
Diversity of Tick-Borne Pathogens in Tick Larvae Feeding on Breeding Birds in France

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Bird Capture and Tick Collection
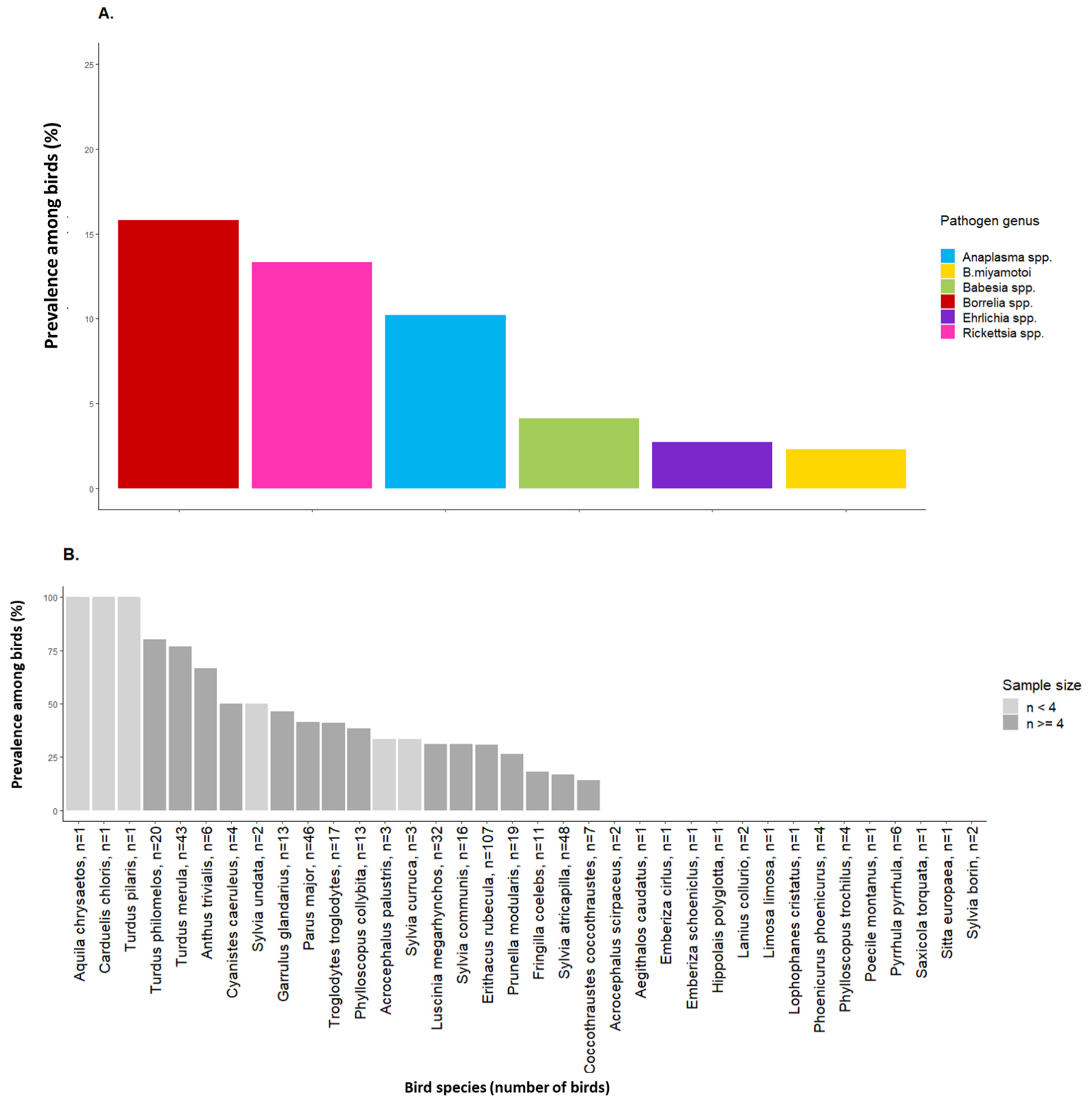
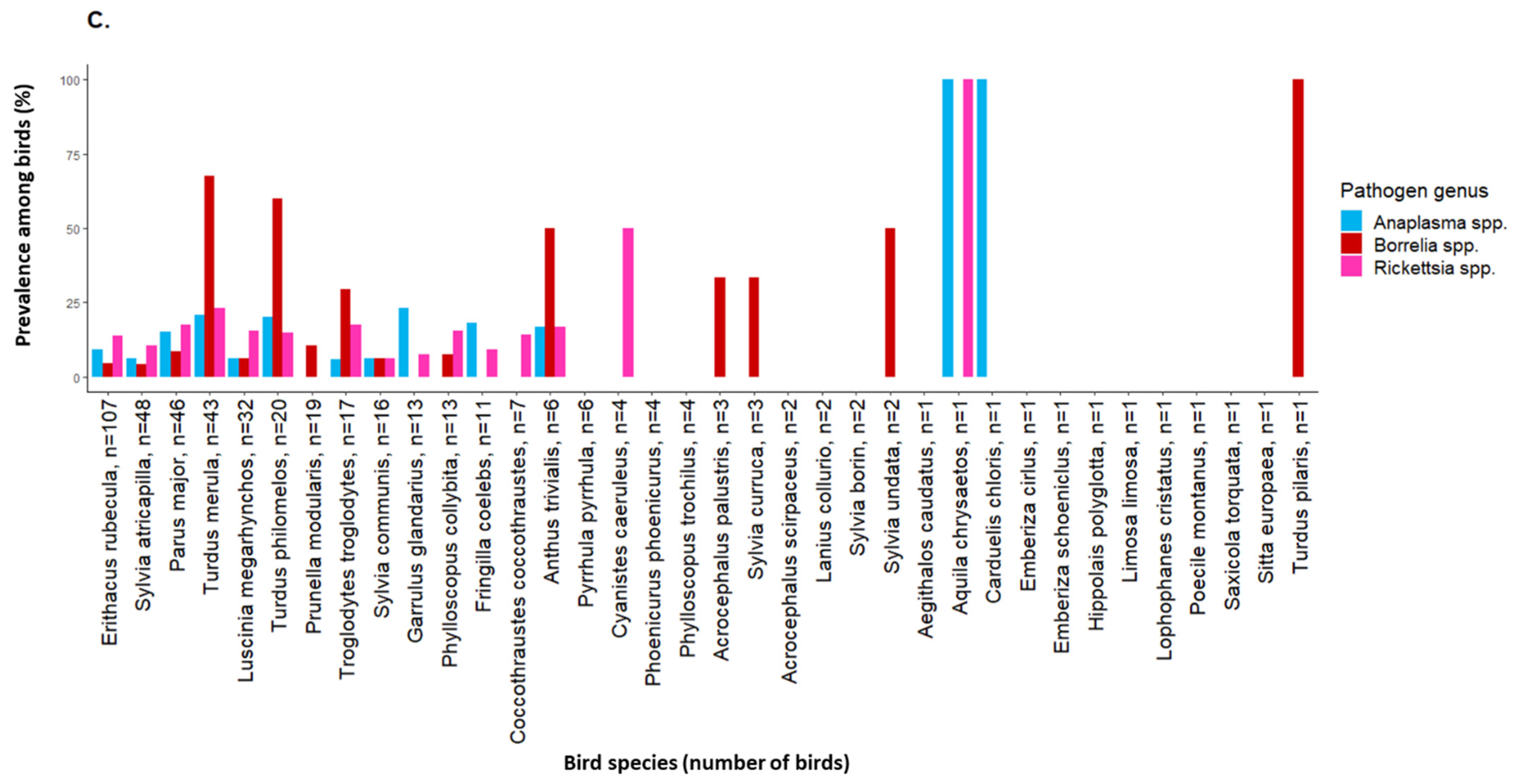
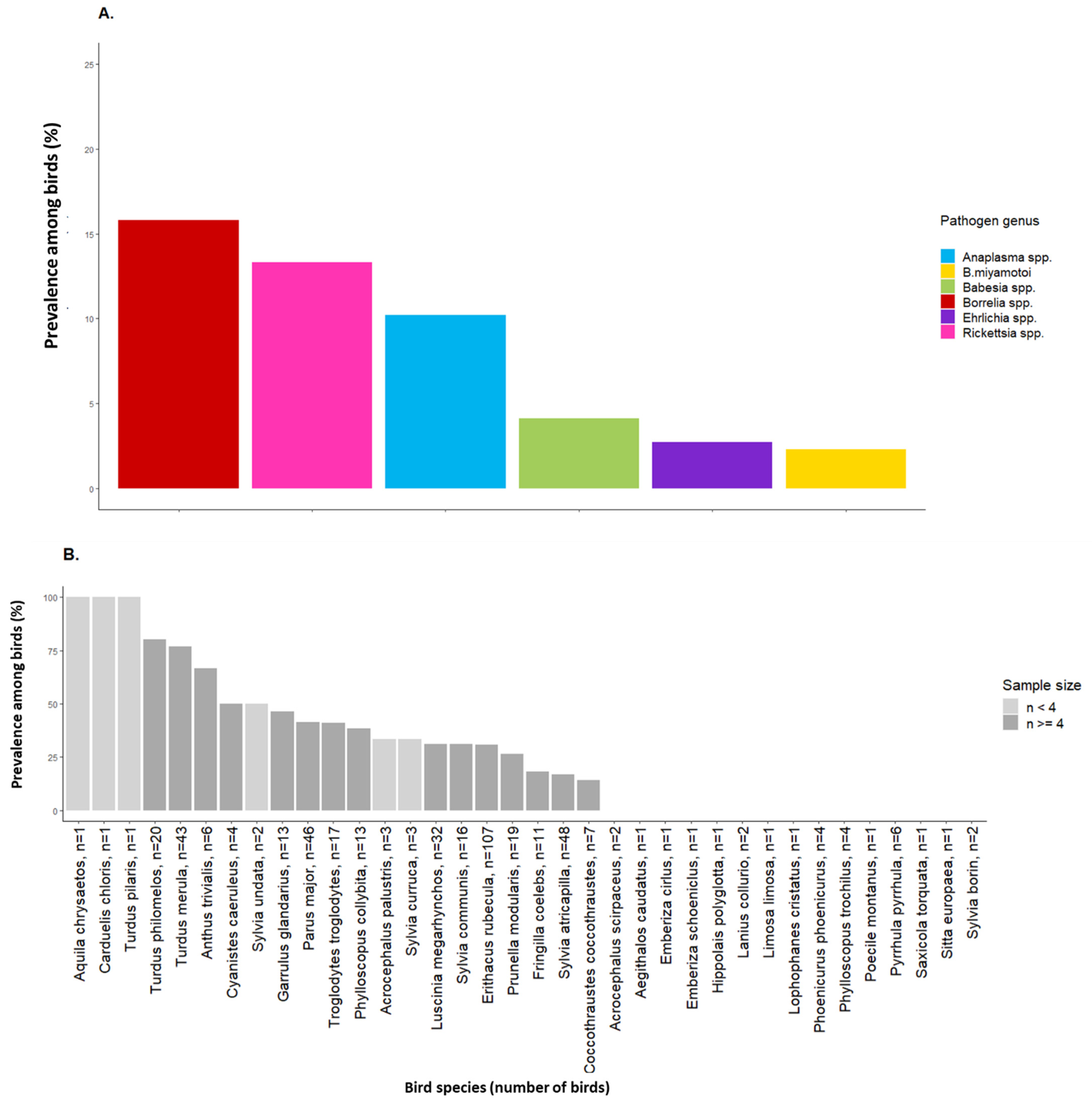
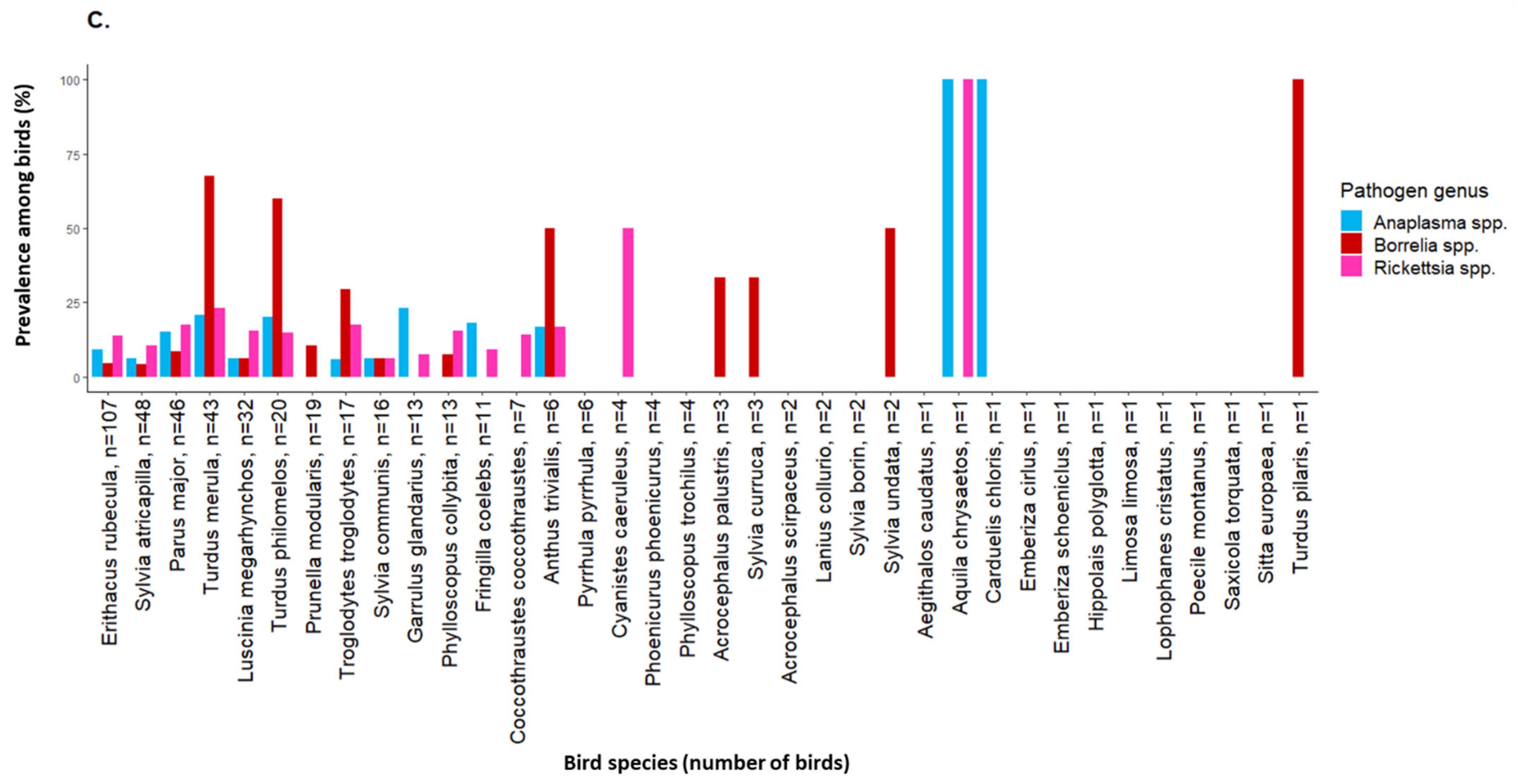
2.2. Tick-Borne Pathogen Infection Rates in Engorged Larvae and Prevalence among Birds
3. Discussion
4. Materials and Methods
4.1. Bird Capture and Tick Collection
4.2. Morphological Tick Identification
4.3. DNA Extraction and Pre-Amplification
4.4. Detection of Tick-Borne Pathogens: DNA Microfluidic Real-Time PCR
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; Sonenshine, D.E. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slowik, T.J.; Lane, R.S. Feeding preferences of the immature stages of three western north American ixodid ticks (Acari) for avian, reptilian, or rodent hosts. J. Med. Entomol. 2009, 46, 115–122. [Google Scholar] [CrossRef] [PubMed]
- McCoy, K.D.; Boulanger, N. Tiques Et Maladies à Tiques: BIOLOGIE, Écologie Évolutive, Épidémiologie; IRD Editions (Collection Didactiques): Marseille, France, 2015. [Google Scholar]
- Rataud, A.; Henry, P.Y.; Moutailler, S.; Marsot, M. Research effort on birds’ reservoir host potential for Lyme borreliosis: A systematic review and perspectives. Transbound Emerg. Dis. 2021, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Falchi, A.; Dantas-Torres, F.; Lorusso, V.; Malia, E.; Lia, R.P.; Otranto, D. Autochthonous and migratory birds as a dispersion source for Ixodes ricinus in southern Italy. Exp. Appl. Acarol. 2012, 58, 167–174. [Google Scholar] [CrossRef]
- Klitgaard, K.; Hojgaard, J.; Isbrand, A.; Madsen, J.J.; Thorup, K.; Bodker, R. Screening for multiple tick-borne pathogens in Ixodes ricinus ticks from birds in Denmark during spring and autumn migration seasons. Ticks Tick-Borne Dis. 2019, 10, 546–552. [Google Scholar] [CrossRef]
- Mysterud, A.; Heylen, D.J.A.; Matthysen, E.; Garcia, A.L.; Jore, S.; Viljugrein, H. Lyme neuroborreliosis and bird populations in northern Europe. Proceedings Biol. Sci. 2019, 286, 20190759. [Google Scholar] [CrossRef] [Green Version]
- Marsot, M.; Henry, P.Y.; Vourc’h, G.; Gasqui, P.; Ferquel, E.; Laignel, J.; Grysan, M.; Chapuis, J.L. Which forest bird species are the main hosts of the tick, Ixodes ricinus, the vector of Borrelia burgdorferi sensu lato, during the breeding season? Int. J. Parasitol. 2012, 42, 781–788. [Google Scholar] [CrossRef]
- Agoulon, A.; Hoch, T.; Heylen, D.; Chalvet-Monfray, K.; Plantard, O. Unravelling the phenology of Ixodes frontalis, a common but understudied tick species in Europe. Ticks Tick-Borne Dis. 2019, 10, 505–512. [Google Scholar] [CrossRef]
- Kaiser, A.; Seitz, A.; Strub, O. Prevalence of Borrelia burgdorferi sensu lato in the nightingale (Luscinia megarhynchos) and other passerine birds. Int. J. Med. Microbiol. IJMM 2002, 291 Suppl 33, 75–79. [Google Scholar] [CrossRef]
