
β-Defensin-1 Regulates Influenza Virus Infection in Human Bronchial Epithelial Cells through the STAT3 Signaling Pathway



Abstract
1. Introduction
2. Materials and Methods
2.1. Propagation of Cells
2.2. Viral Infections
2.3. hBD1 Overexpression Studies
2.4. Transfection of miRNAs and siRNAs
2.5. Defensin β-1 Knock-Down Using siRNAs
2.6. Plaque Assay for IAV Detection
2.7. Gene Expression and Transcription Factor PCR Array
2.8. Microscopy
2.9. Western Immunoblot Analysis
2.10. Argonaute Immunoprecipitation
2.11. Ingenuity Pathway Analysis
2.12. Statistical Analysis
3. Results
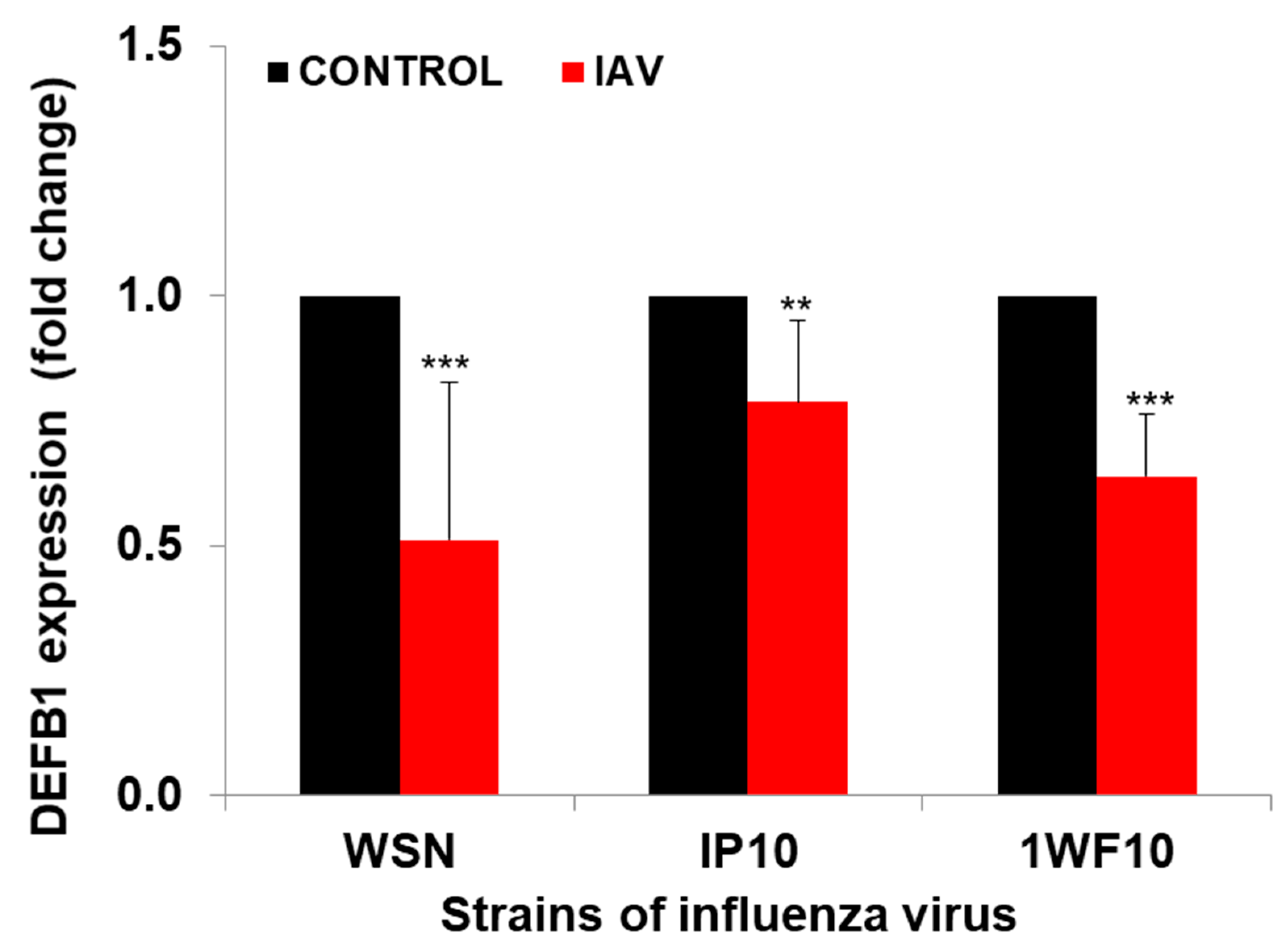
3.1. Role of Defensin-β1 in IAV Replication
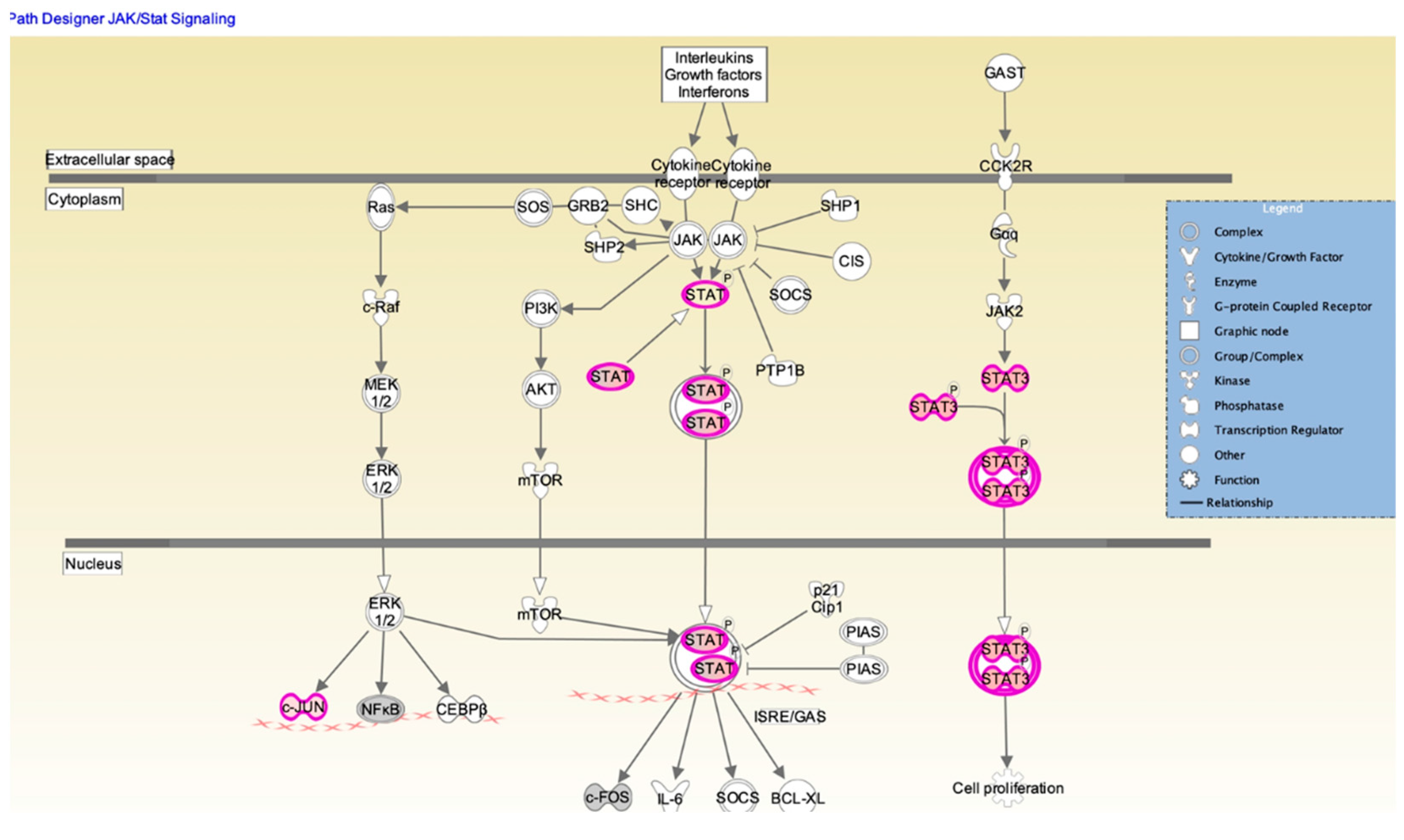
3.2. Defensin-β1 Function through STAT3 Pathway
3.3. Role of miRNAs Regulating the Expression of Defensin-β1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ARNT | Aryl hydrocarbon receptor nuclear translocator |
| ATF1 | Activating transcription factor 1 |
| ATF2 | Activating transcription factor 2 |
| ATF3 | Activating transcription factor 3 |
| ATF4 | Activating transcription factor 4 |
| CEBPA | CCAAT/enhancer binding protein (C/EBP), alpha |
| CEBPB | CCAAT/enhancer binding protein (C/EBP), beta |
| CEBPG | CCAAT/enhancer binding protein (C/EBP), gamma |
| CREB1 | CAMP responsive element binding protein 1 |
| CREBBP | CREB binding protein |
| CTNNB1 | Catenin (cadherin-associated protein), beta 1 |
| DR1 | Downregulator of transcription 1, TBP-binding |
| E2F1 | E2F transcription factor 1 |
| E2F6 | E2F transcription factor 6 |
| EGR1 | Early growth response 1 |
| ELK1 | ELK1, member of ETS oncogene family |
| ESR1 | Estrogen receptor 1 |
| ETS1 | V-Ets erythroblastosis virus E26 oncogene homolog 1 |
| ETS2 | V-Ets erythroblastosis virus E26 oncogene homolog 2 |
| FOS | Fos Proto-Oncogene, AP-1 Transcription Factor Subunit |
| FOXA2 | Forkhead box A2 |
| FOXO1 | Forkhead box O1 |
| GATA1 | GATA binding protein 1 (globin transcription factor 1) |
| GATA2 | GATA binding protein 2 |
| GATA3 | GATA binding protein 3 |
| GTF2B | General transcription factor IIB |
| GTF2F1 | General transcription factor IIF, polypeptide 1, 74 kDa |
| HAND1 | Heart and neural crest derivatives expressed 1 |
| HAND2 | Heart and neural crest derivatives expressed 2 |
| HDAC1 | Histone deacetylase 1 |
| HIF1A | Hypoxia inducible factor 1, alpha subunit |
