Google Scholar] [CrossRef]
- Kristensen, I.A.; Veirum, J.E.; Moller, B.K.; Christiansen, M. Novel STAT1 alleles in a patient with impaired resistance to mycobacteria. J. Clin. Immunol. 2011, 31, 265–271. [Google Scholar] [CrossRef]
- Shamriz, O.; Engelhard, D.; Rajs, A.P.; Kaidar-Shwartz, H.; Casanova, J.L.; Averbuch, D. Mycobacterium szulgai chronic multifocal osteomyelitis in an adolescent with inherited STAT1 deficiency. Pediatr. Infect. Dis. J. 2013, 32, 1345–1347. [Google Scholar] [CrossRef]
- Filipe-Santos, O.; Bustamante, J.; Haverkamp, M.H.; Vinolo, E.; Ku, C.L.; Puel, A.; Frucht, D.M.; Christel, K.; von Bernuth, H.; Jouanguy, E.; et al. X-linked susceptibility to mycobacteria is caused by mutations in NEMO impairing CD40-dependent IL-12 production. J. Exp. Med. 2006, 203, 1745–1759. [Google Scholar] [CrossRef]
- Stuber, E.; Strober, W.; Neurath, M. Blocking the CD40L-CD40 interaction in vivo specifically prevents the priming of T helper 1 cells through the inhibition of interleukin 12 secretion. J. Exp. Med. 1996, 183, 693–698. [Google Scholar] [CrossRef]
- Bustamante, J.; Arias, A.A.; Vogt, G.; Picard, C.; Galicia, L.B.; Prando, C.; Grant, A.V.; Marchal, C.C.; Hubeau, M.; Chapgier, A.; et al. Germline CYBB Mutations That Selectively Affect Macrophages in Kindreds with X-Linked Predisposition to Tuberculous Mycobacterial Disease. Nat. Immunol. 2011, 12, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.-F.; Martinez-Barricarte, R.; Kennedy, J.; Mele, F.; Lazarov, T.; Deenick, E.K.; Ma, C.S.; Breton, G.; Lucero, K.B.; Langlais, D.; et al. Disruption of an Antimycobacterial Circuit between Dendritic and Helper T Cells in Human SPPL2a Deficiency. Nat. Immunol. 2018, 19, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Ong, R.Y.L.; Chan, S.-W.B.; Chew, S.J.; Liew, W.K.; Thoon, K.C.; Chong, C.-Y.; Yung, C.F.; Sng, L.-H.; Tan, A.M.; Bhattacharyya, R.; et al. DISSEMINATED BACILLUS-CALMETTE-GUÉRIN INFECTIONS AND PRIMARY IMMUNODEFICIENCY DISORDERS IN SINGAPORE: A SINGLE CENTER 15-YEAR RETROSPECTIVE REVIEW. Int. J. Infect. Dis. 2020, 97, 117–125. [Google Scholar] [CrossRef]
- Ying, W.; Sun, J.; Liu, D.; Hui, X.; Yu, Y.; Wang, J.; Wang, X. Clinical characteristics and immunogenetics of BCGosis/BCGitis in Chinese children: A 6-year follow-up study. PLoS ONE 2014, 9, e94485. [Google Scholar] [CrossRef]
- Sadeghi-Shabestari, M.; Rezaei, N. Disseminated bacille CalmetteGuerin in Iranian children with severe combined immunodeficiency. Int. J. Infect. Dis. 2009, 13, e420–e423. [Google Scholar] [CrossRef] [PubMed]
- Bernatowska, E.A.; Wolska-Kusnierz, B.; Pac, M.; Kurenko-Deptuch, M.; Zwolska, Z.; Casanova, J.L.; Piatosa, B.; van Dongen, J.; Roszkowski, K.; Mikoluc, B.; et al. Disseminated bacillus Calmette-Guerin infection and immunodeficiency. Emerg. Infect. Dis. 2007, 13, 799–801. [Google Scholar] [CrossRef]
- Glanzmann, B.; Uren, C.; de Villiers, N.; van Coller, A.; Glashoff, R.H.; Urban, M.; Hoal, E.G.; Esser, M.M.; Möller, M.; Kinnear, C.J. Primary immunodeficiency diseases in a tuberculosis endemic region: Challenges and opportunities. Genes. Immun. 2018, 20, 447–454. [Google Scholar] [CrossRef]
