


Evaluation of Entomopathogenic Nematodes against Common Wireworm Species in Potato Cultivation


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Nematodes
2.2. Source of Wireworms
2.3. Evaluation of the Pathogenicity of EPNs in Pot Experiment
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Furlan, L.; Contiero, B.; Chiarini, F.; Colauzzi, M.; Sartori, E.; Benvegnù, I.; Fracasso, F.; Giandon, P. Risk assessment of maize damage by wireworms (Coleoptera: Elateridae) as the first step in implementing IPM and in reducing the environmental impact of soil insecticides. Environ. Sci. Pollut. Res. 2017, 24, 236–251. [Google Scholar] [CrossRef] [PubMed]
- Vernon, R.S.; van Herk, W.G. Wireworms as Pests of Potato. In Insect Pests of Potato: Global Perspectives on Biology and Management; Alyokhin, A., Vincent, C., Giordanengo, P., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2013; pp. 103–164. [Google Scholar]
- Chalfant, R.B.; Seal, D.R. Biology and management of wireworms on sweet potato. In Sweet Potato Pest Management; Jansson, R.K., Raman, K.V., Eds.; CRC Press: Boca Raton, FA, USA, 2019; pp. 303–326. [Google Scholar] [CrossRef]
- Keiser, A.; Häberli, M.; Stamp, P. Quality deficiencies on potato (Solanum tuberosum L.) tubers caused by Rhizoctonia solani, wireworms (Agriotes ssp.) and slugs (Deroceras reticulatum, Arion hortensis) in different farming systems. Field Crop. Res. 2012, 128, 147–155. [Google Scholar] [CrossRef]
- Jansson, R.K.; Lecrone, S.H. Effects of summer cover crop management on wireworm (Coleoptera: Elateridae) abundance and damage to potato. J. Econ. Entomol. 1991, 84, 581–586. [Google Scholar] [CrossRef]
- Nikoukar, A.; Rashed, A. Integrated Pest Management of Wireworms (Coleoptera: Elateridae) and the Rhizosphere in Agroecosystems. Insects 2022, 13, 769. [Google Scholar] [CrossRef]
- Kroschel, J.; Mujica, N.; Okonya, J.; Alyokhin, A. Insect pests affecting potatoes in tropical, subtropical, and temperate regions. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Springer: New York, NY, USA, 2020; pp. 80–123. [Google Scholar] [CrossRef]
- Furlan, L.; Tóth, M. Occurrence of click beetle pest spp. (Coleoptera, Elateridae) in Europe as detected by pheromone traps: Survey results of 1998–2006. IOBC/WPRS Bull. 2007, 30, 19. [Google Scholar]
- Furlan, L.; Benvegnù, I.; Bilò, M.F.; Lehmhus, J.; Ruzzier, E. Species Identification of Wireworms (Agriotes spp.; Coleoptera: Elateridae) of Agricultural Importance in Europe: A New “Horizontal Identification Table”. Insects 2021, 12, 534. [Google Scholar] [CrossRef]
- Poggi, S.; Le Cointe, R.; Lehmhus, J.; Plantegenest, M.; Furlan, L. Alternative strategies for controlling wireworms in field crops: A review. Agriculture 2021, 11, 436. [Google Scholar] [CrossRef]
- Reddy, G.V.; Tangtrakulwanich, K.; Wu, S.; Miller, J.H.; Ophus, V.L.; Prewett, J.; Jaronski, S.T. Evaluation of the effectiveness of entomopathogens for the management of wireworms (Coleoptera: Elateridae) on spring wheat. J. Invertebr. Pathol. 2014, 120, 43–49. [Google Scholar] [CrossRef]