| HNF4A | Hepatocyte nuclear factor 4, alpha |
| HOXA5 | Homeobox A5 |
| HSF1 | Heat shock transcription factor 1 |
| ID1 | Inhibitor of DNA binding 1 |
| IRF1 | Interferon regulatory factor 1 |
| JUN | Jun proto-oncogene |
| JUNB | Jun B proto-oncogene |
| MAX | MYC associated factor X |
| MEF2A | Myocyte enhancer factor 2A |
| MEF2B | Myocyte enhancer factor 2B |
| MEF2C | Myocyte enhancer factor 2C |
| NFAT5 | Nuclear factor of activated T-cells 5, tonicity-responsive |
| NFATC1 | Nuclear factor of activated T-cells, calcineurin-dependent 1 |
| NFATC2 | Nuclear factor of activated T-cells, calcineurin-dependent 2 |
| NFATC3 | Nuclear factor of activated T-cells, calcineurin-dependent 3 |
| NFATC4 | Nuclear factor of activated T-cells, calcineurin-dependent 4 |
| NFKB1 | Nuclear factor of kappa B 1 |
| NFYB | Nuclear transcription factor Y, beta |
| NR3C1 | Nuclear receptor subfamily 3, group C, member 1 |
| PPARA | Peroxisome proliferator-activated receptor alpha |
| PPARG | Peroxisome proliferator-activated receptor gamma |
| RB1 | Retinoblastoma 1 |
| REL | REL V-rel reticuloendotheliosis viral oncogene homolog |
| RELA | V-rel reticuloendotheliosis viral oncogene homolog A |
| RELB | RELB V-rel reticuloendotheliosis viral oncogene homolog B |
| SMAD1 | SMAD family member 1 |
| SMAD4 | SMAD family member 4 |
| SMAD5 | SMAD family member 5 |
| SMAD9 | SMAD family member 9 |
| SP1 | Sp1 transcription factor |
| SP3 | Sp3 transcription factor |
| STAT1 | Signal transducer and activator of transcription 1, |
| STAT2 | Signal transducer and activator of transcription 2, |
| STAT3 | Signal transducer and activator of transcription 3 |
| STAT4 | Signal transducer and activator of transcription 4 |
| STAT5A | Signal transducer and activator of transcription 5A |
| STAT5B | Signal transducer and activator of transcription 5B |
| STAT6 | Signal transducer and activator of transcription 6 |
| TBP | TATA box binding protein |
| TCF7L2 | Transcription Factor 7 Like 2 |
| TFAP2A | Transcription factor AP-2 alpha |
| TGIF1 | TGFB-induced factor homeobox 1 |
| TP53 | Tumor protein p53 |
References