- Lee, W.I.; Huang, J.L.; Yeh, K.W.; Jaing, T.H.; Lin, T.Y.; Huang, Y.C.; Chiu, C.H. Immune defects in active mycobacterial diseases in patients with primary immunodeficiency diseases (PIDs). J. Formos. Med. Assoc. 2011, 110, 750–758. [Google Scholar] [CrossRef]
- Gimenez-Sanchez, F.; Cobos-Carrascosa, E.; Sanchez-Forte, M.; Martinez-Lirola, M.; Lopez-Ruzafa, E.; Galera-Martinez, R.; Del Rosal, T.; Martinez-Gallo, M. Different Penetrance of Disseminated Infections Caused by Nontuberculous Mycobacteria in Mendelian Susceptibility to Mycobacterial Disease Associated with a Novel Mutation. Pediatr. Infect. Dis. J. 2014, 33, 328–330. [Google Scholar] [CrossRef]
- Peñafiel Vicuña, A.K.; Yamazaki Nakashimada, M.; León Lara, X.; Mendieta Flores, E.; Nuñez Núñez, M.E.; Lona-Reyes, J.C.; Hernández Nieto, L.; Ramírez Vázquez, M.G.; Barroso Santos, J.; López Iñiguez, Á.; et al. Mendelian Susceptibility to Mycobacterial Disease: Retrospective Clinical and Genetic Study in Mexico. J. Clin. Immunol. 2022, 6, 210–220. [Google Scholar] [CrossRef]
- Mahdaviani, S.A.; Fallahi, M.; Jamee, M.; Marjani, M.; Tabarsi, P.; Moniri, A.; Farnia, P.; Daneshmandi, Z.; Parvaneh, N.; Casanova, J.L.; et al. Effective anti-mycobacterial treatment for BCG disease in patients with Mendelian Susceptibility to Mycobacterial Disease (MSMD): A case series. Ann. Clin. Microbiol. Antimicrob. 2022, 21, 8. [Google Scholar] [CrossRef]
- Taur, P.D.; Gowri, V.; Pandrowala, A.A.; Iyengar, V.V.; Chougule, A.; Golwala, Z.; Chandak, S.; Agarwal, R.; Keni, P.; Dighe, N.; et al. Clinical and Molecular Findings in Mendelian Susceptibility to Mycobacterial Diseases: Experience From India. Front. Immunol. 2021, 12, 631298. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, H.M.; Glanzmann, B.; Van Coller, A.; Engelbrecht, C.; Abraham, D.R.; Reddy, K.; Möller, M.; Kinnear, C.; Glashoff, R.H.; Esser, M. Mendelian Susceptibility to Mycobacterial Disease in Tuberculosis-Hyperendemic South Africa. S. Afr. Med. J. 2021, 111, 998. [Google Scholar] [CrossRef]
- Azarsiz, E.; Karaca, N.; Karaca, E.; Aksu, G.; Genel, F.; Gulez, N.; Ozen, S.; Kutukculer, N. Eight years of follow-up experience in children with mendelian susceptibility to mycobacterial disease and review of the literature. Asian Pac. J. Allergy Immunol. 2023, 41, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Radwan, N.; Nademi, Z.; Lum, S.H.; Flood, T.; Abinun, M.; Owens, S.; Williams, E.; Gennery, A.R.; Hambleton, S.; Slatter, M.A. Outcome of Hematopoietic Stem Cell Transplantation in patients with Mendelian Susceptibility to Mycobacterial Diseases. J. Clin. Immunol. 2021, 41, 1774–1780. [Google Scholar] [CrossRef] [PubMed]
- Galal, N.; Boutros, J.; Casanova, A.M.; Boisson-Dupuis, S.; Marsafy, A.; Kong, X.-F.; Dupuis, J.F.; Mendelian, J.B. Susceptibility to Mycobacterial o Mycobacterial Disease in Egyptian Children n Egyptian Children. Mediterr. J. Hematol. Infect. Dis. 2012, 4, e2012033 DOI 104084/MJHID2012033. [Google Scholar]