- Ciebiera, O.; Jerzak, L.; Nowak-Chmura, M.; Bochenski, M. Ticks (Acari: Ixodida) on birds (Aves) migrating through the Polish Baltic coast. Exp. Appl. Acarol. 2019, 77, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Norte, A.C.; da Silva, L.P.; Tenreiro, P.J.; Felgueiras, M.S.; Araujo, P.M.; Lopes, P.B.; Matos, C.; Rosa, A.; Ferreira, P.J.; Encarnacao, P.; et al. Patterns of tick infestation and their Borrelia burgdorferi s.l. infection in wild birds in Portugal. Ticks Tick-Borne Dis. 2015, 6, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Norte, A.C.; de Carvalho, I.L.; Ramos, J.A.; Goncalves, M.; Gern, L.; Nuncio, M.S. Diversity and seasonal patterns of ticks parasitizing wild birds in western Portugal. Exp. Appl. Acarol. 2012, 58, 327–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heylen, D.; Fonville, M.; Docters van Leeuwen, A.; Stroo, A.; Duisterwinkel, M.; van Wieren, S.; Diuk-Wasser, M.; de Bruin, A.; Sprong, H. Pathogen communities of songbird-derived ticks in Europe’s low countries. Parasites Vectors 2017, 10, 497. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Flaisz, B.; Takacs, N.; Kontschan, J.; Csorgo, T.; Csipak, A.; Jaksa, B.R.; Kovats, D. Bird ticks in Hungary reflect western, southern, eastern flyway connections and two genetic lineages of Ixodes frontalis and Haemaphysalis concinna. Parasites Vectors 2016, 9, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keskin, A.; Erciyas-Yavuz, K. A Preliminary Investigation on Ticks (Acari: Ixodidae) Infesting Birds in Kizilirmak Delta, Turkey. J. Med. Entomol. 2016, 53, 217–220. [Google Scholar] [CrossRef]
- Kocianova, E.; Rusnakova Taragelova, V.; Harustiakova, D.; Spitalska, E. Seasonal infestation of birds with immature stages of Ixodes ricinus and Ixodes arboricola. Ticks Tick-Borne Dis. 2017, 8, 423–431. [Google Scholar] [CrossRef]
- Pascucci, I.; Di Domenico, M.; Capobianco Dondona, G.; Di Gennaro, A.; Polci, A.; Capobianco Dondona, A.; Mancuso, E.; Camma, C.; Savini, G.; Cecere, J.G.; et al. Assessing the role of migratory birds in the introduction of ticks and tick-borne pathogens from African countries: An Italian experience. Ticks Tick-Borne Dis. 2019, 10, 101272. [Google Scholar] [CrossRef]
- Nowak-Chmura, M. Ixodes eldaricus Djaparidze, 1950 (Ixodidae) on migrating birds—Reported first time in Poland. Vet. Parasitol. 2012, 186, 399–402. [Google Scholar] [CrossRef]
- Heylen, D.; Tijsse, E.; Fonville, M.; Matthysen, E.; Sprong, H. Transmission dynamics of Borrelia burgdorferi s.l. in a bird tick community. Environ. Microbiol. 2013, 15, 663–673. [Google Scholar] [CrossRef]
- Berthová, L.; Slobodník, V.; Slobodník, R.; Olekšák, M.; Sekeyová, Z.; Svitálková, Z.; Kazimírová, M.; Špitalská, E. The natural infection of birds and ticks feeding on birds with Rickettsia spp. and Coxiella burnetii in Slovakia. Exp. Appl. Acarol. 2016, 68, 299–314. [Google Scholar] [CrossRef]
- Wallménius, K.; Barboutis, C.; Fransson, T.; Jaenson, T.G.; Lindgren, P.E.; Nyström, F.; Olsen, B.; Salaneck, E.; Nilsson, K. Spotted fever Rickettsia species in Hyalomma and Ixodes ticks infesting migratory birds in the European Mediterranean area. Parasites Vectors 2014, 7, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capligina, V.; Salmane, I.; Keiss, O.; Vilks, K.; Japina, K.; Baumanis, V.; Ranka, R. Prevalence of tick-borne pathogens in ticks collected from migratory birds in Latvia. Ticks Tick-Borne Dis. 2014, 5, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Dubska, L.; Literak, I.; Kverek, P.; Roubalova, E.; Kocianova, E.; Taragelova, V. Tick-borne zoonotic pathogens in ticks feeding on the common nightingale including a novel strain of Rickettsia sp. Ticks Tick-Borne Dis. 2012, 3, 265–268. [Google Scholar] [CrossRef]