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Becknell, B.; Spencer, J.D.; Carpenter, A.R.; Chen, X.; Singh, A.; Ploeger, S.; Kline, J.; Ellsworth, P.; Li, B.; Proksch, E.; et al. Expression and antimicrobial function of beta-defensin 1 in the lower urinary tract. PLoS ONE 2013, 8, e77714. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, L.; Wu, M. Defensins in innate immunity. Cell Tissue Res. 2011, 343, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Selsted, M.E.; Ouellette, A.J. Mammalian defensins in the antimicrobial immune response. Nat. Immunol. 2005, 6, 551–557. [Google Scholar] [CrossRef]
- Schutte, B.C.; Mitros, J.P.; Bartlett, J.A.; Walters, J.D.; Jia, H.P.; Welsh, M.J.; Casavant, T.L.; McCray, P.B., Jr. Discovery of five conserved beta -defensin gene clusters using a computational search strategy. Proc. Natl. Acad. Sci. USA 2002, 99, 2129–2133. [Google Scholar] [CrossRef] [PubMed]
- Garreis, F.; Schlorf, T.; Worlitzsch, D.; Steven, P.; Bräuer, L.; Jäger, K.; Paulsen, F.P. Roles of human β-defensins in innate immune defense at the ocular surface: Arming and alarming corneal and conjunctival epithelial cells. Histochem. Cell Biol. 2010, 134, 59–73. [Google Scholar] [CrossRef]
- Doss, M.; White, M.R.; Tecle, T.; Hartshorn, K.L. Human defensins and LL-37 in mucosal immunity. J. Leukoc. Biol. 2010, 87, 79–92. [Google Scholar] [CrossRef]
- Ryan, L.K.; Diamond, G. Modulation of Human β-Defensin-1 Production by Viruses. Viruses 2017, 9, 153. [Google Scholar] [CrossRef]
- Wilson, S.S.; Wiens, M.E.; Holly, M.K.; Smith, J.G. Defensins at the Mucosal Surface: Latest Insights into Defensin-Virus Interactions. J. Virol. 2016, 90, 5216–5218. [Google Scholar] [CrossRef]
- Diamond, G.; Beckloff, N.; Ryan, L.K. Host defense peptides in the oral cavity and the lung: Similarities and differences. J. Dent. Res. 2008, 87, 915–927. [Google Scholar] [CrossRef]
- Findlay, E.G.; Currie, S.M.; Davidson, D.J. Cationic host defence peptides: Potential as antiviral therapeutics. BioDrugs 2013, 27, 479–493. [Google Scholar] [CrossRef]
- Ding, J.; Chou, Y.Y.; Chang, T.L. Defensins in viral infections. J. Innate Immun. 2009, 1, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, J.K.; Morens, D.M. The pathology of influenza virus infections. Annu. Rev. Pathol. 2008, 3, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Gentile, D.; Doyle, W.; Whiteside, T.; Fireman, P.; Hayden, F.G.; Skoner, D. Increased interleukin-6 levels in nasal lavage samples following experimental influenza A virus infection. Clin. Diagn. Lab. Immunol. 1998, 5, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Konig, R.; Stertz, S.; Zhou, Y.; Inoue, A.; Hoffmann, H.H.; Bhattacharyya, S.; Alamares, J.G.; Tscherne, D.M.; Ortigoza, M.B.; Liang, Y.; et al. Human host factors required for influenza virus replication. Nature 2010, 463, 813–817. [Google Scholar] [CrossRef]