- Bernatowska, E.; Pac, M.; Heropolitańska-Pliszka, E.; Pietrucha, B.; Dąbrowska-Leonik, N.; Skomska-Pawliszak, M.; Bernat-Sitarz, K.; Krzysztopa-Grzybowska, K.; Wolska-Kuśnierz, B.; Bohynikova, N.; et al. BCG Moreau Polish Substrain Infections in Patients With Inborn Errors of Immunity: 40 Years of Experience in the Department of Immunology, Children’s Memorial Health Institute, Warsaw. Front. Pediatr. 2022, 10, 839111. [Google Scholar] [CrossRef]
- Lyra, P.T.; Souza, E.; Moura, A.C.A.; Matta, M.C.; Torres, L.C.; Coelho, A.V.C.; Rocha, M.Â.W.; Arraes, L.; Oliveira, J.B. Inborn Errors of Immunity in Patients with Adverse Events Following BCG Vaccination in Brazil. J. Clin. Immunol. 2022, 42, 1708–1720. [Google Scholar] [CrossRef]
- Yadav, R.M.; Dalvi, A.; Gupta, M.; Bargir, U.A.; Shabrish, S.; Aluri, J.; Kulkarni, M.; Hule, G.; Kambli, P.; Setia, P.; et al. Spectrum of Inborn errors of immunity in a cohort of 90 patients presenting with complications to BCG vaccination in India. Scand. J. Immunol. 2021, 93, e13010. [Google Scholar] [CrossRef] [PubMed]
- Ong, R.Y.L.; Chan, S.B.; Chew, S.J.; Liew, W.K.; Thoon, K.C.; Chong, C.Y.; Yung, C.F.; Sng, L.H.; Tan, A.M.; Bhattacharyya, R.; et al. Disseminated bacillus-calmette-guérin infections and primary immunodeficiency disorders in singapore: A single center 15-year retrospective review. Int. J. Infect. Dis. 2020, 97, 117–125. [Google Scholar] [CrossRef]
- Sohani, M.; Habibi, S.; Delavari, S.; Shahkarami, S.; Yazdani, R.; Shirmast, P.; Nazari, F.; Moeini Shad, T.; Mamishi, S.; Azizi, G.; et al. Evaluation of patients with primary immunodeficiency associated with Bacille Calmette-Guerin (BCG)-vaccine-derived complications. Allergol. Immunopathol. 2020, 48, 729–737. [Google Scholar] [CrossRef]
- Norouzi, S.; Aghamohammadi, A.; Mamishi, S.; Rosenzweig, S.D.; Rezaei, N. Bacillus Calmette-Guérin (BCG) complications associated with primary immunodeficiency diseases. J. Infect. 2012, 64, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Tabarsi, P.; Marjani, M.; Mansouri, N.; Farnia, P.; Boisson-Dupuis, S.; Bustamante, J.; Abel, L.; Adimi, P.; Casanova, J.L.; Mansouri, D. Lethal tuberculosis in a previously healthy adult with IL-12 receptor deficiency. J. Clin. Immunol. 2011, 31, 537–539. [Google Scholar] [CrossRef] [PubMed]
- Caragol, I.; Raspall, M.; Fieschi, C.; Feinberg, J.; Larrosa, M.N.; Hernandez, M.; Figueras, C.; Bertran, J.M.; Casanova, J.L.; Espanol, T. Clinical tuberculosis in 2 of 3 siblings with interleukin-12 receptor beta1 deficiency. Clin. Infect. Dis. 2003, 37, 302–306. [Google Scholar] [CrossRef]
- Alcais, A.; Fieschi, C.; Abel, L.; Casanova, J.L. Tuberculosis in children and adults: Two distinct genetic diseases. J. Exp. Med. 2005, 202, 1617–1621. [Google Scholar] [CrossRef] [PubMed]
- Ozbek, N.; Fieschi, C.; Yilmaz, B.T.; de Beaucoudrey, L.; Demirhan, B.; Feinberg, J.; Bikmaz, Y.E.; Casanova, J.L. Interleukin-12 receptor beta 1 chain deficiency in a child with disseminated tuberculosis. Clin. Infect. Dis. 2005, 40, e55–e58. [Google Scholar] [CrossRef]
- Brown-Elliott, B.A.; Philley, J.V. Rapidly Growing Mycobacteria. Microbiol. Spectr. 2017, 5, 703–723. [Google Scholar] [CrossRef]
- Sharma, S.; Upadhyay, V. Epidemiology, Diagnosis & Treatment of Non-Tuberculous Mycobacterial Diseases. Indian. J. Med. Res. 2020, 152, 185. [Google Scholar] [CrossRef]