- Eckard, S.; Ansari, M.A.; Bacher, S.; Butt, T.M.; Enkerli, J.; Grabenweger, G. Virulence of in vivo and in vitro produced conidia of Metarhizium brunneum strains for control of wireworms. J. Crop. Prot. 2014, 64, 137–142. [Google Scholar] [CrossRef]
- Furlan, L.; Contiero, B.; Chiarini, F.; Benvegnù, I.; Tóth, M. The use of click beetle pheromone traps to optimize the risk assessment of wireworm (Coleoptera: Elateridae) maize damage. Sci. Rep. 2020, 10, 8780. [Google Scholar] [CrossRef]
- La Forgia, D.; Bruno, P.; Campos-Herrera, R.; Turlings, T.; Verheggen, F. The lure of hidden death: Development of an attract-and-kill strategy against Agriotes obscurus (Coleoptera: Elateridae) combining semiochemicals and entomopathogenic nematodes. Turk. J. Zool. 2021, 45, 347–355. [Google Scholar] [CrossRef]
- Gulzar, S.; Wakil, W.; Shapiro-Ilan, D.I. Potential use of entomopathogenic nematodes against the soil dwelling stages of onion thrips, Thrips tabaci Lindeman: Laboratory, greenhouse and field trials. Biol. Control 2021, 161, 104677. [Google Scholar] [CrossRef]
- Vicente-Díez, I.; Blanco-Pérez, R.; Chelkha, M.; Puelles, M.; Pou, A.; Campos-Herrera, R. Exploring the use of entomopathogenic nematodes and the natural products derived from their symbiotic bacteria to control the grapevine moth, Lobesia botrana (Lepidoptera: Tortricidae). Insects 2021, 12, 1033. [Google Scholar] [CrossRef] [PubMed]
- Peçen, A.; Kepenekci, İ. Efficacy of entomopathogenic nematode isolates from Turkey against wheat stink bug, Aelia rostrata Boheman (Hemiptera: Pentatomidae) adults under laboratory conditions. Egypt. J. Biol. Pest Control 2022, 32, 1–6. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Hazir, S.; Glazer, I. Entomopathogenic Nematodes as Models for Inundative Biological Control. In Nematodes as Model Organisms; Glazer, I., Shapiro-Ilan, D.I., Sternberg, P.W., Eds.; CABI International: Boston, MA, USA, 2022; pp. 293–308. [Google Scholar]
- Usman, M.; Wakil, W.; Shapiro-Ilan, D.I. Entomopathogenic nematodes as biological control agent against Bactrocera zonata and Bactrocera dorsalis (Diptera: Tephritidae). Biol. Control 2021, 163, 104706. [Google Scholar] [CrossRef]
- Kaya, H.K.; Gaugler, R. Entomopathogenic nematodes. Annu. Rev. Entomol. 1993, 38, 181–206. [Google Scholar] [CrossRef]
- Koppenhöfer, A.M.; Shapiro-Ilan, D.I.; Hiltpold, I. Entomopathogenic nematodes in sustainable food production. Front. Sustain. Food Syst. 2020, 4, 125. [Google Scholar] [CrossRef]
- Stock, S.P. Partners in crime: Symbiont-assisted resource acquisition in Steinernema entomopathogenic nematodes. Curr. Opin. Insect Sci. 2019, 32, 22–27. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M. Xenorhabdus spp.: An overview of the useful facets of mutualistic bacteria of entomopathogenic nematodes. Life 2022, 12, 1360. [Google Scholar] [CrossRef]