- Karlas, A.; Machuy, N.; Shin, Y.; Pleissner, K.P.; Artarini, A.; Heuer, D.; Becker, D.; Khalil, H.; Ogilvie, L.A.; Hess, S.; et al. Genome-wide RNAi screen identifies human host factors crucial for influenza virus replication. Nature 2010, 463, 818–822. [Google Scholar] [CrossRef]
- Gong, T.; Jiang, Y.; Wang, Y.; Yang, D.; Li, W.; Zhang, Q.; Feng, W.; Wang, B.; Jiang, Z.; Li, M. Recombinant mouse beta-defensin 2 inhibits infection by influenza A virus by blocking its entry. Arch. Virol. 2010, 155, 491–498. [Google Scholar] [CrossRef]
- Othumpangat, S.; Lindsley, W.G.; Beezhold, D.H.; Kashon, M.L.; Burrell, C.N.; Mubareka, S.; Noti, J.D. Differential Expression of Serum Exosome microRNAs and Cytokines in Influenza A and B Patients Collected in the 2016 and 2017 Influenza Seasons. Pathogens 2021, 10, 149. [Google Scholar] [CrossRef]
- Othumpangat, S.; Noti, J.D.; Blachere, F.M.; Beezhold, D.H. Expression of non-structural-1A binding protein in lung epithelial cells is modulated by miRNA-548an on exposure to influenza A virus. Virology 2013, 447, 84–94. [Google Scholar] [CrossRef]
- Othumpangat, S.; Noti, J.D.; Beezhold, D.H. Lung epithelial cells resist influenza A infection by inducing the expression of cytochrome c oxidase VIc which is modulated by miRNA 4276. Virology 2014, 468–470, 256–264. [Google Scholar] [CrossRef]
- Othumpangat, S.; Bryan, N.B.; Beezhold, D.H.; Noti, J.D. Upregulation of miRNA-4776 in Influenza Virus Infected Bronchial Epithelial Cells Is Associated with Downregulation of NFKBIB and Increased Viral Survival. Viruses 2017, 9, 94. [Google Scholar] [CrossRef]
- Kuhn, D.E.; Martin, M.M.; Feldman, D.S.; Terry, A.V., Jr.; Nuovo, G.J.; Elton, T.S. Experimental validation of miRNA targets. Methods 2008, 44, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Dauletbaev, N.; Gropp, R.; Frye, M.; Loitsch, S.; Wagner, T.O.; Bargon, J. Expression of human beta defensin (HBD-1 and HBD-2) mRNA in nasal epithelia of adult cystic fibrosis patients, healthy individuals, and individuals with acute cold. Respiration 2002, 69, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Jiang, L.; Lai, W.; Qin, Y.; Zhang, T.; Wang, S.; Ye, X. MicroRNA-33a disturbs influenza A virus replication by targeting ARCN1 and inhibiting viral ribonucleoprotein activity. J. Gen. Virol. 