- Johnson, M.M.; Odell, J.A. Nontuberculous Mycobacterial Pulmonary Infections. J. Thorac. Dis. 2014, 6, 11. [Google Scholar]
- Schulz, S.M.; Köhler, G.; Schütze, N.; Knauer, J.; Straubinger, R.K.; Chackerian, A.A.; Witte, E.; Wolk, K.; Sabat, R.; Iwakura, Y.; et al. Protective Immunity to Systemic Infection with Attenuated Salmonella enterica Serovar Enteritidis in the Absence of IL-12 Is Associated with IL-23-Dependent IL-22, but Not IL-17. J. Immunol. 2008, 181, 7891–7901. [Google Scholar] [CrossRef]
- MacLennan, C.; Fieschi, C.; Lammas, D.A.; Picard, C.; Dorman, S.E.; Sanal, O.; MacLennan, J.M.; Holland, S.M.; Ottenhoff, T.H.; Casanova, J.L.; et al. Interleukin (IL)-12 and IL-23 are key cytokines for immunity against Salmonella in humans. J. Infect. Dis. 2004, 190, 1755–1757. [Google Scholar] [CrossRef]
- Happel, K.I.; Dubin, P.J.; Zheng, M.; Ghilardi, N.; Lockhart, C.; Quinton, L.J.; Odden, A.R.; Shellito, J.E.; Bagby, G.J.; Nelson, S.; et al. Divergent roles of IL-23 and IL-12 in host defense against Klebsiella pneumoniae. J. Exp. Med. 2005, 202, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Ozen, M.; Ceyhan, M.; Sanal, O.; Bayraktar, M.; Mesci, L. Recurrent Salmonella bacteremia in interleukin-12 receptor beta1 deficiency. J. Trop. Pediatr. 2006, 52, 296–298. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Filiz, S.; Kocacik Uygun, D.F.; Verhard, E.M.; van Dissel, J.T.; Uygun, V.; Bassorgun, C.; Bingol, A.; Yegin, O.; van de Vosse, E. Cutaneous leukocytoclastic vasculitis due to Salmonella enteritidis in a child with interleukin-12 receptor beta-1 deficiency. Pediatr. Dermatol. 2014, 31, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Alejo, N.; Santos-Argumedo, L. Innate Defects of the IL-12/IFN-γ Axis in Susceptibility to Infections by Mycobacteria and Salmonella. J. Interferon Cytokine Res. 2014, 34, 307–317. [Google Scholar] [CrossRef] [PubMed]
- van Coller, A.; Glanzmann, B.; Cornelissen, H.; Möller, M.; Kinnear, C.; Esser, M.; Glashoff, R. Phenotypic and immune functional profiling of patients with suspected Mendelian Susceptibility to Mycobacterial Disease in South Africa. BMC Immunol. 2021, 22, 62. [Google Scholar] [CrossRef] [PubMed]
- Wu, U.-I.; Chuang, Y.-C.; Sheng, W.-H.; Sun, H.-Y.; Jhong, Y.-T.; Wang, J.-Y.; Chang, S.-C.; Wang, J.-T.; Chen, Y.-C. Use of QuantiFERON-TB Gold Intube assay in screening for neutralizing anti-interferon-γ autoantibodies in patients with disseminated nontuberculous mycobacterial infection. Clin. Microbiol. Infect. 2018, 24, 159–165. [Google Scholar] [CrossRef]
- Meyts, I.; Bosch, B.; Bolze, A.; Boisson, B.; Itan, Y.; Belkadi, A.; Pedergnana, V.; Moens, L.; Picard, C.; Cobat, A.; et al. Exome and genome sequencing for inborn errors of immunity. J. Allergy Clin. Immunol. 2016, 138, 957–969. [Google Scholar] [CrossRef]
- Rosain, J.; Oleaga-Quintas, C.; Deswarte, C.; Verdin, H.; Marot, S.; Syridou, G.; Mansouri, M.; Mahdaviani, S.A.; Venegas-Montoya, E.; Tsolia, M.; et al. A Variety of Alu-Mediated Copy Number Variations Can Underlie IL-12Rbeta1 Deficiency. J. Clin. Immunol. 2018, 38, 617–627. [Google Scholar] [CrossRef]