- Hazir, S.; Kaya, H.K.; Stock, S.P.; Keskin, N. Entomopathogenic nematodes (Steinernematidae and Heterorhabditidae) for biological control of soil pests. Turk. J. Biol. 2003, 27, 181–202. [Google Scholar]
- Shapiro-Ilan, D.; Dolinski, C. Entomopathogenic nematode application technology. In Nematode Pathogenesis of Insects and Other Pests; Campos-Herrera, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 231–254. [Google Scholar] [CrossRef]
- Grewal, P.S.; Selvan, S.; Gaugler, R. Nematodes: Niche breadth for infection, establishment, and reproduction. J. Therm. Biol. 1994, 19, 245–253. [Google Scholar]
- Lewis, E.E.; Campbell, J.; Griffin, C.; Kaya, H.; Peters, A. Behavioral ecology of entomopathogenic nematodes. Biol. Control 2006, 38, 66–79. [Google Scholar] [CrossRef]
- Noujeim, E.; Rehayem, M.; Nemer, N. Comparison of indigenous and exotic entomopathogenic nematode strains for control of the cedar web-spinning sawfly, Cephalcia tannourinensis in vitro. Biocontrol Sci. Technol. 2015, 25, 843–851. [Google Scholar] [CrossRef]
- Sandhi, R.K.; Reddy, G.V. Effects of entomopathogenic nematodes and symbiotic bacteria on non-target arthropods. In Microbes for Sustainable Insect Pest Management; Khan, A.M., Ahmad, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 247–273. [Google Scholar] [CrossRef]
- Gümüş Askar, A.; Yüksel, E.; Öcal, A.; Özer, G.; Kütük, H.; Dababat, A.; İmren, M. Identification and control potential of entomopathogenic nematodes against the black cutworm, Agrotis ipsilon (Fabricius) (Lepidoptera: Noctuidae), in potato-growing areas of Turkey. J. Plant Dis. Prot. 2022, 129, 911–922. [Google Scholar] [CrossRef]
- Gümüş Askar, A.; Yüksel, E.; Dinçer, D.; Bozbuğa, R.; Öcal, A.; Kütük, H.; Dababat, A.A.; Özer, G.; İmren, M. Molecular Identification, Occurrence and Biodiversity of Wireworm Species (Agriotes spp.) (Coleoptera: Elateridae) in Major Potato Cultivated Areas of Türkiye. J. Insect Biodivers. 2023; submitted. [Google Scholar]
- Kaya, H.K.; Stock, S.P. Techniques in insect nematology. In Manual of Techniques in Insect Pathology, 1st ed.; Lacey, L.A., Ed.; Academic Press: Cambridge, MA, USA, 1997; pp. 281–324. [Google Scholar] [CrossRef]
- Metwally, H.M.; Hafez, G.A.; Hussein, M.A.; Hussein, M.A.; Salem, H.A.; Saleh, M.M.E. Low cost artificial diet for rearing the greater wax moth, Galleria mellonella L. (Lepidoptera: Pyralidae) as a host for entomopathogenic nematodes. Egypt. J. Biol. Pest Control 2012, 22, 15. [Google Scholar]
- Ansari, M.A.; Evans, M.; Butt, T.M. Identification of pathogenic strains of entomopathogenic nematodes and fungi for wireworm control. Crop Prot. 2009, 28, 269–272. [Google Scholar] [CrossRef]
- Sufyan, M.; Neuhoff, D.; Furlan, L. Larval development of Agriotes obscurus under laboratory and semi-natural conditions. Bull. Insectol. 2014, 67, 227–235. [Google Scholar]
- Ritter, C.; Richter, E. Control methods and monitoring of Agriotes wireworms (Coleoptera: Elateridae). J. Plant Dis. Prot. 2013, 120, 4–15. [Google Scholar]