2016, 97, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Peschel, A.; Sahl, H.-G. The co-evolution of host cationic antimicrobial peptides and microbial resistance. Nat. Rev. Microbiol. 2006, 4, 529–536. [Google Scholar] [CrossRef]
- Krishnakumari, V.; Rangaraj, N.; Nagaraj, R. Antifungal Activities of Human Beta-Defensins HBD-1 to HBD-3 and Their C-Terminal Analogs Phd1 to Phd3. Antimicrob. Agents Chemother. 2009, 53, 256–260. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, I.; Lehrer, R.I. Widespread expression of beta-defensin hBD-1 in human secretory glands and epithelial cells. FEBS Lett. 1996, 396, 319–322. [Google Scholar] [CrossRef]
- Ryan, L.K.; Dai, J.; Yin, Z.; Megjugorac, N.; Uhlhorn, V.; Yim, S.; Schwartz, K.D.; Abrahams, J.M.; Diamond, G.; Fitzgerald-Bocarsly, P. Modulation of human beta-defensin-1 (hBD-1) in plasmacytoid dendritic cells (PDC), monocytes, and epithelial cells by influenza virus, Herpes simplex virus, and Sendai virus and its possible role in innate immunity. J. Leukoc. Biol. 2011, 90, 343–356. [Google Scholar] [CrossRef]
- Andresen, E.; Gunther, G.; Bullwinkel, J.; Lange, C.; Heine, H. Increased expression of beta-defensin 1 (DEFB1) in chronic obstructive pulmonary disease. PLoS ONE 2011, 6, e21898. [Google Scholar] [CrossRef]
- Grubor, B.; Gallup, J.M.; Meyerholz, D.K.; Crouch, E.C.; Evans, R.B.; Brogden, K.A.; Lehmkuhl, H.D.; Ackermann, M.R. Enhanced surfactant protein and defensin mRNA levels and reduced viral replication during parainfluenza virus type 3 pneumonia in neonatal lambs. Clin. Diagn. Lab. Immunol. 2004, 11, 599–607. [Google Scholar] [CrossRef]
- Chong, K.T.; Thangavel, R.R.; Tang, X. Enhanced expression of murine beta-defensins (MBD-1, -2,- 3, and -4) in upper and lower airway mucosa of influenza virus infected mice. Virology 2008, 380, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Khongnomnan, K.; Makkoch, J.; Poomipak, W.; Poovorawan, Y.; Payungporn, S. Human miR-3145 inhibits influenza A viruses replication by targeting and silencing viral PB1 gene. Exp. Biol. Med. 2015, 240, 1630–1639. [Google Scholar] [CrossRef] [PubMed]
