
Detection of Coronaviruses in Bats in Lebanon during 2020

 ,
,  ,
,  ,
,  ,
,  , , , , and
, , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Processing
2.2. Molecular Detection
2.3. Phylogenetic Analysis
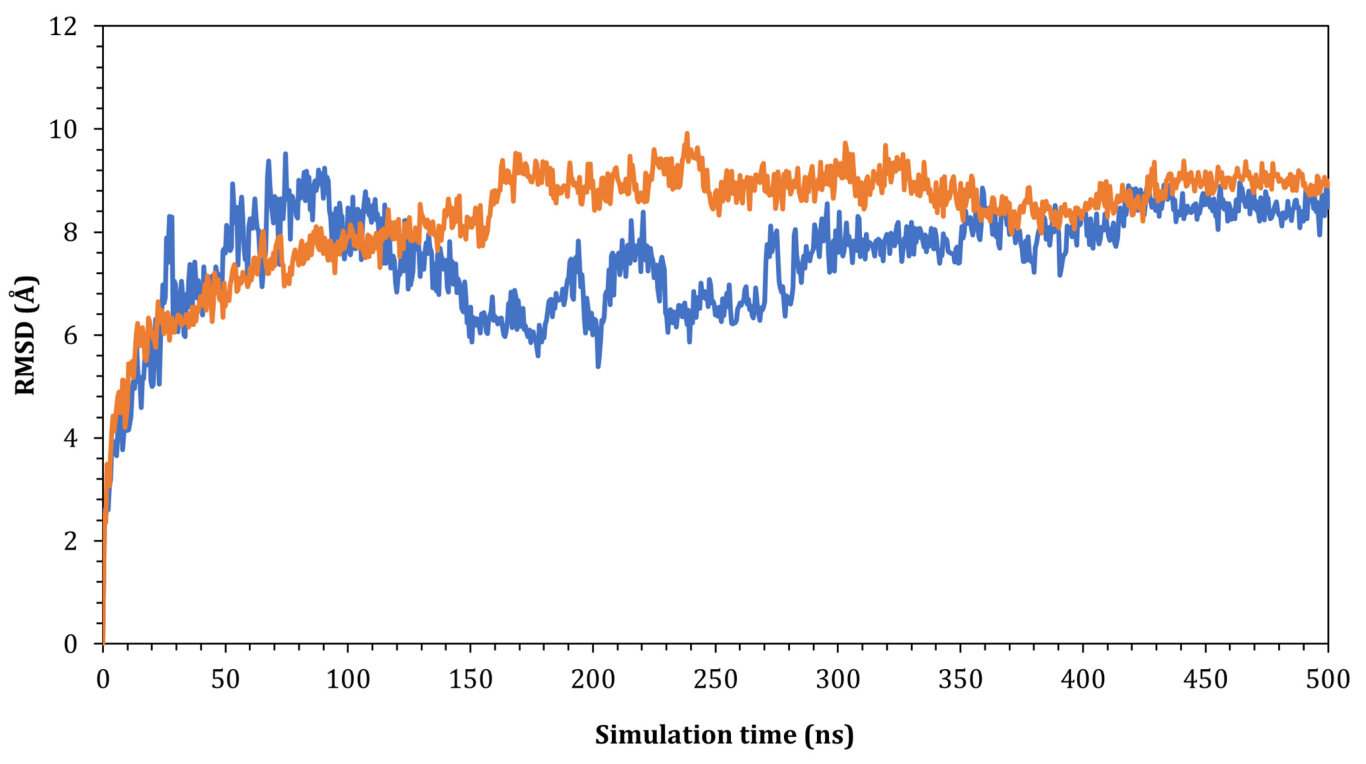
2.4. Homology Modeling, Protein-Protein Docking, and Molecular Dynamic Simulation
2.5. Statistical Analysis
3. Results
3.1. Prevalence of CoVs in Bats
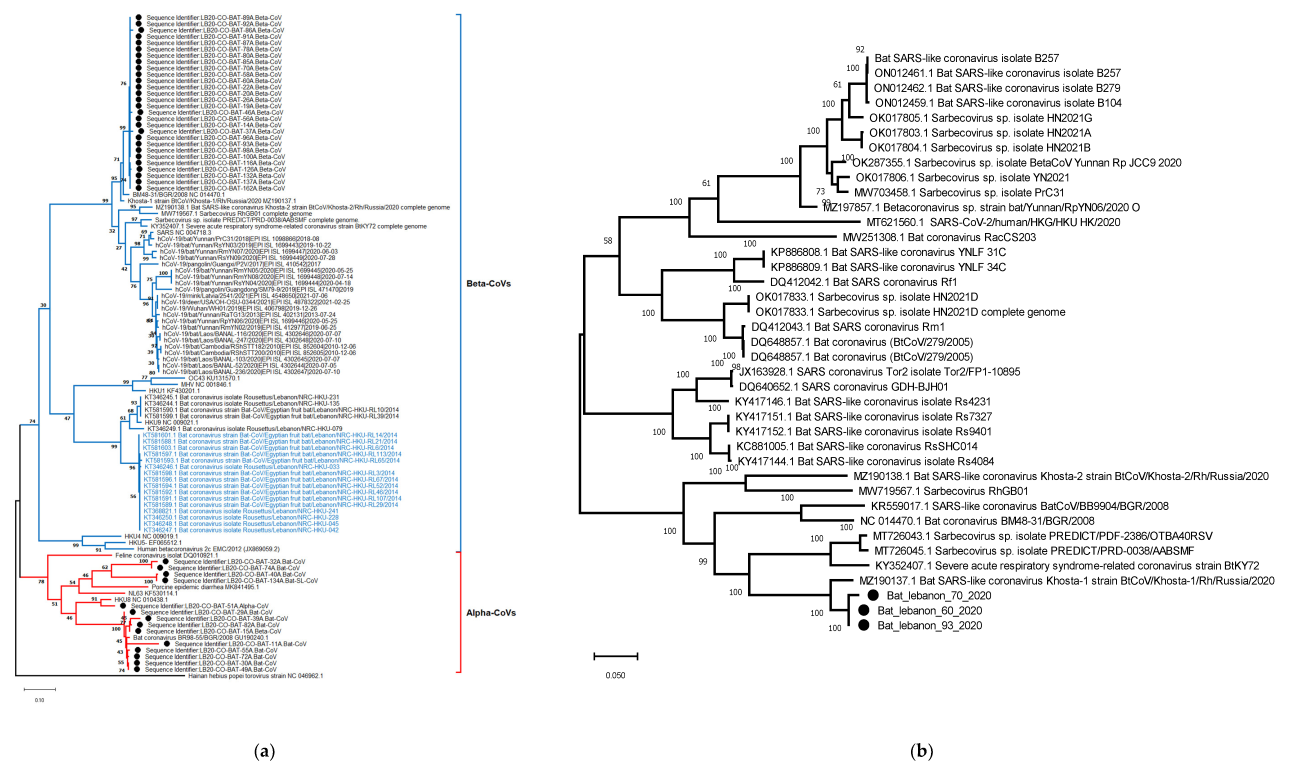
3.2. Diversity of CoVs in Lebanon Based on Partial Sequence of RdRp
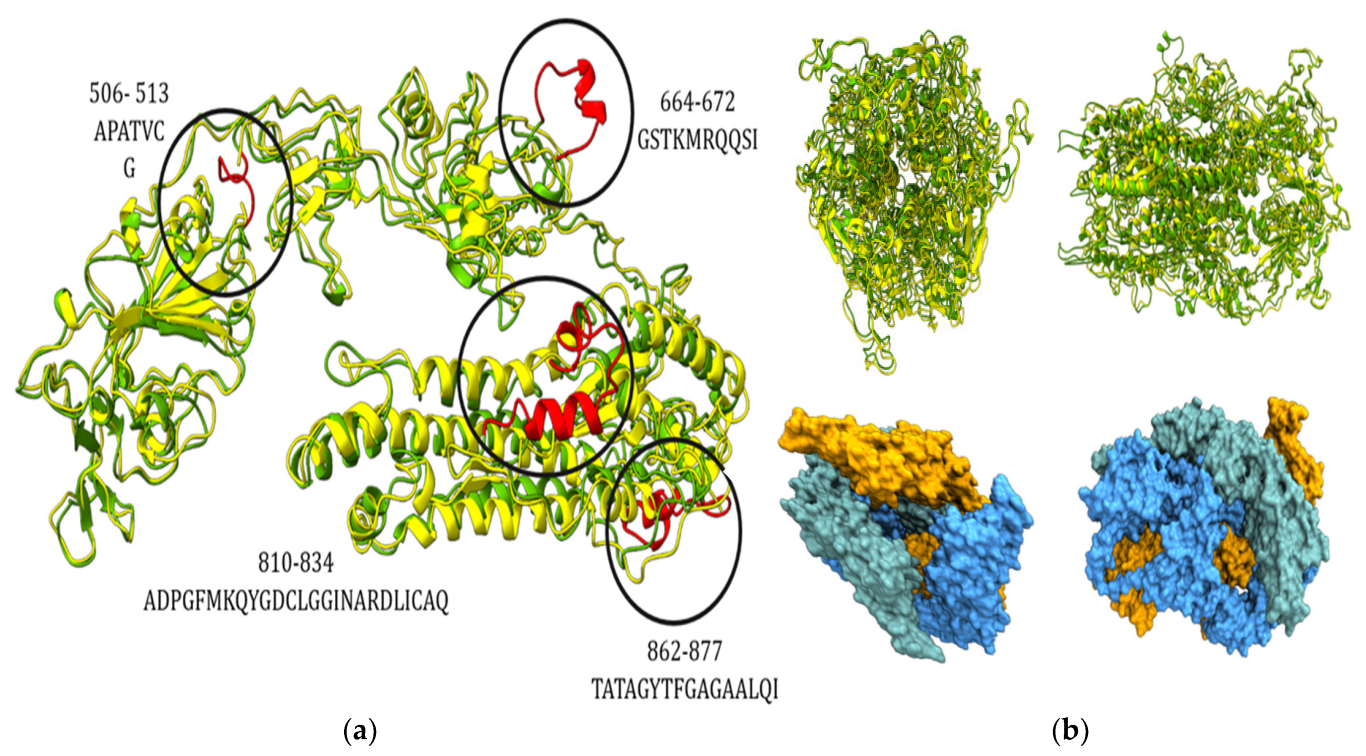
3.3. Genetic and Structural Characterization of the Spike Glycoprotein of Lebanon SARS-Like CoVs and the Potential Significance of Binding to Human ACE2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Delaune, D.; Hul, V.; Karlsson, E.A.; Hassanin, A.; Ou, T.P.; Baidaliuk, A.; Gambaro, F.; Prot, M.; Tu, V.T.; Chea, S.; et al. A novel SARS-CoV-2 related coronavirus in bats from Cambodia. Nat. Commun. 2021, 12, 6563. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Ji, J.; Chen, X.; Bi, Y.; Li, J.; Wang, Q.; Hu, T.; Song, H.; Zhao, R.; Chen, Y.; et al. Identification of novel bat coronaviruses sheds light on the evolutionary origins of SARS-CoV-2 and related viruses. Cell 2021, 184, 4380–4391.e14. [Google Scholar] [CrossRef] [PubMed]
