
Evidence for Bartonella quintana in Lice Collected from the Clothes of Ethiopian Homeless Individuals


 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Molecular Diagnostics
2.3. Ethical Clearance
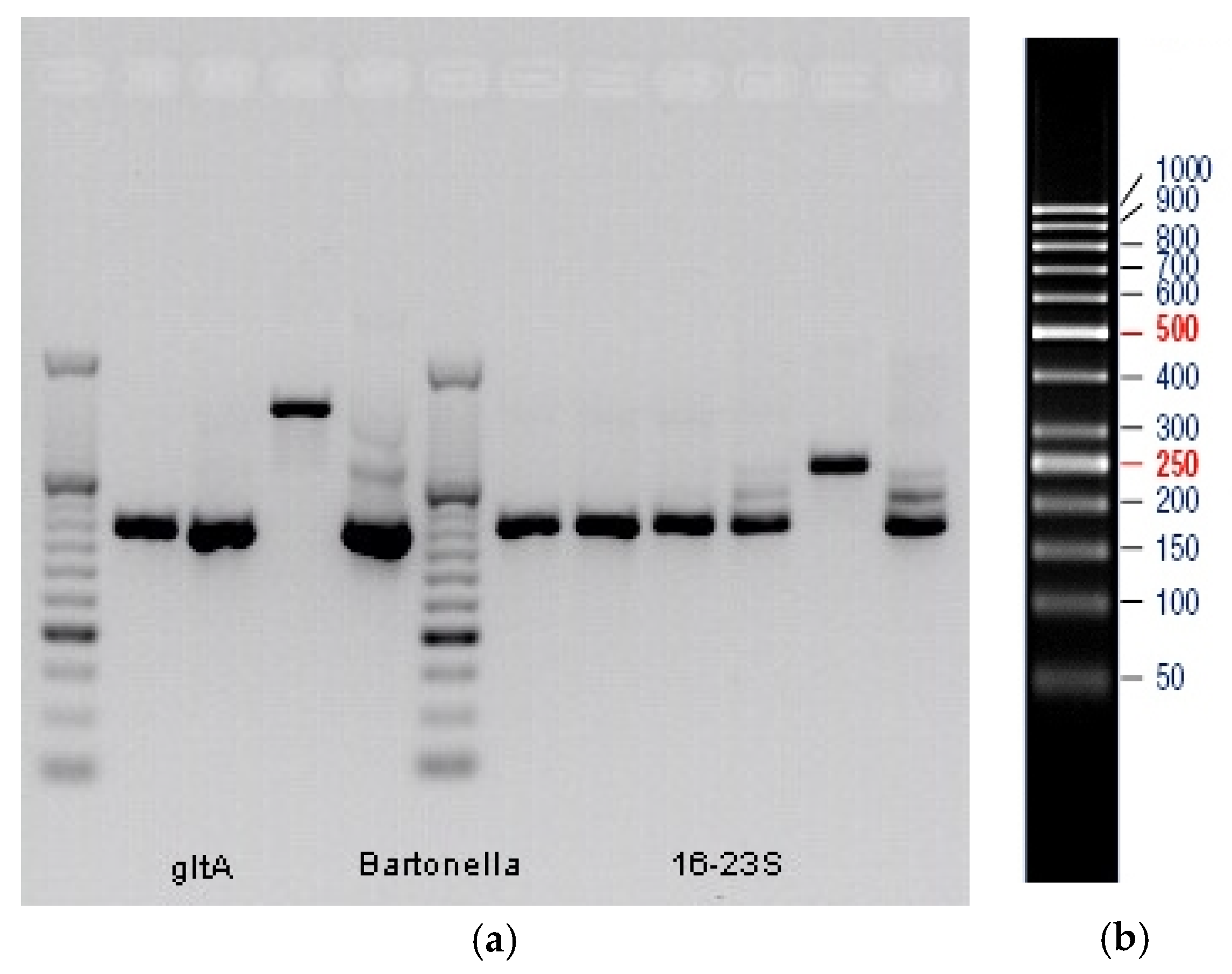
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
| 16S rRNA Gene-Specific Pan-Eubacterial PCR | |
|---|---|
| Forward primer | 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) |
| Reverse primer | 1392r (5′-ACGGGCGGTGTGTRC-3′) |
| Reaction mix | 30 µL volumes containing 15 µL of Qiagen (Hilden, Germany) Hot Start Mix, 6 µL of PCR-grade H2O, 3 µL of each primer (final concentration 10 pmol), 5 µL of DNA eluate |
| Initial denaturation | 15 min at 95 °C |
| Denaturation | 30 s at 94 °C |
| Annealing | 30 s at 54 °C |
| Amplification | 1.5 min at 72 °C |
| Cycle numbers | 30 cycles |
| Final extension | 7 min at 72 °C |
| Gel electrophoresis | Application of 6µL PCR product + 2 µL loading dye to a 1.5% agarose gel with 0.065% Gel Red nucleic acid stain |
| Bartonella Genus 16S-23S-ITS-Specific PCR (First Round) | Bartonella Genus 16S-23S-ITS-Specific PCR (Second Round) | |
|---|---|---|
| Forward primer | new-bigF (5′-GGAAGGTTTTCCGGTTTATC-3′) | new-bigF (5′-GGAAGGTTTTCCGGTTTATC-3′) |
| Reverse primer | new-bogR (5′-GTCTGAATATA(C/T)CTTCTCTTCAC-3′) | bigR (5′-TCCCAGCTGAGCTACG-3′) |
| Reaction mix | 25 µL volumes containing 12.5 µL of Qiagen (Hilden, Germany) Hot Start Mix, 9.5 µL of PCR-grade H2O, 1 µL of each primer (final concentration 10 pmol), 1 µL of DNA eluate | |
| Initial denaturation | 15 min at 95 °C | |
| Denaturation | 30 s at 95 °C | |
| Annealing | 30 s at 55 °C | |
| Amplification | 1.0 min at 72 °C | |
| Cycle numbers | 35 cycles | |
| Final extension | 5 min at 72 °C | |
| Gel electrophoresis | Application of 6 µL PCR product + 2 µL loading dye to a 1.5% agarose gel with 0.065% Gel Red nucleic acid stain | |
| Bartonella Genus gltA Gene, First Round | Bartonella Genus gltA Gene, Second Round | |
|---|---|---|
| Forward primer | 443f (5′-GCTATGTCTGCATTCTATCA-3′) | 781f (5′-GGGGACCAGCTCATGGTGG-3′) |
| Reverse primer | 1137r (5′-AATGCAAAAAGAACAGTAAACA-3′) | 1137r (5′-AATGCAAAAAGAACAGTAAACA-3′) |
| Reaction mix | 50 µL volumes containing 25 µL of Qiagen (Hilden, Germany) Hot Start Mix, 20 µL of PCR-grade H2O, 2 µL of each primer (final concentration 10 pmol), 1 µL of DNA eluate | |
| Initial denaturation | 15 min at 95 °C | |
| Denaturation | 30 s at 95 °C | |
| Annealing | 30 s at 55 °C | |
| Amplification | 1.0 min at 72 °C | |
| Cycle numbers | 35 cycles | |
| Final extension | 5 min at 72 °C | |
| Gel electrophoresis | Application of 6µL PCR product + 2 µL loading dye to a 1.5% agarose gel with 0.065% Gel Red nucleic acid stain | |
| 16S rRNA Gene PCR Targeting Relapsing-Fever-Associated Borrelia spp. | |
|---|---|
| Forward primer | 5′-GGCTTAGAACTAACGCTGGCAGTGC-3′ |
| Reverse primer | 5′-CCCTTTACGCCCAATAATCCCGA-3′ |