- Kaeuferle, T.; Bartel, S.; Dehmel, S.; Krauss-Etschmann, S. MicroRNA methodology: Advances in miRNA technologies. Methods Mol. Biol. 2014, 1169, 121–130. [Google Scholar] [PubMed]
- Izaurralde, E. Breakers and blockers-miRNAs at work. Science 2015, 349, 380–382. [Google Scholar] [CrossRef]
- Jonas, S.; Izaurralde, E. Towards a molecular understanding of microRNA-mediated gene silencing. Nat. Rev. Genet. 2015, 16, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, J.; Lee, S.; Lee, B.; Kim-Ha, J. Phylogenetic comparison of oskar mRNA localization signals. Biochem. Biophys. Res. Commun. 2014, 444, 98–103. [Google Scholar] [CrossRef] [PubMed]
- De Tullio, G.; De Fazio, V.; Sgherza, N.; Minoia, C.; Serrati, S.; Merchionne, F.; Loseto, G.; Iacobazzi, A.; Rana, A.; Petrillo, P.; et al. Challenges and opportunities of microRNAs in lymphomas. Molecules 2014, 19, 14723–14781. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.A.; Zhong, Z.; Wen, Z.; Darnell, J.E., Jr. STAT signaling is active during early mammalian development. Dev. Dyn. 1997, 208, 190–198. [Google Scholar] [CrossRef]
- Kuchipudi, S.V. The Complex Role of STAT3 in Viral Infections. J. Immunol. Res. 2015, 2015, 272359. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fold Change | |||||
|---|---|---|---|---|---|
| Gene Symbol | Plasmid Control | DEFB1 Control | DEFB1 siRNA | Control+ A(H1N1) | DEFB1+ A(H1N1) |
| HPRT1 | 1.00 | 1.26 | 0.99 | 1.00 | 1.03 |
| HMBS | 1.00 | 1.30 | 1.40 | 1.00 | 0.48 |
| TBP | 1.00 | 1.18 | 1.03 | 1.00 | 0.42 |
| PGK1 | 1.00 | 1.07 | 1.33 | 1.00 | 0.71 |
| UBC | 1.00 | 1.71 | 0.89 | 1.00 | 0.41 |
| PPIA | 1.00 | 1.28 | 1.51 | 1.00 | 0.78 |
| ARNT | 1.00 | 1.28 | 1.24 | 1.00 | 0.48 |
| ATF1 | 1.00 | 4.26 | 1.52 | 1.00 | 1.76 |
| ATF2 | 1.00 | 1.28 | 0.96 | 1.00 | 0.60 |
| ATF3 | 1.00 | 1.03 | 0.42 | 1.00 | 3.16 |
| ATF4 | 1.00 | 1.32 | 1.14 | 1.00 | 0.33 |
| CEBPA | 1.00 | 1.02 | 0.42 | 1.00 | 0.31 |
| CEBPB | 1.00 | 1.16 | 0.90 | 1.00 | 0.67 |
| CEBPG | 1.00 | 0.60 | 0.92 | 1.00 | 0.14 |
| CREB1 | 1.00 | 1.20 | 0.85 | 1.00 | 0.72 |
| CREBBP | 1.00 | 0.97 | 0.57 | 1.00 | 0.42 |
| CTNNB1 | 1.00 | 1.48 | 1.07 | 1.00 | 0.84 |
| DR1 | 1.00 | 0.92 | 1.21 | 1.00 | 0.33 |
| E2F1 | 1.00 | 0.93 | 1.70 | 1.00 | 0.19 |
| E2F6 | 1.00 | 1.04 | 0.74 | 1.00 | 0.26 |
| EGR1 | 1.00 | 1.13 | 0.81 | 1.00 | 4.95 |