- Wacharapluesadee, S.; Tan, C.W.; Maneeorn, P.; Duengkae, P.; Zhu, F.; Joyjinda, Y.; Kaewpom, T.; Chia, W.N.; Ampoot, W.; Lim, B.L.; et al. Author Correction: Evidence for SARS-CoV-2 related coronaviruses circulating in bats and pangolins in Southeast Asia. Nat. Commun. 2021, 12, 972, Erratum in Nat. Commun. 2021, 12, 1430. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Mendelsohn, E.; Zong, C.; Zhang, W.; Hagan, E.; Wang, N.; Li, S.; Yan, H.; Huang, H.; Zhu, G.; et al. Human-animal interactions and bat coronavirus spillover potential among rural residents in Southern China. Biosaf. Health 2019, 1, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Benda, P.; Abi-Said, M.; Bou Jaoude, I.; Ka, R.; Lučan, R.; Sadek, R.; Ševčík, M.; Uhrin, M.; Horacek, I. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 13. Review of distribution and ectoparasites of bats in Lebanon. Acta Soc. Zool. Bohem. 2016, 80, 207–316. [Google Scholar]
- Abi-Said, M. Monitoring, threats and conservation of hibernating bats roosts in Lebanon. Jordan J. Nat. Hist. 2014, 1, 152–164. [Google Scholar]
- Shehata, M.M.; Chu, D.K.; Gomaa, M.R.; AbiSaid, M.; El Shesheny, R.; Kandeil, A.; Bagato, O.; Chan, S.M.; Barbour, E.K.; Shaib, H.S.; et al. Surveillance for Coronaviruses in Bats, Lebanon and Egypt, 2013–2015. Emerg. Infect. Dis. 2016, 22, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Masangkay, J.S.; Nagata, N.; Morikawa, S.; Mizutani, T.; Fukushi, S.; Alviola, P.; Omatsu, T.; Ueda, N.; Iha, K.; et