- Öğretmen, A.; Yüksel, E.; Canhilal, R. Susceptibility of larvae of wireworms (Agriotes spp.)(Coleoptera: Elateridae) to some Turkish isolates of entomopathogenic nematodes under laboratory and field conditions. Biol. Control 2020, 149, 104320. [Google Scholar] [CrossRef]
- Williams, L.; Cherry, R.; Shapiro-Ilan, D. Effect of Host Size on Susceptibility of Melanotus communis (Coleoptera: Elateridae) Wireworms to Entomopathogens. J. Nematol. 2022, 54, 10.2478/jofnem-2022-0033. [Google Scholar] [CrossRef]
- La Forgia, D.; Jaffuel, G.; Campos-Herrera, R.; Verheggen, F.; Turlings, T. Efficiency of an Attract-and-Kill System with Entomopathogenic Nematodes against Wireworms (Coleoptera: Elateridae). IOBC/WPRS Bull. 2020, 150, 91–95. [Google Scholar]
- Campos-Herrera, R.; Gutiérrez, C. Screening Spanish Isolates of Steinernematid Nematodes for Use as Biological Control Agents through Laboratory and Greenhouse Microcosm Studies. J. Invertebr. Pathol. 2009, 100, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Toba, H.H.; Lindegren, J.E.; Turner, J.E.; Vail, P.V. Susceptibility of the Colorado Potato Beetle and the Sugarbeet Wireworm to Steinernema feltiae and S. glaseri. J. Nematol. 1983, 15, 597–601. [Google Scholar] [PubMed]
- Morton, A.; Garcia-del-Pino, F. Laboratory and Field Evaluation of Entomopathogenic Nematodes for Control of Agriotes obscurus (L.) (Coleoptera: Elateridae). J. Appl. Entomol. 2017, 141, 241–246. [Google Scholar] [CrossRef]
- Sandhi, R.K.; Shapiro-Ilan, D.; Sharma, A.; Reddy, G.V. Efficacy of entomopathogenic nematodes against the sugarbeet wireworm, Limonius californicus (Mannerheim) (Coleoptera: Elateridae). Biol. Control 2020, 143, 104190. [Google Scholar] [CrossRef]
- Abbas, M.S.T. Pathogenicity of entomopathogenic nematodes to dipteran leaf miners, house flies and mushroom flies. Egypt. J. Biol. Pest Control 2022, 32, 76. [Google Scholar] [CrossRef]
- Yüksel, E. Biocontrol potential of endosymbiotic bacteria of entomopathogenic nematodes against the tomato leaf miner, Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae). Egypt. J. Biol. Pest Control 2022, 32, 135. [Google Scholar] [CrossRef]
- Köhl, J.; Postma, J.; Nicot, P.; Ruocco, M.; Blum, B. Stepwise screening of microorganisms for commercial use in biological control of plant-pathogenic fungi and bacteria. Biol. Control 2011, 57, 1–12. [Google Scholar] [CrossRef]
- Simoes, N.; Rosa, J.S. Pathogenicity and host specificity of entomopathogenic nematodes. Biocontrol Sci. Technol. 1996, 6, 403–412. [Google Scholar] [CrossRef]
- Alonso, V.; Nasrolahi, S.; Dillman, A.R. Host-specific activation of entomopathogenic nematode infective juveniles. Insects 2018, 9, 59. [Google Scholar] [CrossRef]