| ELK1 | 1.00 | 1.12 | 0.69 | 1.00 | 0.34 |
| ESR1 | 1.00 | 1.31 | 0.36 | 1.00 | 0.17 |
| ETS1 | 1.00 | 1.66 | 1.31 | 1.00 | 0.29 |
| ETS2 | 1.00 | 0.76 | 1.01 | 1.00 | 1.65 |
| FOXA2 | 1.00 | 0.65 | 0.97 | 1.00 | 0.28 |
| FOXO1 | 1.00 | 3.06 | 0.97 | 1.00 | 0.64 |
| GATA3 | 1.00 | 1.19 | 1.09 | 1.00 | 3.01 |
| GTF2B | 1.00 | 0.22 | 1.59 | 1.00 | 0.11 |
| GTF2F1 | 1.00 | 1.13 | 0.79 | 1.00 | 0.46 |
| HDAC1 | 1.00 | 1.08 | 1.30 | 1.00 | 0.45 |
| HIF1A | 1.00 | 0.95 | 0.45 | 1.00 | 0.09 |
| HSF1 | 1.00 | 1.86 | 1.01 | 1.00 | 0.58 |
| ID1 | 1.00 | 0.57 | 1.02 | 1.00 | 0.38 |
| IRF1 | 1.00 | 2.14 | 0.92 | 1.00 | 2.42 |
| JUN | 1.00 | 0.77 | 0.93 | 1.00 | 1.04 |
| JUNB | 1.00 | 1.48 | 1.61 | 1.00 | 1.44 |
| MAX | 1.00 | 0.97 | 1.00 | 1.00 | 0.64 |
| MEF2A | 1.00 | 1.48 | 0.77 | 1.00 | 0.45 |
| MEF2B | 1.00 | 1.76 | 1.20 | 1.00 | 0.61 |
| MEF2C | 1.00 | 0.07 | 0.43 | 1.00 | 4.19 |
| NFAT5 | 1.00 | 1.00 | 1.26 | 1.00 | 0.14 |
| NFATC1 | 1.00 | 0.77 | 0.78 | 1.00 | 0.20 |
| NFATC2 | 1.00 | 1.12 | 0.39 | 1.00 | 0.39 |
| NFATC3 | 1.00 | 1.29 | 1.13 | 1.00 | 0.44 |
| NFATC4 | 1.00 | 1.54 | 1.17 | 1.00 | 0.34 |
| NFKB1 | 1.00 | 1.88 | 0.77 | 1.00 | 1.10 |
| NFYB | 1.00 | 0.55 | 0.87 | 1.00 | 0.16 |
| NR3C1 | 1.00 | 0.76 | 0.96 | 1.00 | 0.49 |
| PPARA | 1.00 | 1.68 | 0.78 | 1.00 | 0.30 |
| PPARG | 1.00 | 0.27 | 0.28 | 1.00 | 0.10 |
| RB1 | 1.00 | 1.13 | 1.01 | 1.00 | 0.33 |
| REL | 1.00 | 1.12 | 1.19 | 1.00 | 5.99 |
| RELA | 1.00 | 1.18 | 1.00 | 1.00 | 0.61 |
| RELB | 1.00 | 2.29 | 1.36 | 1.00 | 0.93 |
| SMAD1 | 1.00 | 0.98 | 0.66 | 1.00 | 0.31 |
| SMAD4 | 1.00 | 0.97 | 0.92 | 1.00 | 0.32 |
| SMAD5 | 1.00 | 1.22 | 1.50 | 1.00 | 0.46 |
| SMAD9 | 1.00 | 3.36 | 1.81 | 1.00 | 1.93 |
| SP1 | 1.00 | 0.88 | 1.12 | 1.00 | 0.38 |
| SP3 | 1.00 | 0.68 | 0.48 | 1.00 | 0.26 |
| STAT1 | 1.00 | 0.61 | 0.34 | 1.00 | 1.25 |
| STAT2 | 1.00 | 0.69 | 0.74 | 1.00 | 2.18 |
| STAT3 | 1.00 | 2.03 | 0.75 | 1.00 | 2.16 |
| STAT4 | 1.00 | 1.25 | 1.58 | 1.00 | 0.80 |
| STAT5A | 1.00 | 1.01 | 0.78 | 1.00 | 0.65 |
| STAT5B | 1.00 | 1.13 | 1.01 | 1.00 | 0.31 |
| STAT6 | 1.00 | 1.68 | 1.27 | 1.00 | 1.07 |
| TBP | 1.00 | 1.37 | 0.82 | 1.00 | 0.42 |
| TCF7L2 | 1.00 | 1.43 | 1.00 | 1.00 | 0.40 |
| TFAP2A | 1.00 | 1.56 | 0.46 | 1.00 | 0.64 |
| TGIF1 | 1.00 | 1.92 | 1.77 | 1.00 | 1.07 |
| TP53 | 1.00 | 0.74 | 0.79 | 1.00 | 0.31 |