| Reaction mix | 30 µL volumes containing 15 µL of Qiagen (Hilden, Germany) Hot Start Mix, 6 µL of PCR-grade H2O, 3 µL of each primer (final concentration 10 pmol), 5 µL of DNA eluate |
| Initial denaturation | 15 min at 95 °C |
| Denaturation | 15 s at 94 °C |
| Annealing | 30 s at 54 °C |
| Amplification | 1 min at 72 °C |
| Cycle numbers | 40 cycles |
| Final extension | 7 min at 72 °C |
| Gel electrophoresis | Application of 6µL PCR product + 2 µL loading dye to a 1.5% agarose gel with 0.065% Gel Red nucleic acid stain |
References
- Barbieri, R.; Drancourt, M.; Raoult, D. The role of louse-transmitted diseases in historical plague pandemics. Lancet Infect. Dis. 2021, 21, e17–e25. [Google Scholar] [CrossRef]
- Kahlig, P.; Neumayr, A.; Paris, D.H. Louse-borne relapsing fever-A systematic review and analysis of the literature: Part 2-Mortality, Jarisch-Herxheimer reaction, impact on pregnancy. PLoS Negl. Trop. Dis. 2021, 15, e0008656. [Google Scholar] [CrossRef]
- Brouqui, P.; Raoult, D. Arthropod-borne diseases in homeless. Ann. N. Y. Acad. Sci. 2006, 1078, 223–235. [Google Scholar] [CrossRef]
- Billeter, S.A.; Levy, M.G.; Chomel, B.B.; Breitschwerdt, E.B. Vector transmission of Bartonella species with emphasis on the potential for tick transmission. Med. Vet. Entomol. 2008, 22, 1–15. [Google Scholar] [CrossRef]
- Azad, A.F.; Beard, C.B. Rickettsial pathogens and their arthropod vectors. Emerg. Infect. Dis. 1998, 4, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.J.; Ammerman, N.C.; Beier-Sexton, M.; Sobral, B.S.; Azad, A.F. Louse- and flea-borne rickettsioses: Biological and genomic analyses. Vet. Res. 2009, 40, 12. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.; Leo, N.; Prociv, P.; Barker, S.C. Potential role of head lice, Pediculus humanus capitis, as vectors of Rickettsia prowazekii. Parasitol. Res. 2003, 90, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.L.; Chang, C.C.; Chuang, S.T.; Chomel, B.B. Bartonella species and their ectoparasites: Selective host adaptation or strain selection between the vector and the mammalian host? Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Badiaga, S.; Brouqui, P. Human louse-transmitted infectious diseases. Clin. Microbiol. Infect. 2012, 18, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Brouqui, P. Arthropod-borne diseases associated with political and social disorder. Annu. Rev. Entomol. 2011, 56, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Amanzougaghene, N.; Fenollar, F.; Raoult, D.; Mediannikov, O. Where Are We With Human Lice? A Review of the Current State of Knowledge. Front. Cell. Infect. Microbiol. 2020, 9, 474. [Google Scholar] [CrossRef]
- Boutellis, A.; Abi-Rached, L.; Raoult, D. The origin and distribution of human lice in the world. Infect. Genet. Evol. 2014, 23, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Jones, D. The neglected saliva: Medically important toxins in the saliva of human lice. Parasitology 1998, 116, S73–S81. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Roux, V. The body louse as a vector of reemerging human diseases. Clin. Infect. Dis. 1999, 29, 888–911. [Google Scholar] [CrossRef]