al. Bat coronaviruses and experimental infection of bats, the Philippines. Emerg. Infect. Dis. 2010, 16, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Gui, M.; Song, W.; Zhou, H.; Xu, J.; Chen, S.; Xiang, Y.; Wang, X. Cryo-electron microscopy structures of the SARS-CoV spike glycoprotein reveal a prerequisite conformational state for receptor binding. Cell Res. 2017, 27, 119–129. [Google Scholar] [CrossRef] [PubMed]
- van Zundert, G.C.P.; Rodrigues, J.P.G.L.M.; Trellet, M.; Schmitz, C.; Kastritis, P.L.; Karaca, E.; Melquiond, A.S.J.; van Dijk, M.; de Vries, S.J.; Bonvin, A.M.J.J. The HADDOCK2.2 Web Server: User-Friendly Integrative Modeling of Biomolecular Complexes. J. Mol. Biol. 2016, 428, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Honorato, R.V.; Koukos, P.I.; Jiménez-García, B.; Tsaregorodtsev, A.; Verlato, M.; Giachetti, A.; Rosato, A.; Bonvin, A.M.J.J. Structural Biology in the Clouds: The WeNMR-EOSC Ecosystem. Front. Mol. Biosci. 2021, 8, 729513. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, X.; Hu, T.; Li, J.; Song, H.; Liu, Y.; Wang, P.; Liu, D.; Yang, J.; Holmes, E.C.; et al. A Novel Bat Coronavirus Closely Related to SARS-CoV-2 Contains Natural Insertions at the S1/S2 Cleavage Site of the Spike Protein. Curr. Biol. 2020, 30, 3896. [Google Scholar] [CrossRef] [PubMed]