- Xia, L.; Liu, X.-H.; Yuan, Y.; Lowrie, D.B.; Fan, X.-Y.; Li, T.; Hu, Z.-D.; Lu, S.-H. An Updated Review on MSMD Research Globally and A Literature Review on the Molecular Findings, Clinical Manifestations, and Treatment Approaches in China. Front. Immunol. 2022, 13, 18. [Google Scholar] [CrossRef]
- Nekooie-Marnany, N.; Deswarte, C.; Ostadi, V.; Bagherpour, B.; Taleby, E.; Ganjalikhani-Hakemi, M.; Le Voyer, T.; Rahimi, H.; Rosain, J.; Pourmoghadas, Z.; et al. Impaired IL-12- and IL-23-Mediated Immunity Due to IL-12Rβ1 Deficiency in Iranian Patients with Mendelian Susceptibility to Mycobacterial Disease. J. Clin. Immunol. 2018, 38, 787–793. [Google Scholar] [CrossRef]
- Obregón-Henao, A.; Arnett, K.A.; Henao-Tamayo, M.; Massoudi, L.; Creissen, E.; Andries, K.; Lenaerts, A.J.; Ordway, D.J. Susceptibility of Mycobacterium Abscessus to Antimycobacterial Drugs in Preclinical Models. Antimicrob. Agents Chemother. 2015, 59, 6904–6912. [Google Scholar] [CrossRef]
- Kuruvilla, M.; de la Morena, M.T. Antibiotic Prophylaxis in Primary Immune Deficiency Disorders. J. Allergy Clin. Immunol. Pract. 2013, 1, 573–582. [Google Scholar] [CrossRef]
- Rosenzweig, S.D.; Holland, S.M. Defects in the interferon-gamma and interleukin-12 pathways. Immunol. Rev. 2005, 203, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Alaki, E.M.; Aljobair, F.; Alaklobi, F.; Al Shamrani, M.; Al-Zahim, F.; Dongues, A.; Casanova, J.-L. Chronic Disseminated Salmonellosis in a Patient With Interleukin-12p40 Deficiency. Pediatr. Infect. Dis. J. 2018, 37, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Uppuluri, R.; Vellaichamy Swaminathan, V.; Ravichandran, N.; Melarcode Ramanan, K.; Raj, R. Mendelian susceptibility to mycobacterial disease-Challenges in hematopoietic stem cell transplantation. Pediatr. Blood Cancer 2020, 67, e28187. [Google Scholar] [CrossRef] [PubMed]
- Rottman, M.; Soudais, C.; Vogt, G.; Renia, L.; Emile, J.-F.; Decaluwe, H.; Gaillard, J.-L.; Casanova, J.-L. IFN-gamma mediates the rejection of haematopoietic stem cells in IFN-gammaR1-deficient hosts. PLoS Med. 2008, 5, e26. [Google Scholar] [CrossRef] [PubMed]
- Kohn, L.A.; Kohn, D.B. Gene Therapies for Primary Immune Deficiencies. Front. Immunol. 2021, 12, 648951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Thrasher, A.J.; Zhang, F. Gene therapy and genome editing for primary immunodeficiency diseases. Genes. Dis. 2019, 7, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Hahn, K.; Pollmann, L.; Nowak, J.; Nguyen, A.H.H.; Haake, K.; Neehus, A.-L.; Waqas, S.F.H.; Pessler, F.; Baumann, U.; Hetzel, M.; et al. Human Lentiviral Gene Therapy Restores the Cellular Phenotype of Autosomal Recessive Complete IFN-γR1 Deficiency. Mol. Ther. Methods Clin. Dev. 2020, 17, 785–795. [Google Scholar] [CrossRef]
- Hetzel, M.; Lopez-Rodriguez, E.; Mucci, A.; Nguyen, A.H.H.; Suzuki, T.; Shima, K.; Buchegger, T.; Dettmer, S.; Rodt, T.; Bankstahl, J.P.; et al. Effective hematopoietic stem cell-based gene therapy in a murine model of hereditary pulmonary alveolar proteinosis. Haematologica 2020, 105, 1147–1157. [Google Scholar] [CrossRef]