| miRNA | Position at UTR | Seed Match | Context ++Score | Context Percentile | Weighted ++Score | Conserved Branch Length |
|---|---|---|---|---|---|---|
| Hsa-miR-6875-3p | 15–21 | 7mer-1A | −0.16 | 91 | −0.16 | 0 |
| Hsa -miR-4659b-3p | 15–21 | 7mer-1A | −0.17 | 91 | −0.17 | 0 |
| Hsa-miR-4659a-3p | 15–21 | 7mer-1A | −0.17 | 91 | −0.17 | 0 |
| Hsa -miR-4520-5p | 24–30 | 7mer-1A | −0.49 | 96 | −0.49 | 0 |
| Hsa -miR-5696 | 35–41 | 7mer-1A | −0.24 | 98 | −0.24 | 0 |
| Hsa -miR-664b-3p | 35–41 | 7mer-1A | −0.15 | 97 | −0.15 | 0.014 |
| Hsa -miR-579-3p | 35–41 | 7mer-1A | −0.15 | 96 | −0.15 | 0.014 |
| Hsa -miR-6853-3p | 36–42 | 7mer-m8 | −0.43 | 99 | −0.43 | 0 |
| Hsa s-miR-3163 | 44–50 | 7mer-1A | −0.07 | 97 | −0.07 | 0 |
| Hsa -miR-340-5p | 45–51 | 7mer-1A | −0.11 | 97 | −0.11 | 0.252 |
| Hsa -miR-186-5p | 54–60 | 7mer-m8 | −0.26 | 99 | −0.26 | 0.239 |
| Has-miR-4477a | 60–66 | 7mer-1A | −0.17 | 97 | −0.17 | 0 |
| Hsa -miR-548e-5p | 75–81 | 7mer-m8 | −0.22 | 98 | −0.22 | 0 |
| Hsa -miR-202-3p | 87–93 | 7mer-m8 | −0.45 | 98 | −0.45 | 0.014 |
| Hsa -miR-3173-3p | 90–96 | 7mer-m8 | 0.48 | 99 | 0.48 | 0.085 |
| Hsa -miR-6891-5p | 90–96 | 7mer-m8 | 0.48 | 99 | 0.48 | 0.085 |
| Hsa -miR-186-3p | 94–100 | 7mer-m8 | −0.3 | 99 | −0.3 | 0 |
| Hsa -miR-3136-3p | 96–103 | 8mer | −1.02 | 99 | −1.02 | 0 |
| Hsa -miR-7155-3p | 96–103 | 8mer | −1.02 | 99 | −1.02 | 0 |
| Hsa -miR-328-3p | 98–104 | 7mer-1A | −0.44 | 98 | −0.44 | 0.133 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Othumpangat, S.; Noti, J.D. β-Defensin-1 Regulates Influenza Virus Infection in Human Bronchial Epithelial Cells through the STAT3 Signaling Pathway. Pathogens 2023, 12, 123. https://doi.org/10.3390/pathogens12010123
Othumpangat S, Noti JD. β-Defensin-1 Regulates Influenza Virus Infection in Human Bronchial Epithelial Cells through the STAT3 Signaling Pathway. Pathogens. 2023; 12(1):123. https://doi.org/10.3390/pathogens12010123
Chicago/Turabian StyleOthumpangat, Sreekumar, and John D. Noti. 2023. "β-Defensin-1 Regulates Influenza Virus Infection in Human Bronchial Epithelial Cells through the STAT3 Signaling Pathway" Pathogens 12, no. 1: 123. https://doi.org/10.3390/pathogens12010123
APA StyleOthumpangat, S., & Noti, J. D. (2023). β-Defensin-1 Regulates Influenza Virus Infection in Human Bronchial Epithelial Cells through the STAT3 Signaling Pathway. Pathogens, 12(1), 123. https://doi.org/10.3390/pathogens12010123