- Alkhovsky, S.; Lenshin, S.; Romashin, A.; Vishnevskaya, T.; Vyshemirsky, O.; Bulycheva, Y.; Lvov, D.; Gitelman, A. SARS-like Coronaviruses in Horseshoe Bats (Rhinolophus spp.) in Russia, 2020. Viruses 2022, 14, 113. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Number of Bats (Percentage of Total Bats) | Number Positive (% Samples in Each Category) | p-Value * |
|---|---|---|---|
| Sample type | |||
| Oral | 311 (50%) | 20 (6.4%) | <0.0001 |
| Rectal | 311 (50%) | 57 (18.3%) | |
| Type of bat | |||
| Miniopterus schreibersii | 51 (16.4%) | 12 (23.5%) | <0.0001 |
| Myotis emarginatus | 20 (6.4%) | 2 (10%) | |
| Myotis capaccinii | 6 (1.9%) | 0 (0%) | |
| Myotis sp. | 76 (24.4%) | 0 (0%) | |
| Rhinolophus ferrumequinum | 109 (35.0%) | 40 (36.6%) | |
| Rhinolophus hipposideros | 49 (15.8%) | 3 (6.1%) | |
| Governorates | |||
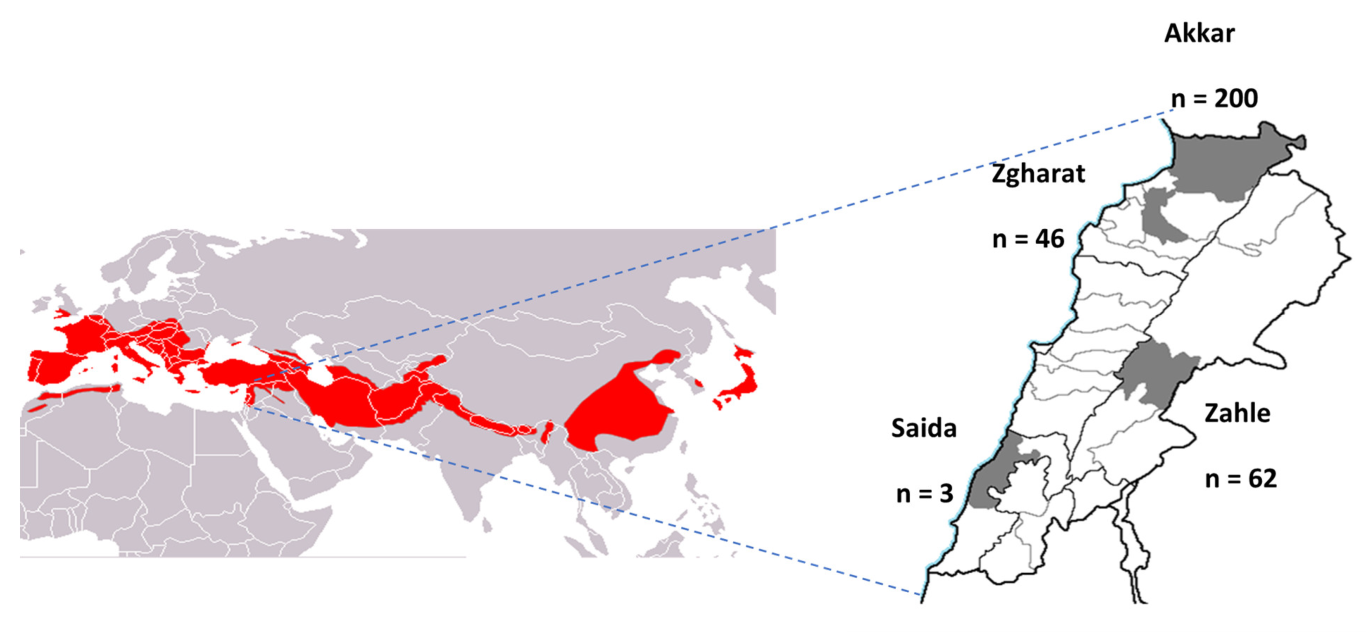
| Akkar | 200 (64.3%) | 43 (21.5%) | <0.0001 |
| Saida | 3 (1.0%) | 3 (100%) | |
| Zahle | 62 (19.9%) | 11 (17.7%) | |
| Zgharat | 46 (14.8%) | 0 (0%) | |
| Gender | |||
| Male | 140 (45%) | 28 (20%) | NS |
| Female | 171 (55%) | 29 (16.9%) | |
| Age | |||
| Adult | 288 (92.6%) | 43 (14.9%) | <0.0001 |
| Juvenile | 23 (7.3%) | 14 (60.8%) |
| Complex | HADDOCK Score | Cluster Size | RMSD from the Overall Lowest-Energy Structure | Z-Score | Van der Waals Energy |
|---|---|---|---|---|---|