| Clinical Manifestations | Mycobacterial Infections | Bacterial Infections | Viral Infections | Fungal Infections | Parasitic Infections | References | |
|---|---|---|---|---|---|---|---|
| AR complete IFN-γR1 deficiency | Skin lesions, Pneumonia, B-cell lymphoma, pineal germinoma, and Kaposi sarcoma | MAC, disseminated M. kansasii, M. chelonae, M. fortuitum, M. mageritense, M. peregrinum, M. smegmatis, M. scrofulaceum, M. tuberculosis, M. tuberculosis | Listeria monocytogenes, Streptococcus | Cytomegalovirus (CMV), varicella-zoster virus (VZV), human herpes virus-8 (HHV8), piscine ortho-reovirus subtype 3 (PRV-3), respiratory syncytial virus (RSV)), parainfluenza virus (PIV) | [9,25,26,27,28] | ||
| AR partial IFN-γR1 deficiency | Osteomyelitis | BCG, M. avium, M. avium complex, M. abcessus, M. szulgai, M. tuberculosis | Salmonella sp., Klebsiella pneumoniae, Haemophilus influenzae, Legionella sp., Mycoplasma pneumoniae, Shigella sonnei | VZV, molluscum contagiosum, RSV | Toxoplasma gondii, Cryptosporidium sp. | [9] | |
| AD partial IFN-γR1 deficiency | Osteomyelitis | BCG, M. abcessus, M. avium complex, M. asiaticum, M. bohemicum, M. chelonei, M. gordonae, M. kansasii, M. scrofulaceum, M. bovis, M. tuberculosis, Coccidiodes sp. | Salmonella sp. | VZV | Histoplasma capsulatum, Cocciodiodes spp. | [9,25] | |
| AR complete IFN-γR2 defect | cutaneous squamous cell carcinoma | BCG, M. abscessus, M. avium, M. fortuitum, M. porcium, and M. simiae | Salmonella sp. | CMV | [9,33] | ||
| AR partial IFN-γR2 defect | Osteomylitis | BCG, M. abscessus, M. bovis, M. elephantis, M. fortuitum, and M. simiae | [9] | ||||
| AD partial IFN-γR2 defect | Osteomylitis, lymphoreticular, pulmonary, gastrointestinal, central nervous system | BCG disease, M. bovis | [9] | ||||
| IFN-γ deficiency | disseminated BCGosis | [13] | |||||
| IL12Rβ1 deficiency | Chronic mucocutaneous candidiasis, leukocytoclastic vasculitis, esophageal carcinoma, Inflammatory bowel disease (IBD) | BCG, M. avium, M. avium intracellulare complex, M. chelonae, M. fortuitum, M. fortuitum-chelonae complex, M. genevense, M. gordonae, M. tilburgii, M. triplex, M. simiae | S. enteritidis, S. typhimurium, S. dublin, S. hadar, S. typhi O and typhi H, S. group B and D, S. portland, S. paratyphi, Klebsiella pneumoniae, Nocardia | Candida sp., Cryptococcus neoformans, Coccidiodes sp., Paraccocidiodes brasiliensis, and Histoplasma sp. | Toxoplasma and Leishmania | [8,21,22,23,19] | |
| gp91phox deficiency Q231P and T178P | disseminated BCGosis and regional BCGitis | M. tuberculosis | Herpes zoster | [9,45] | |||
| NEMO deficiency | Cervical abscess | disseminated mycobacterial diseases (Mycobacterium avium complex is the most common); recurrent BCG infection | Haemophilus influenzae type b, Enterobacter sp., and Salmonella sp. | [9] | |||
| IRF8 deficiency | disseminated BCGosis, calcifications in the brain parenchyma, oral candidiasis | BCG | Viral infections | Candida spp. | [9] | ||
| STAT1 deficiency | Osteomyelitis | BCG, M. avium, M. szulgai, M. tuberculosis | Salmonella | EBV, CMV and VZV | [9,36,38,41] | ||