- Zhang, X.; Li, L.; Kesner, L.; Robert, C.A.M. Chemical host-seeking cues of entomopathogenic nematodes. Curr. Opin. Insect Sci. 2021, 44, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Eidt, D.C.; Thurston, G.S. Physical deterrents to infection by entomopathogenic nematodes in wireworms (Coleoptera: Elateridae) and other soil insects. Can. Entomol. 1995, 127, 423–429. [Google Scholar] [CrossRef]
- Rahatkhah, Z.; Karimi, J.; Ghadamyari, M.; Brivio, M.F. Immune defenses of Agriotes lineatus larvae against entomopathogenic nematodes. BioControl 2015, 60, 641–653. [Google Scholar] [CrossRef]
- Bode, H.B.; Müller, R. The impact of bacterial genomics on natural product research. Angew. Chem. Int. Ed. 2005, 44, 6828–6846. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B. Entomopathogenic bacteria as a source of secondary metabolites. Curr. Opin. Chem. Biol. 2009, 13, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Shawer, R.; Donati, I.; Cellini, A.; Spinelli, F.; Mori, N. Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii. Insects 2018, 9, 148. [Google Scholar] [CrossRef]
- Tobias, N.J.; Shi, Y.M.; Bode, H.B. Refining the natural product repertoire in entomopathogenic bacteria. Trends Microbiol. 2018, 26, 833–840. [Google Scholar] [CrossRef]
- Hasan, M.A.; Ahmed, S.; Mollah, M.M.I.; Lee, D.; Kim, Y. Variation in pathogenicity of different strains of Xenorhabdus nematophila; Differential immunosuppressive activities and secondary metabolite production. J. Invertebr. Pathol. 2019, 166, 107221. [Google Scholar] [CrossRef]
- Feldhaar, H.; Gross, R. Immune reactions of insects on bacterial pathogens and mutualists. Microbes Infect. 2008, 10, 1082–1088. [Google Scholar] [CrossRef]
- Abdolmaleki, A.; Rafiee Dastjerdi, H.; Tanha Maafi, Z.; Naseri, B. Virulence of two entomopathogenic nematodes through their interaction with Beauveria bassiana and Bacillus thuringiensis against Pieris brassicae (Lepidoptera: Pieridae). J. Crop Prot. 2017, 6, 287–299. [Google Scholar]
- Ebssa, L.; Koppenhöfer, A.M. Efficacy and persistence of entomopathogenic nematodes for black cutworm control in turfgrass. Biocontrol Sci. Technol. 2011, 21, 779–796. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Gouge, D.H.; Piggott, S.J.; Fife, J.P. Application technology and environmental considerations for use of entomopathogenic nematodes in biological control. Biol. Control 2006, 38, 124–133. [Google Scholar] [CrossRef]
- Gulzar, S.; Usman, M.; Wakil, W.; Gulcu, B.; Hazir, C.; Karagoz, M.; Shapiro-Ilan, D.I. Environmental tolerance of entomopathogenic nematodes differs among nematodes arising from host cadavers versus aqueous suspension. J. Invertebr. Pathol. 2020, 175, 107452. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, W.F.; All, J.N. Effects of temperature and placement site on the dispersal of the entomopathogenic nematode, Steinernema carpocapsae in four soils. Chin. J. Entomol. 1996, 16, 95–106. [Google Scholar]
- Hummel, R.L.; Walgenbach, J.F.; Barbercheck, M.E.; Kennedy, G.G.; Hoyt, G.D.; Arellano, C. Effects of production practices on soil-borne entomopathogens in western North Carolina vegetable systems. Environ. Entomol. 2002, 31, 84–91. [Google Scholar] [CrossRef]