| CNT | −104.2 ± 6.7 | 31 | 1.3 ± 1.1 | −1.7 | −83.0 ± 1.8 |
| Novel | −206.6 ± 15.9 | 15 | 0.5 ± 0.4 | −2.4 | −81.0 ± 12.9 |
| Wild | ACE2 | Distance (Å) | Novel | ACE2 | Distance (Å) | ||
|---|---|---|---|---|---|---|---|
| Chain C | Thr320 | Asp597 | 3.01 | Chain C | Asn324 | Glu564 | 3.00 |
| Asn321 | Lys596 | 3.12 | Cys326 | Gln388 | 3.17 | ||
| Leu322 | Lys596 | 3.08 | Gln330 | Gln388 | 3.50 | ||
| Asn330 | Glu238 | 2.93 | Gln330 | Pro389 | 3.35 | ||
| Asp351 | Asn601 | 2.91 | Gln330 | Arg559 | 2.53 | ||
| Thr359 | Asp615 | 3.13 | Ser349 | Gln89 | 2.71 | ||
| Chain B | Tyr440 | Pro253 | 2.82 | Cys351 | Glu22 | 3.35 | |
| Asn479 | Asp157 | 3.08 | Cys351 | Asn90 | 3.81 | ||
| Asn479 | Asn159 | 3.29 | Asp354 | Lys26 | 3.19 | ||
| Asp480 | Tyr255 | 2.79 | Asp354 | Asn90 | 2.84 | ||
| Thr487 | Tyr613 | 3.23 | Tyr359 | Asp30 | 2.91 | ||
| Gly488 | Ala614 | 3.44 | Asp486 | Gln325 | 2.92 | ||
| Chain C | Lys514 | Gln598 | 2.70 | Cys512 | Gln89 | 2.72 | |
| Lys514 | Asn601 | 2.57 | Chain B | Ile432 | Lys68 | 2.45 | |
| Lys434 | Glu75 | 2.75 | |||||
| Ala462 | Met82 | 3.29 | |||||
| Leu473 | Gln81 | 3.19 | |||||
| Leu473 | Met82 | 3.77 | |||||
| Cys475 | Leu79 | 3.55 | |||||
| Cys475 | Gln81 | 3.55 | |||||
| Tyr476 | Glu75 | 2.77 | |||||
| Tyr482 | Glu35 | 3.07 | |||||