- Estrada, B. Ectoparasitic infestations in homeless children. Semin. Pediatr. Infect. Dis. 2003, 14, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.; Hovius, J.W.; Bergström, S. Pathogenesis of Relapsing Fever. Curr. Issues Mol. Biol. 2021, 42, 519–550. [Google Scholar]
- Cutler, S.J. Relapsing Fever Borreliae: A Global Review. Clin. Lab. Med. 2015, 35, 847–865. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.J.; Abdissa, A.; Trape, J.F. New concepts for the old challenge of African relapsing fever borreliosis. Clin. Microbiol. Infect. 2009, 15, 400–406. [Google Scholar] [CrossRef]
- Antinori, S.; Mediannikov, O.; Corbellino, M.; Raoult, D. Louse-borne relapsing fever among East African refugees in Europe. Travel Med. Infect. Dis. 2016, 14, 110–114. [Google Scholar] [CrossRef]
- Eiset, A.H.; Wejse, C. Review of infectious diseases in refugees and asylum seekers-current status and going forward. Public Health Rev. 2017, 38, 22. [Google Scholar] [CrossRef]
- Ahmed, M.A.; Abdel Wahab, S.M.; Abdel Malik, M.O.; Abdel Gadir, A.M.; Salih, S.Y.; Omer, A.; Al Hassan, A.M. Louse-borne relapsing fever in the Sudan. A historical review and a clinico-pathological study. Trop. Geogr. Med. 1980, 32, 106–211. [Google Scholar] [PubMed]
- Warrell, D.A. Louse-borne relapsing fever (Borrelia recurrentis infection). Epidemiol. Infect. 2019, 147, e106. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.J. Relapsing fever—A forgotten disease revealed. J. Appl. Microbiol. 2010, 108, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- El-Bahnsawy, M.M.; Labib, N.A.; Abdel-Fattah, M.A.; Ibrahim, A.M.; Morsy, T.A. Louse and tick borne relapsing fevers. J. Egypt. Soc. Parasitol. 2012, 42, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, T.; Feldt, T.; Bosselmann, M.; Tufa, T.B.; Lemma, G.; Holtfreter, M.; Häussinger, D. Outbreak of Louse-Borne Relapsing Fever among Urban Dwellers in Arsi Zone, Central Ethiopia, from July to November 2016. Am. J. Trop. Med. Hyg. 2018, 98, 1599–1602. [Google Scholar] [CrossRef] [PubMed]
- Maurin, M.; Raoult, D. Bartonella (Rochalimaea) quintana infections. Clin. Microbiol. Rev. 1996, 9, 273–292. [Google Scholar] [CrossRef] [PubMed]
- Leibler, J.H.; Zakhour, C.M.; Gadhoke, P.; Gaeta, J.M. Zoonotic and Vector-Borne Infections Among Urban Homeless and Marginalized People in the United States and Europe, 1990-2014. Vector Borne Zoonotic Dis. 2016, 16, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Ohl, M.E.; Spach, D.H. Bartonella quintana and urban trench fever. Clin. Infect. Dis. 2000, 31, 131–135. [Google Scholar] [CrossRef]
- Ruiz, J. Bartonella quintana, past, present, and future of the scourge of World War I. APMIS. 2018, 126, 831–837. [Google Scholar] [CrossRef]
- Anstead, G.M. The centenary of the discovery of trench fever, an emerging infectious disease of World War 1. Lancet Infect. Dis. 2016, 16, e164–e172. [Google Scholar] [CrossRef]