- Elfving, K.; Olsen, B.; Bergström, S.; Waldenström, J.; Lundkvist, A.; Sjöstedt, A.; Mejlon, H.; Nilsson, K. Dissemination of spotted fever rickettsia agents in Europe by migrating birds. PLoS ONE 2010, 5, e8572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, R.I.; Mainwaring, M.C.; Du Feu, R. Detection of spotted fever group Rickettsia spp. from bird ticks in the U.K. Med. Vet. Entomol. 2010, 24, 340–343. [Google Scholar] [CrossRef]
- Hornok, S.; Boldogh, S.A.; Takács, N.; Juhász, A.; Kontschán, J.; Földi, D.; Koleszár, B.; Morandini, P.; Gyuranecz, M.; Szekeres, S. Anaplasmataceae closely related to Ehrlichia chaffeensis and Neorickettsia helminthoeca from birds in Central Europe, Hungary. Antonie Van Leeuwenhoek 2020, 113, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Lommano, E.; Dvorak, C.; Vallotton, L.; Jenni, L.; Gern, L. Tick-borne pathogens in ticks collected from breeding and migratory birds in Switzerland. Ticks Tick-Borne Dis. 2014, 5, 871–882. [Google Scholar] [CrossRef]
- Csank, T.; Bhide, K.; Bencúrová, E.; Dolinská, S.; Drzewnioková, P.; Major, P.; Korytár, Ľ.; Bocková, E.; Bhide, M.; Pistl, J. Detection of West Nile virus and tick-borne encephalitis virus in birds in Slovakia, using a universal primer set. Arch. Virol. 2016, 161, 1679–1683. [Google Scholar] [CrossRef] [Green Version]
- Kazarina, A.; Japina, K.; Keiss, O.; Salmane, I.; Bandere, D.; Capligina, V.; Ranka, R. Detection of tick-borne encephalitis virus in I. ricinus ticks collected from autumn migratory birds in Latvia. Ticks Tick-Borne Dis. 2015, 6, 178–180. [Google Scholar] [CrossRef]
- Mancuso, E.; Toma, L.; Polci, A.; d’Alessio, S.G.; Di Luca, M.; Orsini, M.; Di Domenico, M.; Marcacci, M.; Mancini, G.; Spina, F.; et al. Crimean-Congo hemorrhagic fever virus genome in tick from migratory bird, Italy. Emerg. Infect. Dis. 2019, 25, 1418–1420. [Google Scholar] [CrossRef] [Green Version]
- Richter, D.; Debski, A.; Hubalek, Z.; Matuschka, F.-R. Absence of Lyme disease spirochetes in larval Ixodes ricinus ticks. Vector-Borne Zoonotic Dis. 2012, 12, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Hauck, D.; Jordan, D.; Springer, A.; Schunack, B.; Pachnicke, S.; Fingerle, V.; Strube, C. Transovarial transmission of Borrelia spp., Rickettsia spp. and Anaplasma phagocytophilum in Ixodes ricinus under field conditions extrapolated from DNA detection in questing larvae. Parasites Vectors 2020, 13, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtenbach, K.; Hanincová, K.; Tsao, J.I.; Margos, G.; Fish, D.; Ogden, N.H. Fundamental processes in the evolutionary ecology of Lyme borreliosis. Nat. Rev. Microbiol. 2006, 4, 660–669. [Google Scholar] [CrossRef]
- Pajoro, M.; Pistone, D.; Boccazzi, I.V.; Mereghetti, V.; Bandi, C.; Fabbi, M.; Scattorin, F.; Sassera, D.; Montagna, M. Molecular screening for bacterial pathogens in ticks (Ixodes ricinus) collected on migratory birds captured in northern Italy. Folia Parasitol. 2018, 65, 008. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.N.; Jenkins, A.; Kjelland, V. Tick-borne pathogens in Ixodes ricinus ticks collected from migratory birds in southern Norway. PLoS ONE 2020, 15, e0230579. [Google Scholar] [CrossRef] [Green Version]