| Simulation Time (ns) | 0 | 50 | 100 | 150 | 200 | 250 | 300 | 350 | 400 | 450 | 500 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| H-bonds | 20 | 15 | 19 | 22 | 20 | 21 | 22 | 20 | 19 | 22 | 22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kandeil, A.; Abi-Said, M.; Badra, R.; El-Shesheny, R.; Al-Karmalawy, A.A.; Alnajjar, R.; Khalid, Z.; Kamel, M.N.; Abi Habib, W.; Abdallah, J.; et al. Detection of Coronaviruses in Bats in Lebanon during 2020. Pathogens 2023, 12, 876. https://doi.org/10.3390/pathogens12070876
Kandeil A, Abi-Said M, Badra R, El-Shesheny R, Al-Karmalawy AA, Alnajjar R, Khalid Z, Kamel MN, Abi Habib W, Abdallah J, et al. Detection of Coronaviruses in Bats in Lebanon during 2020. Pathogens. 2023; 12(7):876. https://doi.org/10.3390/pathogens12070876
Chicago/Turabian StyleKandeil, Ahmed, Mounir Abi-Said, Rebecca Badra, Rabeh El-Shesheny, Ahmed A. Al-Karmalawy, Radwan Alnajjar, Zumama Khalid, Mina Nabil Kamel, Walid Abi Habib, Jad Abdallah, and et al. 2023. "Detection of Coronaviruses in Bats in Lebanon during 2020" Pathogens 12, no. 7: 876. https://doi.org/10.3390/pathogens12070876
APA StyleKandeil, A., Abi-Said, M., Badra, R., El-Shesheny, R., Al-Karmalawy, A. A., Alnajjar, R., Khalid, Z., Kamel, M. N., Abi Habib, W., Abdallah, J., Dhanasekaran, V., Webby, R., & Kayali, G. (2023). Detection of Coronaviruses in Bats in Lebanon during 2020. Pathogens, 12(7), 876. https://doi.org/10.3390/pathogens12070876