- Shaurub, E.H.; Soliman, N.A.; Hashem, A.G.; Abdel-Rahman, A.M. Infectivity of four entomopathogenic nematodes in relation to environmental factors and their effects on the biochemistry of the Medfly Ceratitis capitata (Wied.)(Diptera: Tephritidae). Neotrop. Entomol. 2015, 44, 610–618. [Google Scholar]
- Hirao, A.; Ehlers, R.U. Effect of temperature on the development of Steinernema carpocapsae and Steinernema feltiae (Nematoda: Rhabditida) in liquid culture. Appl. Microbiol. Biotechnol. 2009, 84, 1061–1067. [Google Scholar]
- Chung, H.J.; Lee, D.W.; Yoon, H.S.; Lee, S.M.; Park, C.G.; Choo, H.Y. Temperature and dose effects on the pathogenicity and reproduction of two Korean isolates of Heterorhabditis bacteriophora (Nematoda: Heterorhabditidae). J. Asia Pac. Entomol. 2010, 13, 277–282. [Google Scholar] [CrossRef]
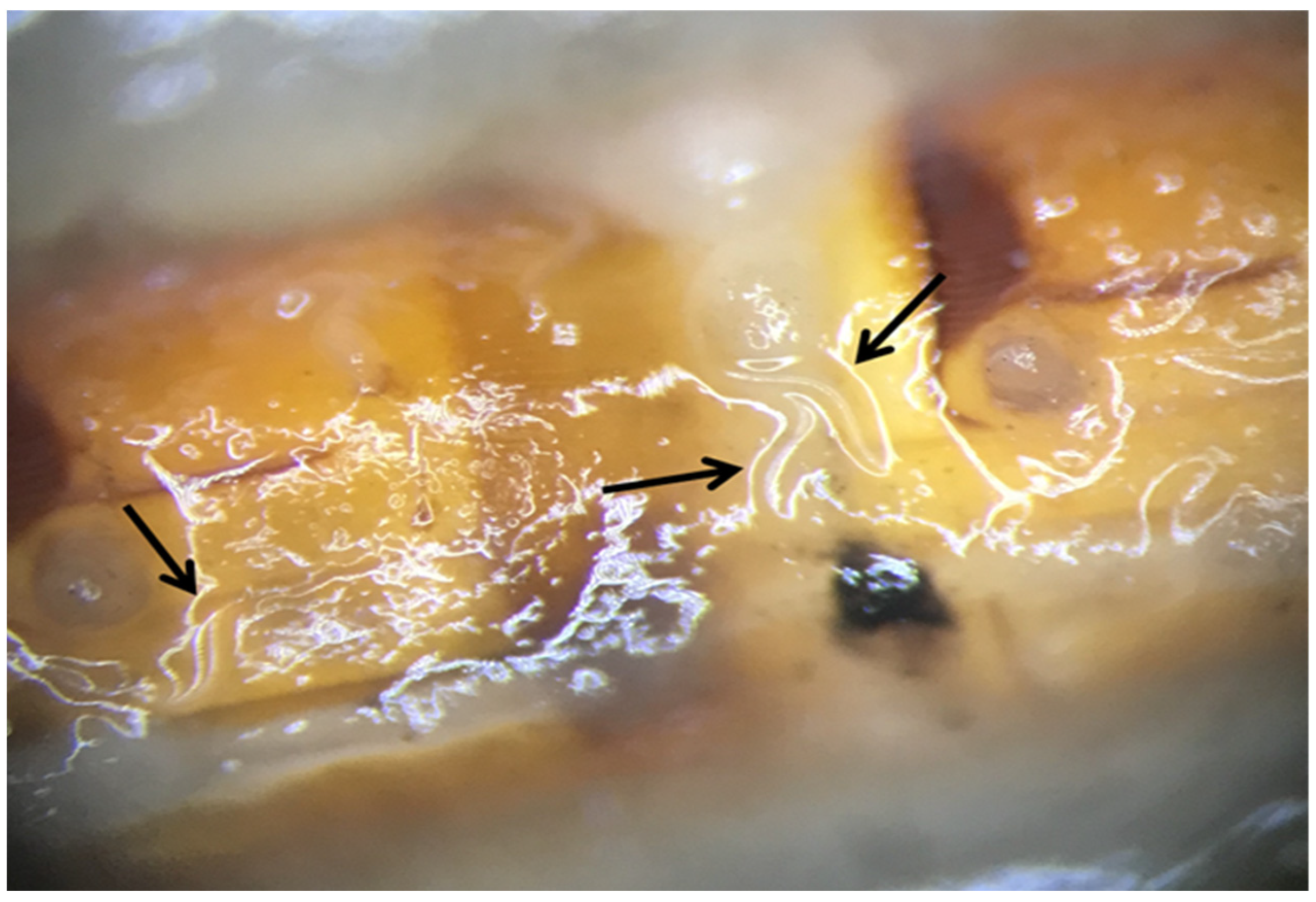

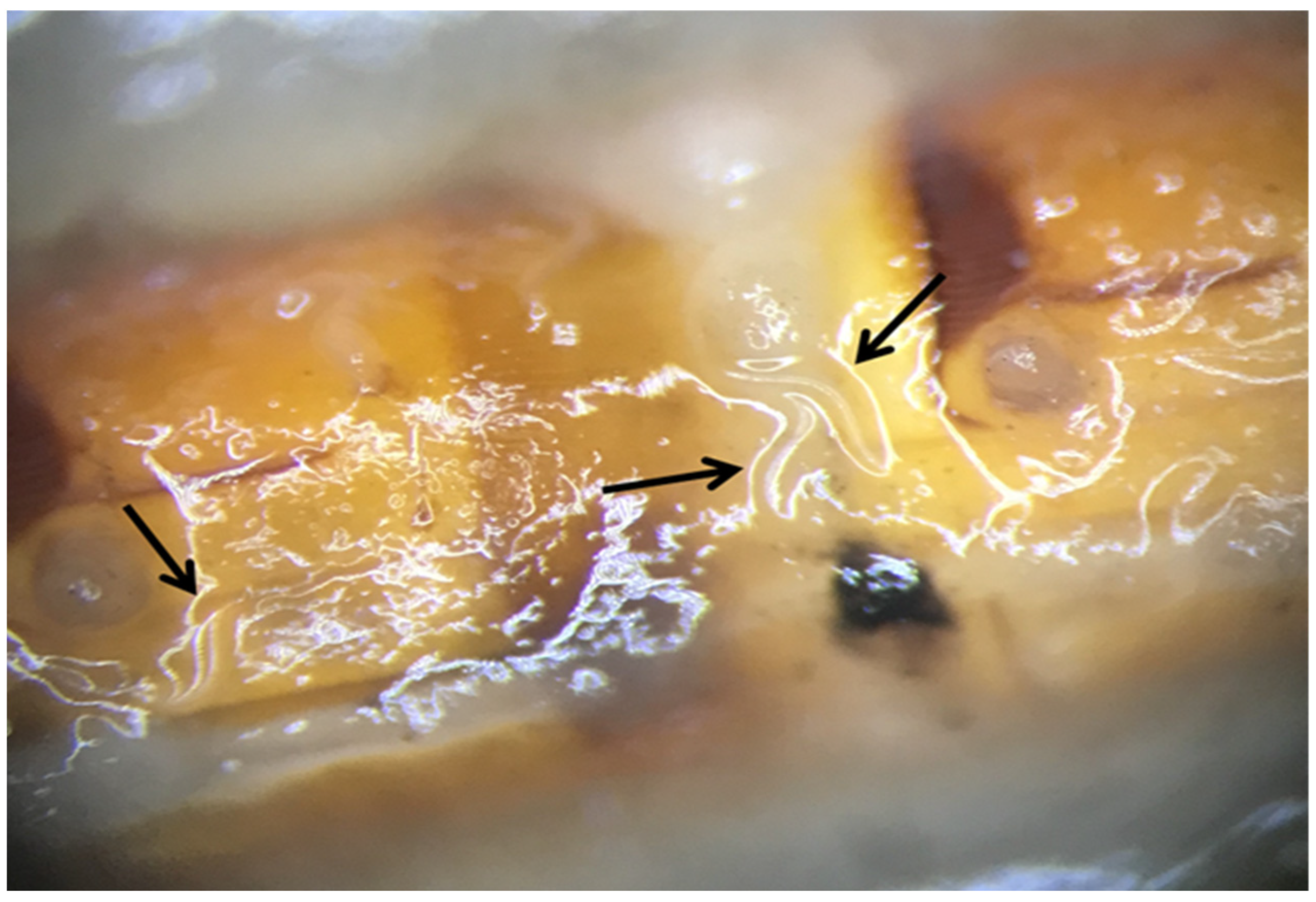{kind=link}
| Entomopathogenic Nematodes | Strain | Habitat | Coordinates | GenBank Accession Number |
|---|---|---|---|---|
| Steinernema carpocapsae | Sc_BL22 | Potato | 40°47′11″ N 31°38′78″ E | OK632299 |
| Steinernema feltiae | Sf_BL24 | Potato | 40°47′14″ N 31°39′10″ E | OK632300 |
| Steinernema feltiae | Sf_KAY4 | Potato | 38°20′28″ N 35°27′49″ E | OK632306 |
| Heterorhabditis bacteriophora | Hb_N3 | Potato | 38°02′19″ N 34°44′18″ E | OK632328 |
| Heterorhabditis bacteriophora | Hb_KAY10 | Potato | 38°16′49″ N 35°25′15″ E | OK632308 |
| Heterorhabditis bacteriophora | Hb_AF12 | Potato | 37°55′31″ N 29°52′19″ E | OK632288 |
| Sources | Agriotes rufipalpis | Agriotes sputator | |||
|---|---|---|---|---|---|
| Degree of Freedom | F Value | p Value | F Value | p Value | |
| Nematode (N) | 5 | 9.268 | <0.01 | 17.455 | <0.01 |
| Concentration (C) | 5 | 295.432 | <0.01 | 825.869 | <0.01 |
| Temperature (T) | 1 | 42.688 | <0.01 | 57.436 | <0.01 |
| C × N | 25 | 1.735 | 0.020 | 3.327 | <0.01 |
| C × T | 5 | 15.012 | <0.01 | 5.465 | <0.01 |
| N × T | 5 | 2.746 | 0.020 | 2.042 | 0.074 |
| C × N × T | 25 | 1.554 | 0.051 | 4.895 | <0.01 |
| Error1 | 216 | ||||
| Exposure time (t) | 1 | 491.683 | <0.01 | 337.559 | <0.01 |
| t × C | 5 | 94.844 | <0.01 | 28.474 | <0.01 |
| t × N | 5 | 1.225 | 0.298 | 2.961 | 0.013 |
| t × T | 1 | 77.778 | 0.540 | 3.194 | 0.075 |
| t × C × N | 25 | 0.799 | 0.742 | 1.457 | 0.081 |
| t × C × T | 5 | 5.321 | 0.358 | 1.636 | 0.152 |
| t × N × T | 5 | 5.054 | 0.011 | 3.206 | 0.008 |
| t × C × N × T | 25 | 1.717 | 0.022 | 1.340 | 0.137 |
| Error2 | 216 | ||||
| Temperatures | Nematodes * | Mortality Rates (%) 6 Days after Treatment (DAT) | ||||
|---|---|---|---|---|---|---|
| Control | 25 IJs/cm2 | 50 IJs/cm2 | 100 IJs/cm2 | 150 IJs/cm2 | ||
| 25 °C | Sc_BL22 | 0.0 ± 0.0A a a b | 7.5 ± 5.0Ba | 7.5 ± 5.0Ba | 10.0 ± 8.1Ba | 12.5 ± 5.0Ba |
| Sf_BL24 | 0.0 ± 0.0Aa | 2.5 ± 5.0Aa | 5.0 ± 5.7Aa | 15.0 ± 5.7Ba | 25.0 ± 5.7Cb | |