- Dubska, L.; Literak, I.; Kocianova, E.; Taragelova, V.; Sverakova, V.; Sychra, O.; Hromadko, M. Synanthropic birds influence the distribution of Borrelia species: Analysis of Ixodes ricinus ticks feeding on passerine birds. Appl. Environ. Microbiol. 2011, 77, 1115–1117. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.; Kahl, O.; Zintl, A. What do we still need to know about Ixodes ricinus? Ticks Tick-Borne Dis. 2021, 12, 101682. [Google Scholar] [CrossRef]
- Paulauskas, A.; Sakalauskas, P.; Kaminskienė, E.; Šimkevičius, K.; Kibiša, A.; Radzijevskaja, J. First record of Haemaphysalis concinna (Acari: Ixodidae) in Lithuania. Ticks Tick-Borne Dis. 2020, 11, 101460. [Google Scholar] [CrossRef]
- Stanko, M.; Derdáková, M.; Špitalská, E.; Kazimírová, M. Ticks and their epidemiological role in Slovakia: From the past till present. Biologia 2021, 77, 1575–1610. [Google Scholar] [CrossRef]
- Heylen, D.J.; Van Oosten, A.R.; Devriendt, N.; Elst, J.; De Bruyn, L.; Matthysen, E. Seasonal feeding activity of the tree-hole tick, Ixodes arboricola. Parasitology 2014, 141, 1044–1051. [Google Scholar] [CrossRef] [Green Version]
- Rubel, F.; Brugger, K.; Walter, M.; Vogelgesang, J.R.; Didyk, Y.M.; Fu, S.; Kahl, O. Geographical distribution, climate adaptation and vector competence of the Eurasian hard tick Haemaphysalis concinna. Ticks Tick-Borne Dis. 2018, 9, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- Stanek, G.; Wormser, G.P.; Gray, J.; Strle, F. Lyme borreliosis. Lancet 2012, 379, 461–473. [Google Scholar] [CrossRef]
- Ben, I.; Lozynskyi, I. Prevalence of Anaplasma phagocytophilum in Ixodes ricinus and Dermacentor reticulatus and coinfection with Borrelia burgdorferi and Tick-Borne Encephalitis virus in western Ukraine. Vector Borne Zoonotic Dis. 2019, 19, 793–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stańczak, J.; Biernat, B.; Racewicz, M.; Zalewska, M.; Matyjasek, A. Prevalence of different Rickettsia spp. in Ixodes ricinus and Dermacentor reticulatus ticks (Acari: Ixodidae) in north-eastern Poland. Ticks Tick-Borne Dis. 2018, 9, 427–434. [Google Scholar] [CrossRef]
- Welc-Falęciak, R.; Bajer, A.; Paziewska-Harris, A.; Baumann-Popczyk, A.; Siński, E. Diversity of Babesia in Ixodes ricinus ticks in Poland. Adv. Med. Sci. 2012, 57, 364–369. [Google Scholar] [CrossRef]
- Wagemakers, A.; Jahfari, S.; de Wever, B.; Spanjaard, L.; Starink, M.V.; de Vries, H.J.C.; Sprong, H.; Hovius, J.W. Borrelia miyamotoi in vectors and hosts in The Netherlands. Ticks Tick-Borne Dis. 2017, 8, 370–374. [Google Scholar] [CrossRef]
- Špitalská, E.; Literák, I.; Sparagano, O.A.E.; Golovchenko, M.; Kocianová, E. Ticks (Ixodidae) from passerine birds in the Carpathian region. Wien. Klin. Wochenschr. 2006, 118, 759–764. [Google Scholar] [CrossRef]
- Gehringer, H.; Schacht, E.; Maylaender, N.; Zeman, E.; Kaysser, P.; Oehme, R.; Pluta, S.; Splettstoesser, W.D. Presence of an emerging subclone of Francisella tularensis holarctica in Ixodes ricinus ticks from south-western Germany. Ticks Tick-Borne Dis. 2013, 4, 93–100. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Straube, E.; Neubauer, H.; Schmoock, G. Coxiella burnetii and coinfections in Ixodes ricinus ticks in Central Germany. Vector Borne Zoonotic Dis. 2011, 11, 1205–1207. [Google Scholar] [CrossRef]
- Franke, J.; Fritzsch, J.; Tomaso, H.; Straube, E.; Dorn, W.; Hildebrandt, A. Coexistence of pathogens in host-seeking and feeding ticks within a single natural habitat in Central Germany. Appl. Environ. Microbiol. 2010, 76, 6829–6836. [Google Scholar] [CrossRef] [Green Version]