- Lam, J.C.; Fonseca, K.; Pabbaraju, K.; Meatherall, B.L. Case Report: Bartonella quintana Endocarditis Outside of the Europe-African Gradient: Comprehensive Review of Cases within North America. Am. J. Trop. Med. Hyg. 2019, 100, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Spach, D.H.; Kanter, A.S.; Dougherty, M.J.; Larson, A.M.; Coyle, M.B.; Brenner, D.J.; Swaminathan, B.; Matar, G.M.; Welch, D.F.; Root, R.K.; et al. Bartonella (Rochalimaea) quintana bacteremia in inner-city patients with chronic alcoholism. N. Engl. J. Med. 1995, 332, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Maurin, M.; Birtles, R.; Raoult, D. Current knowledge of Bartonella species. Eur. J. Clin. Microbiol. Infect. Dis. 1997, 16, 487–506. [Google Scholar] [CrossRef] [PubMed]
- Veracx, A.; Raoult, D. Biology and genetics of human head and body lice. Trends Parasitol. 2012, 28, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Bechah, Y.; Capo, C.; Mege, J.L.; Raoult, D. Epidemic typhus. Lancet Infect Dis. 2008, 8, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Laroche, M.; Raoult, D.; Parola, P. Insects and the Transmission of Bacterial Agents. Microbiol. Spectr. 2018, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Kazár, J.; Brezina, R. Control of rickettsial diseases. Eur. J. Epidemiol. 1991, 7, 282–286. [Google Scholar]
- Brinkmann, A.; Nitsche, A.; Kohl, C. Viral Metagenomics on Blood-Feeding Arthropods as a Tool for Human Disease Surveillance. Int. J. Mol. Sci. 2016, 17, 1743. [Google Scholar] [CrossRef]
- Margos, G.; Hepner, S.; Mang, C.; Marosevic, D.; Reynolds, S.E.; Krebs, S.; Sing, A.; Derdakova, M.; Reiter, M.A.; Fingerle, V. Lost in plasmids: Next generation sequencing and the complex genome of the tick-borne pathogen Borrelia burgdorferi. BMC Genom. 2017, 18, 422. [Google Scholar] [CrossRef]
- Ruegger, P.M.; Della Vedova, G.; Jiang, T.; Borneman, J. Improving probe set selection for microbial community analysis by leveraging taxonomic information of training sequences. BMC Bioinform. 2011, 12, 394. [Google Scholar] [CrossRef]
- Telfer, S.; Bown, K.J.; Sekules, R.; Begon, M.; Hayden, T.; Birtles, R. Disruption of a host-parasite system following the introduction of an exotic host species. Parasitology 2005, 130 Pt 6, 661–668. [Google Scholar] [CrossRef]
- Logan, J.M.J.; Hall, J.L.; Chalker, V.J.; O’Connell, B.; Birtles, R.J. Bartonella clarridgeiae infection in a patient with aortic root abscess and endocarditis. Access Microbiol. 2019, 1, e000064. [Google Scholar] [CrossRef]
- Regier, Y.; Komma, K.; Weigel, M.; Kraiczy, P.; Laisi, A.; Pulliainen, A.T.; Hain, T.; Kempf, V.A.J. Combination of microbiome analysis and serodiagnostics to assess the risk of pathogen transmission by ticks to humans and animals in central Germany. Parasit Vectors 2019, 12, 11. [Google Scholar] [CrossRef]
- DIN EN ISO/IEC 17025; Allgemeine Anforderungen an die Kompetenz von Prüf- und Kalibrierlaboratorien. Deutsche Industrienorm (DIN): Berlin, Germany, 2018.