| Sf_KAY4 | 0.0 ± 0.0Aa | 2.5 ± 5.0Aa | 5.0 ± 5.7Aa | 25.0 ± 5.7Bb | 25.0 ± 5.7Bb | |
| Hb_N3 | 0.0 ± 0.0Aa | 2.5 ± 5.0Aa | 2.5 ± 5.0Aa | 15.0 ± 10.0Ba | 27.5 ± 9.5Cb | |
| Hb_KAY10 | 0.0 ± 0.0Aa | 5.0 ± 5.7Aa | 5.0 ± 10.0Aa | 15.0 ± 10.0Ba | 20.0 ± 8.1Bab | |
| Hb_AF12 | 0.0 ± 0.0Aa | 7.5 ± 5.0Ba | 7.5 ± 9.5Ba | 15.0 ± 5.7Ca | 15.0 ± 5.7Ca | |
| 30 °C | Sc_BL22 | 0.0 ± 0.0Aa | 12.5 ± 5.0Ba | 12.5 ± 9.5Ba | 15.0 ± 8.1Ba | 22.5 ± 5.0Ca |
| Sf_BL24 | 0.0 ± 0.0Aa | 7.5 ± 9.5Ba | 17.5 ± 5.0Cab | 30.0 ± 8.1Db | 32.5 ± 5.0Dab | |
| Sf_KAY4 | 0.0 ± 0.0Aa | 15.0 ± 5.7Ba | 25.0 ± 5.7Cb | 27.5 ± 5.0Cb | 32.5 ± 5.0Dab | |
| Hb_N3 | 0.0 ± 0.0Aa | 2.5 ± 5.0Aa | 10.0 ± 11.5Ba | 25.0 ± 12.9Cb | 27.5 ± 9.5Ca | |
| Hb_KAY10 | 0.0 ± 0.0Aa | 10.0 ± 0.0Ba | 10.0 ± 0.0Ba | 12.5 ± 5.0Ba | 25.0 ± 10.0Ca | |
| Hb_AF12 | 0.0 ± 0.0Aa | 15.0 ± 10.0Ba | 20.0 ± 0.0Bb | 25.0 ± 5.7Bb | 37.5 ± 5.0Cb | |
| Temperatures | Nematodes * | Mortality Rates (%) 18 Days after Treatment (DAT) | ||||
|---|---|---|---|---|---|---|
| Control | 25 Ijs/cm2 | 50 Ijs/cm2 | 100 Ijs/cm2 | 150 Ijs/cm2 | ||
| 25 °C | Sc_BL22 | 0.0 ± 0.0A a a b | 30.0 ± 0.0Bb | 45.0 ± 10.0Bab | 82.5 ± 5.0Cba | 85.0 ± 11.5Cb |
| Sf_BL24 | 0.0 ± 0.0Aa | 25.0 ± 5.7Bab | 37.5 ± 5.0Ba | 77.5 ± 5.0Ca | 85.0 ± 12.9Cb | |
| Sf_KAY4 | 0.0 ± 0.0Aa | 25.0 ± 5.7Bab | 37.5 ± 5.0Ba | 80.5 ± 15.0Ca | 85.0 ± 5.7Cb | |
| Hb_N3 | 0.0 ± 0.0Aa | 25.0 ± 5.7Bab | 55.5 ± 5.0Cb | 77.5 ± 5.0Da | 80.0 ± 8.1Da | |
| Hb_KAY10 | 0.0 ± 0.0Aa | 20.0 ± 8.1Ba | 60.0 ± 8.1Cb | 80.0 ± 5.7Ca | 80.5 ± 12.5Da | |
| Hb_AF12 | 0.0 ± 0.0Aa | 25.0 ± 5.7Bab | 50.0 ± 0.0Cab | 72.5 ± 5.0Da | 85.0 ± 12.9Db | |
| 30 °C | Sc_BL22 | 0.0 ± 0.0Aa | 30.0 ± 0.0Ba | 55.0 ± 10.0Ca | 82.5 ± 5.0Bda | 85.0 ± 11.5Da |
| Sf_BL24 | 0.0 ± 0.0Aa | 35.0 ± 5.7Aa | 67.5 ± 5.0Bab | 87.5 ± 5.0Ca | 87.5 ± 10.5Ca | |
| Sf_KAY4 | 0.0 ± 0.0Aa | 25.0 ± 5.7Aa | 67.5 ± 5.0Bab | 82.5 ± 15.0Ca | 85.0 ± 5.7Ca | |
| Hb_N3 | 0.0 ± 0.0Aa | 35.0 ± 5.7Ba | 65.5 ± 5.0Bab | 80.5 ± 5.0Ca | 80.0 ± 8.1Ca | |
| Hb_KAY10 | 0.0 ± 0.0Aa | 40.0 ± 8.1Cab | 75.0 ± 8.1Db | 80.0 ± 5.7Da | 82.5 ± 12.5Da | |
| Hb_AF12 | 0.0 ± 0.0Aa | 45.0 ± 5.7Cb | 75.0 ± 0.0Db | 85.5 ± 5.0Da | 85.0 ± 12.9Da | |
| Temperatures | Nematodes * | Mortality Rates (%) 6 Days after Treatment (DAT) | ||||
|---|---|---|---|---|---|---|
| Control | 25 Ijs/cm2 | 50 Ijs/cm2 | 100 Ijs/cm2 | 150 Ijs/cm2 | ||
| 25 °C | Sc_BL22 | 0.0 ± 0.0A a a b | 5.0 ± 5.7Aa | 10.0 ± 0.0Aba | 12.5 ± 9.5Aba | 17.5 ± 5.0Ba |
| Sf_BL24 | 0.0 ± 0.0Aa | 0.0 ± 0.0Aa | 7.5 ± 9.5Ba | 27.5 ± 5.0Cab | 27.5 ± 5.0Cab | |
| Sf_KAY4 | 0.0 ± 0.0Aa | 2.5 ± 5.0Aa | 5.0 ± 5.7Aa | 25.0 ± 5.7Bab | 42.5 ± 5.0Cb | |
| Hb_N3 | 0.0 ± 0.0Aa | 7.5 ± 9.5Ba | 17.5 ± 5.0Cab | 20.0 ± 0.0Cab | 37.5 ± 5.0Db | |
| Hb_KAY10 | 0.0 ± 0.0Aa | 0.0 ± 0.0Aa | 12.5 ± 9.5Bab | 40.0 ± 8.1Cb | 47.5 ± 9.5Cb | |
| Hb_AF12 | 0.0 ± 0.0Aa | 7.5 ± 9.5Ba | 15.0 ± 5.7BCb | 32.5 ± 5.0Cab | 45.0 ± 5.7Db | |
| 30 °C | Sc_BL22 | 0.0 ± 0.0Aa | 5.0 ± 5.7Aa | 15.0 ± 5.7Ba | 32.5 ± 5.0Cb | 50.0 ± 8.1Db |