- Toma, L.; Mancini, F.; Di Luca, M.; Cecere, J.G.; Bianchi, R.; Khoury, C.; Quarchioni, E.; Manzia, F.; Rezza, G.; Ciervo, A. Detection of microbial agents in ticks collected from migratory birds in central Italy. Vector Borne Zoonotic Dis. 2014, 14, 199–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitalska, E.; Literak, I.; Kocianova, E.; Taragel’ova, V. The importance of Ixodes arboricola in transmission of Rickettsia spp., Anaplasma phagocytophilum, and Borrelia burgdorferi sensu lato in the Czech Republic, Central Europe. Vector Borne Zoonotic Dis. 2011, 11, 1235–1241. [Google Scholar] [CrossRef]
- Van Oosten, A.R.; Heylen, D.J.; Matthysen, E. Host specificity of a bird-specialised endophilic ectoparasite, the tree-hole tick Ixodes arboricola. Parasitol. Res. 2014, 113, 4397–4405. [Google Scholar] [CrossRef]
- Palomar, A.M.; Portillo, A.; Santibáñez, P.; Mazuelas, D.; Roncero, L.; García-Álvarez, L.; Santibáñez, S.; Gutiérrez, Ó.; Oteo, J.A. Detection of tick-borne Anaplasma bovis, Anaplasma phagocytophilum and Anaplasma centrale in Spain. Med. Vet. Entomol. 2015, 29, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Tijsse-Klasen, E.; Hansford, K.M.; Jahfari, S.; Phipps, P.; Sprong, H.; Medlock, J.M. Spotted fever group rickettsiae in Dermacentor reticulatus and Haemaphysalis punctata ticks in the UK. Parasites Vectors 2013, 6, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phipps, L.P.; Hansford, K.M.; Hernández-Triana, L.M.; Golding, M.; McGinley, L.; Folly, A.J.; Vaux, A.G.C.; de Marco, M.F.; Carter, D.P.; Medlock, J.M.; et al. Detection of Borrelia and Babesia species in Haemaphysalis punctata ticks sampled in Southern England. Ticks Tick-Borne Dis. 2022, 13, 101902. [Google Scholar] [CrossRef]
- Capek, M.; Literak, I.; Kocianova, E.; Sychra, O.; Najer, T.; Trnka, A.; Kverek, P. Ticks of the Hyalomma marginatum complex transported by migratory birds into Central Europe. Ticks Tick-Borne Dis. 2014, 5, 489–493. [Google Scholar] [CrossRef]
- Pereira, A.; Parreira, R.; Cotão, A.J.; Nunes, M.; Vieira, M.L.; Azevedo, F.; Campino, L.; Maia, C. Tick-borne bacteria and protozoa detected in ticks collected from domestic animals and wildlife in central and southern Portugal. Ticks Tick-Borne Dis. 2018, 9, 225–234. [Google Scholar] [CrossRef]
- Norte, A.C.; Margos, G.; Becker, N.S.; Albino Ramos, J.; Nuncio, M.S.; Fingerle, V.; Araujo, P.M.; Adamik, P.; Alivizatos, H.; Barba, E.; et al. Host dispersal shapes the population structure of a tick-borne bacterial pathogen. Mol. Ecol. 2019, 29, 485–501. [Google Scholar] [CrossRef]
- Mtierová, Z.; Derdáková, M.; Chvostáč, M.; Didyk, Y.M.; Mangová, B.; Rusňáková Tarageľová, V.; Selyemová, D.; Šujanová, A.; Václav, R. Local population structure and seasonal variability of Borrelia garinii genotypes in Ixodes ricinus ticks, Slovakia. Int. J. Environ. Res. Public Health 2020, 17, 3607. [Google Scholar] [CrossRef]
- Norte, A.C.; Ramos, J.A.; Gern, L.; Nuncio, M.S.; Lopes de Carvalho, I. Birds as reservoirs for Borrelia burgdorferi s.l. in Western Europe: Circulation of B. turdi and other genospecies in bird-tick cycles in Portugal. Environ. Microbiol. 2013, 15, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Kurtenbach, K.; De Michelis, S.; Etti, S.; Schäfer, S.M.; Sewell, H.-S.; Brade, V.; Kraiczy, P. Host association of Borrelia burgdorferi sensu lato–the key role of host complement. Trends Microbiol. 2002, 10, 74–79. [Google Scholar] [CrossRef]