- Pérez-Tanoira, R.; Ramos-Rincón, J.M.; Martín-Martín, I.; Prieto-Pérez, L.; Tefasmariam, A.; Tiziano, G.; Anda, P.; González-Martín-Niño, R.M.; Rodríguez-Vargas, M.; Górgolas, M.; et al. Molecular Survey of Rickettsia spp., Anaplasma spp., Ehrlichia spp., Bartonella spp., and Borrelia spp. in Fleas and Lice in Ethiopia. Vector Borne Zoonotic Dis. 2020, 20, 10–14. [Google Scholar] [CrossRef]
- Cutler, S.; Abdissa, A.; Adamu, H.; Tolosa, T.; Gashaw, A. Bartonella quintana in Ethiopian lice. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 17–21. [Google Scholar] [CrossRef]
- Angelakis, E.; Diatta, G.; Abdissa, A.; Trape, J.F.; Mediannikov, O.; Richet, H.; Raoult, D. Altitude-dependent Bartonella quintana genotype C in head lice, Ethiopia. Emerg. Infect. Dis. 2011, 17, 2357–2359. [Google Scholar] [CrossRef]
- Tasher, D.; Raucher-Sternfeld, A.; Tamir, A.; Giladi, M.; Somekh, E. Bartonella quintana, an Unrecognized Cause of Infective Endocarditis in Children in Ethiopia. Emerg. Infect. Dis. 2017, 23, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tufa, T.B.; Margos, G.; Fingerle, V.; Hartberger, C.; Poppert, S.; Birtles, R.J.; Kraiczy, P.; Kempf, V.A.J.; Frickmann, H.; Feldt, T. Evidence for Bartonella quintana in Lice Collected from the Clothes of Ethiopian Homeless Individuals. Pathogens 2023, 12, 1299. https://doi.org/10.3390/pathogens12111299
Tufa TB, Margos G, Fingerle V, Hartberger C, Poppert S, Birtles RJ, Kraiczy P, Kempf VAJ, Frickmann H, Feldt T. Evidence for Bartonella quintana in Lice Collected from the Clothes of Ethiopian Homeless Individuals. Pathogens. 2023; 12(11):1299. https://doi.org/10.3390/pathogens12111299
Chicago/Turabian StyleTufa, Tafese Beyene, Gabriele Margos, Volker Fingerle, Christine Hartberger, Sven Poppert, Richard J. Birtles, Peter Kraiczy, Volkhard A. J. Kempf, Hagen Frickmann, and Torsten Feldt. 2023. "Evidence for Bartonella quintana in Lice Collected from the Clothes of Ethiopian Homeless Individuals" Pathogens 12, no. 11: 1299. https://doi.org/10.3390/pathogens12111299
APA StyleTufa, T. B., Margos, G., Fingerle, V., Hartberger, C., Poppert, S., Birtles, R. J., Kraiczy, P., Kempf, V. A. J., Frickmann, H., & Feldt, T. (2023). Evidence for Bartonella quintana in Lice Collected from the Clothes of Ethiopian Homeless Individuals. Pathogens, 12(11), 1299. https://doi.org/10.3390/pathogens12111299