| Sf_BL24 | 0.0 ± 0.0Aa | 10.0 ± 11.5Ba | 22.5 ± 9.5Ca | 25.0 ± 12.9Cab | 37.5 ± 5.0Da | |
| Sf_KAY4 | 0.0 ± 0.0Aa | 10.0 ± 8.1Ba | 20.0 ± 0.0Ca | 32.5 ± 5.0Db | 40.0 ± 8.1Da | |
| Hb_N3 | 0.0 ± 0.0Aa | 7.5 ± 9.5Ba | 17.5 ± 5.0Ca | 20.0 ± 0.0Ca | 37.5 ± 5.0Da | |
| Hb_KAY10 | 0.0 ± 0.0Aa | 15.0 ± 10.0Ba | 20.0 ± 8.1Ba | 35.0 ± 5.7Cb | 50.0 ± 8.1Db | |
| Hb_AF12 | 0.0 ± 0.0Aa | 15.0 ± 5.7Ba | 22.5 ± 5.0Ba | 27.5 ± 5.0BCb | 40.0 ± 8.1Ca | |
| Temperatures | Nematodes * | Mortality Rates (%) 18 Days after Treatment (DAT) | ||||
|---|---|---|---|---|---|---|
| Control | 25 Ijs/cm2 | 50 Ijs/cm2 | 100 Ijs/cm2 | 150 Ijs/cm2 | ||
| 25 °C | Sc_BL22 | 0.0 ± 0.0A a a b | 45.0 ± 5.7Ba | 55.0 ± 5.7Ba | 75.0 ± 5.7Ca | 85.0 ± 5.7Ca |
| Sf_BL24 | 0.0 ± 0.0Aa | 35.0 ± 5.7Ba | 65.0 ± 5.7Cb | 75.0 ± 5.7Ca | 75.0 ± 5.7Ca | |
| Sf_KAY4 | 0.0 ± 0.0Aa | 42.5 ± 5.0Ca | 52.5 ± 5.0Ca | 72.5 ± 5.0Da | 72.5 ± 5.0Da | |
| Hb_N3 | 0.0 ± 0.0Aa | 52.5 ± 5.0Cab | 62.5 ± 5.0Ca | 82.5 ± 5.0Da | 82.5 ± 5.0Da | |
| Hb_KAY10 | 0.0 ± 0.0Aa | 60.0 ± 11.5Cb | 70.0 ± 11.5CDb | 80.0 ± 11.5Da | 85.0 ± 11.5Da | |
| Hb_AF12 | 0.0 ± 0.0Aa | 57.5 ± 5.0Cab | 77.5 ± 5.0Dc | 77.5 ± 5.0Da | 80.0 ± 5.0Da | |
| 30 °C | Sc_BL22 | 0.0 ± 0.0Aa | 55.0 ± 5.7Bca | 75.0 ± 5.7Ca | 85.0 ± 5.7Ca | 85.0 ± 5.7Ca |
| Sf_BL24 | 0.0 ± 0.0Aa | 55.0 ± 5.7Ca | 75.0 ± 5.7Da | 85.0 ± 5.7Da | 85.0 ± 5.7Da | |
| Sf_KAY4 | 0.0 ± 0.0Aa | 52.5 ± 5.0Ca | 72.5 ± 5.0Da | 82.5 ± 5.0Da | 82.5 ± 5.0Da | |
| Hb_N3 | 0.0 ± 0.0Aa | 55.0 ± 5.0Ca | 80.0 ± 5.0Da | 82.5 ± 5.0Da | 82.5 ± 5.0Da | |
| Hb_KAY10 | 0.0 ± 0.0Aa | 65.0 ± 11.5Cb | 80.0 ± 11.5Da | 85.0 ± 11.5Da | 85.0 ± 11.5Da | |
| Hb_AF12 | 0.0 ± 0.0Aa | 67.5 ± 5.0Bb | 80.0 ± 5.0Ca | 82.5 ± 5.0Ca | 87.5 ± 5.0Ca | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Askar, A.G.; Yüksel, E.; Bozbuğa, R.; Öcal, A.; Kütük, H.; Dinçer, D.; Canhilal, R.; Dababat, A.A.; İmren, M. Evaluation of Entomopathogenic Nematodes against Common Wireworm Species in Potato Cultivation. Pathogens 2023, 12, 288. https://doi.org/10.3390/pathogens12020288
Askar AG, Yüksel E, Bozbuğa R, Öcal A, Kütük H, Dinçer D, Canhilal R, Dababat AA, İmren M. Evaluation of Entomopathogenic Nematodes against Common Wireworm Species in Potato Cultivation. Pathogens. 2023; 12(2):288. https://doi.org/10.3390/pathogens12020288
Chicago/Turabian StyleAskar, Arife Gümüş, Ebubekir Yüksel, Refik Bozbuğa, Atilla Öcal, Halil Kütük, Dilek Dinçer, Ramazan Canhilal, Abdelfattah A. Dababat, and Mustafa İmren. 2023. "Evaluation of Entomopathogenic Nematodes against Common Wireworm Species in Potato Cultivation" Pathogens 12, no. 2: 288. https://doi.org/10.3390/pathogens12020288
APA StyleAskar, A. G., Yüksel, E., Bozbuğa, R., Öcal, A., Kütük, H., Dinçer, D., Canhilal, R., Dababat, A. A., & İmren, M. (2023). Evaluation of Entomopathogenic Nematodes against Common Wireworm Species in Potato Cultivation. Pathogens, 12(2), 288. https://doi.org/10.3390/pathogens12020288