- Humair, P.F.; Postic, D.; Wallich, R.; Gern, L. An avian reservoir (Turdus merula) of the Lyme borreliosis spirochetes. Zent. Fur Bakteriol. Int. J. Med. Microbiol. 1998, 287, 521–538. [Google Scholar] [CrossRef]
- Gern, L.; Rais, O. Efficient transmission of Borrelia burgdorferi between cofeeding Ixodes ricinus ticks (Acari: Ixodidae). J. Med. Entomol. 1996, 33, 189–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randolph, S.; Gern, L.; Nuttall, P. Co-feeding ticks: Epidemiological significance for tick-borne pathogen transmission. Parasitol. Today 1996, 12, 472–479. [Google Scholar] [CrossRef]
- Gryczynska, A.; Zgodka, A.; PLoSki, R.; Siemiatkowski, M. Borrelia burgdorferi sensu lato infection in passerine birds from the Mazurian Lake region (Northeastern Poland). Avian Pathol. 2004, 33, 69–75. [Google Scholar] [CrossRef]
- Wilhelmsson, P.; Jaenson, T.G.T.; Olsen, B.; Waldenström, J.; Lindgren, P.E. Migratory birds as disseminators of ticks and the tick-borne pathogens Borrelia bacteria and tick-borne encephalitis (TBE) virus: A seasonal study at Ottenby Bird Observatory in South-eastern Sweden. Parasites Vectors 2020, 13, 607. [Google Scholar] [CrossRef]
- Hornok, S.; Kováts, D.; Csörgő, T.; Meli, M.L.; Gönczi, E.; Hadnagy, Z.; Takács, N.; Farkas, R.; Hofmann-Lehmann, R. Birds as potential reservoirs of tick-borne pathogens: First evidence of bacteraemia with Rickettsia helvetica. Parasites Vectors 2014, 7, 128. [Google Scholar] [CrossRef] [Green Version]
- Biernat, B.; Stańczak, J.; Michalik, J.; Sikora, B.; Wierzbicka, A. Prevalence of infection with Rickettsia helvetica in Ixodes ricinus ticks feeding on non-rickettsiemic rodent hosts in sylvatic habitats of west-central Poland. Ticks Tick-Borne Dis. 2016, 7, 135–141. [Google Scholar] [CrossRef]
- Socolovschi, C.; Mediannikov, O.; Raoult, D.; Parola, P. The relationship between spotted fever group Rickettsiae and ixodid ticks. Vet. Res. 2009, 40, 34. [Google Scholar] [CrossRef] [Green Version]
- Hornok, S.; Csörgő, T.; de la Fuente, J.; Gyuranecz, M.; Privigyei, C.; Meli, M.L.; Kreizinger, Z.; Gönczi, E.; Fernández de Mera, I.G.; Hofmann-Lehmann, R. Synanthropic birds associated with high prevalence of tick-borne rickettsiae and with the first detection of Rickettsia aeschlimannii in Hungary. Vector Borne Zoonotic Dis. 2013, 13, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Silva, M.M.; Sousa, R.; Santos, A.S.; Melo, P.; Encarnação, V.; Bacellar, F. Ticks parasitizing wild birds in Portugal: Detection of Rickettsia aeschlimannii, R. helvetica and R. massiliae. Exp. Appl. Acarol. 2006, 39, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Mancini, F.; Ciccozzi, M.; Lo Presti, A.; Cella, E.; Giovanetti, M.; Di Luca, M.; Toma, L.; Bianchi, R.; Khoury, C.; Rezza, G.; et al. Characterization of spotted fever group Rickettsiae in ticks from a city park of Rome, Italy. Ann. Dell’istituto Super. Di Sanita 2015, 51, 284–290. [Google Scholar] [CrossRef]