| ISG15 deficiency | Intracranial calcifications, recurrent inflammatory necrotizing ulcerative skin lesions, Pulmonary involvement, epileptic seizures | BCG | E. coli, and methicillin-resistant Staphylococcus aureus | Aspergillus sp. | [9] | ||
| IL-12B deficiency | BCG disease | BCG, M. tuberculosis | Salmonella, Nocardia sp., and Klebsiella sp. | Candida sp. | [9,20,32] | ||
| IL-23 receptor deficiency | BCGadenitis, BCGosis | BCG disease | [12] | ||||
| IL-12Rβ2 deficiency | pulmonary TB | BCG disease | [12] | ||||
| TYK2 deficiency | BCG disease, atopy | BCG and M. tuberculosis | Staphyalococcus sp., Other intracellular bacteria | Herpes virus family | Candida spp. | [8] | |
| SPPL2A deficiency | BCG lymphadenitis, spastic paraplegia | BCG | [8,12,46] | ||||
| RORγ deficiency | CMC, with onychomycosis, disseminated BCG disease, Hypoplasia of the thymus and impaired lymphoid development, pneumonia | BCG, Mycobacterium tuberculosis | [8] | ||||
| JAK1 deficiency | recurrent sinopulmonary infections, global developmental delay, bladder carcinoma | mycobacterial infection | Viral infections | [8] | |||
| T-bet deficiency | BCGosis in early infancy, reactive airway disease | BCG | [14] | ||||
| USP18 deficiency | Intracranial calcifications | BCG | [18] | ||||
| ZNFX1 deficiency | BCGosis, recurrent respiratory infections, pneumonia, and interstitial lung disease | BCG | CMV | [15] | |||
| IRF1 deficiency | BCG adenitis, BCGosis | BCG, M. avium intracellulare | Varicella zoster virus | Histoplasma | [16] | ||
| MCST1 deficiency | BCGosis, osteomyelitis | BCG | [17] |
| Study | Year | Origin | Total No. of IEI Patients with BCG Complications | No. of SCID Patients | No. of CGD Patients | No. of MSMD Patients | No. of Other IEI Patients |
|---|---|---|---|---|---|---|---|
| 2022 | Poland | 180 | 72 | 63 | 6 | 39 |
| 2022 | Brazil | 9 | 2 | 4 | 3 | - |
| 2020 | India | 52 | 13 | 15 | 19 | 5 |
| 2020 | Singapore | 10 | 4 | - | 3 | 3 |
| 2020 | Iran | 137 | 75 | 15 | 47 | - |
| 2010 | China | 12 | 3 | 7 | 2 | - |
| 2010 | Portugal | 3 | 1 | - | 2 | - |
| 2009 | Taiwan | 18 | 12 | 2 | 3 | 1 |
| 2007 | Japan | 19 | 4 | 5 | 4 | 6 |
| 2007 | France | 3 | - | - | 3 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalvi, A.; Bargir, U.A.; Natraj, G.; Shah, I.; Madkaikar, M. Diagnosis and Management of Infections in Patients with Mendelian Susceptibility to Mycobacterial Disease. Pathogens 2024, 13, 203. https://doi.org/10.3390/pathogens13030203
Dalvi A, Bargir UA, Natraj G, Shah I, Madkaikar M. Diagnosis and Management of Infections in Patients with Mendelian Susceptibility to Mycobacterial Disease. Pathogens. 2024; 13(3):203. https://doi.org/10.3390/pathogens13030203
Chicago/Turabian StyleDalvi, Aparna, Umair Ahmed Bargir, Gita Natraj, Ira Shah, and Manisha Madkaikar. 2024. "Diagnosis and Management of Infections in Patients with Mendelian Susceptibility to Mycobacterial Disease" Pathogens 13, no. 3: 203. https://doi.org/10.3390/pathogens13030203
APA StyleDalvi, A., Bargir, U. A., Natraj, G., Shah, I., & Madkaikar, M. (2024). Diagnosis and Management of Infections in Patients with Mendelian Susceptibility to Mycobacterial Disease. Pathogens, 13(3), 203. https://doi.org/10.3390/pathogens13030203