- Mărcuţan, I.D.; Kalmár, Z.; Ionică, A.M.; D’Amico, G.; Mihalca, A.D.; Vasile, C.; Sándor, A.D. Spotted fever group rickettsiae in ticks of migratory birds in Romania. Parasites Vectors 2016, 9, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martello, E.; Selmi, M.; Ragagli, C.; Ambrogi, C.; Stella, M.C.; Mannelli, A.; Tomassone, L. Rickettsia slovaca in immature Dermacentor marginatus and tissues from Apodemus spp. in the northern Apennines, Italy. Ticks Tick-Borne Dis. 2013, 4, 518–521. [Google Scholar] [CrossRef]
- Hoffman, T.; Wilhelmsson, P.; Barboutis, C.; Fransson, T.; Jaenson, T.G.T.; Lindgren, P.E.; Von Loewenich, F.D.; Lundkvist, Å.; Olsen, B.; Salaneck, E. A divergent Anaplasma phagocytophilum variant in an Ixodes tick from a migratory bird; Mediterranean basin. Infect. Ecol. Epidemiol. 2020, 10, 1729653. [Google Scholar] [CrossRef] [Green Version]
- Palomar, A.M.; Santibanez, P.; Mazuelas, D.; Roncero, L.; Santibanez, S.; Portillo, A.; Oteo, J.A. Role of birds in dispersal of etiologic agents of tick-borne zoonoses, Spain, 2009. Emerg. Infect. Dis. 2012, 18, 1188–1191. [Google Scholar] [CrossRef]
- Johnston, E.; Tsao, J.I.; Muñoz, J.D.; Owen, J. Anaplasma phagocytophilum infection in American robins and gray catbirds: An assessment of reservoir competence and disease in captive wildlife. J. Med. Entomol. 2013, 50, 163–170. [Google Scholar] [CrossRef]
- Keesing, F.; Hersh, M.H.; Tibbetts, M.; McHenry, D.J.; Duerr, S.; Brunner, J.; Killilea, M.; LoGiudice, K.; Schmidt, K.A.; Ostfeld, R.S. Reservoir competence of vertebrate hosts for Anaplasma phagocytophilum. Emerg. Infect. Dis. 2012, 18, 2013–2016. [Google Scholar] [CrossRef]
- Karshima, S.N.; Karshima, M.N.; Ahmed, M.I. Animal reservoirs of zoonotic Babesia species: A global systematic review and meta-analysis of their prevalence, distribution and species diversity. Vet. Parasitol. 2021, 298, 109539. [Google Scholar] [CrossRef]
- Baráková, I.; Derdáková, M.; Selyemová, D.; Chvostáč, M.; Špitalská, E.; Rosso, F.; Collini, M.; Rosà, R.; Tagliapietra, V.; Girardi, M.; et al. Tick-borne pathogens and their reservoir hosts in northern Italy. Ticks Tick-Borne Dis. 2018, 9, 164–170. [Google Scholar] [CrossRef]
- Wilhelmsson, P.; Pawełczyk, O.; Jaenson, T.G.T.; Waldenström, J.; Olsen, B.; Forsberg, P.; Lindgren, P.E. Three Babesia species in Ixodes ricinus ticks from migratory birds in Sweden. Parasites Vectors 2021, 14, 183. [Google Scholar] [CrossRef] [PubMed]
- Monks, D.; Fisher, M.; Forbes, N. Ixodes frontalis and avian tick—related syndrome in the United Kingdom. J. Small Anim. Pract. 2006, 47, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.Z.; André, M.R.; Werther, K.; de Sousa, E.; Gavioli, F.A.; Alves Junior, J.R. Migratory and carnivorous birds in Brazil: Reservoirs for Anaplasma and Ehrlichia species? Vector Borne Zoonotic Dis. 2012, 12, 705–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacchi, A.B.V.; André, M.R.; Calchi, A.C.; de Santi, M.; Guimarães, A.; Pires, J.R.; Baldani, C.D.; Werther, K.; Machado, R.Z. Molecular and serological detection of arthropod-borne pathogens in carnivorous birds from Brazil. Vet. Parasitol. Reg. Stud. Rep. 2021, 23, 100539. [Google Scholar] [CrossRef] [PubMed]
- Heylen, D.; De Coninck, E.; Jansen, F.; Madder, M. Differential diagnosis of three common Ixodes spp. ticks infesting songbirds of Western Europe: Ixodes arboricola, I. frontalis and I. ricinus. Ticks Tick-Borne Dis. 2014, 5, 693–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heylen, D.; Krawczyk, A.; Lopes de Carvalho, I.; Nuncio, M.S.; Sprong, H.; Norte, A.C. Bridging of cryptic Borrelia cycles in European songbirds. Environ. Microbiol. 2017, 19, 1857–1867. [Google Scholar] [CrossRef]
- Tsao, J.I.; Hamer, S.A.; Han, S.; Sidge, J.L.; Hickling, G.J. The contribution of wildlife hosts to the rise of ticks and tick-borne diseases in North America. J. Med. Entomol. 2021, 58, 1565–1587. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa: A guide to species identification; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Pérez-Eid, C. Les Tiques: Identification, Biologie, Importance médicale Et Vétérinaire; Lavoisier: Paris, France, 2007. [Google Scholar]
- Banović, P.; Díaz-Sánchez, A.A.; Galon, C.; Simin, V.; Mijatović, D.; Obregón, D.; Moutailler, S.; Cabezas-Cruz, A. Humans infested with Ixodes ricinus are exposed to a diverse array of tick-borne pathogens in Serbia. Ticks Tick-Borne Dis. 2021, 12, 101609. [Google Scholar] [CrossRef]
- Banović, P.; Díaz-Sánchez, A.A.; Galon, C.; Foucault-Simonin, A.; Simin, V.; Mijatović, D.; Papić, L.; Wu-Chuang, A.; Obregón, D.; Moutailler, S. A One Health approach to study the circulation of tick-borne pathogens: A preliminary study. One Health 2021, 13, 100270. [Google Scholar] [CrossRef]
- Boularias, G.; Azzag, N.; Galon, C.; Šimo, L.; Boulouis, H.J.; Moutailler, S. High-throughput microfluidic real-time PCR for the detection of multiple microorganisms in Ixodid cattle ticks in northeast Algeria. Pathogens 2021, 10, 362. [Google Scholar] [CrossRef] [PubMed]
- Loh, S.-M.; Gofton, A.W.; Lo, N.; Gillett, A.; Ryan, U.M.; Irwin, P.J.; Oskam, C.L. Novel Borrelia species detected in echidna ticks, Bothriocroton concolor, in Australia. Parasites Vectors 2016, 9, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rar, V.A.; Fomenko, N.V.; Dobrotvorsky, A.K.; Livanova, N.N.; Rudakova, S.A.; Fedorov, E.G.; Astanin, V.B.; Morozova, O.V. Tickborne pathogen detection, western Siberia, Russia. Emerg. Infect. Dis. 2005, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [Green Version]
- Masatani, T.; Hayashi, K.; Andoh, M.; Tateno, M.; Endo, Y.; Asada, M.; Kusakisako, K.; Tanaka, T.; Gokuden, M.; Hozumi, N. Detection and molecular characterization of Babesia, Theileria, and Hepatozoon species in hard ticks collected from Kagoshima, the southern region in Japan. Ticks Tick-Borne Dis. 2017, 8, 581–587. [Google Scholar] [CrossRef]
- Bunikis, J.; Garpmo, U.; Tsao, J.; Berglund, J.; Fish, D.; Barbour, A.G. Sequence typing reveals extensive strain diversity of the Lyme borreliosis agents Borrelia burgdorferi in North America and Borrelia afzelii in Europe. Microbiology 2004, 150, 1741–1755. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Life Stage | Male | Female | Nymph | Larva (Engorged) | Unidentified | Total (%) | |
|---|---|---|---|---|---|---|---|
| Species | |||||||
| Genus Ixodes | |||||||
| I. ricinus | 0 | 29 | 1591 | 1167 (1039) | 0 | 2787 (89.5%) | |
| I. frontalis | 0 | 16 | 8 | 2 (2) | 0 | 26 (0.8%) | |
| I. arboricola | 0 | 3 | 17 | 2 (0) | 0 | 22 (0.7%) | |
| I. spp. | 0 | 0 | 0 | 4 (2) | 0 | 4 (0.1%) | |
| Genus Haemaphysalis | |||||||
| H. concinna | 0 | 0 | 5 | 11 (11) | 0 | 16 (0.5%) | |
| H. punctata | 0 | 0 | 1 | 15 (15) | 0 | 16 (0.5%) | |
| H. spp. | 0 | 0 | 0 | 3 (3) | 0 | 3 (0.1%) | |
| Genus Hyalomma | |||||||
| H. spp. | 0 | 0 | 5 | 0 | 0 | 5 (0.2%) | |
| Genus Rhipicephalus | |||||||
| R. spp. | 1 | 1 | 0 | 0 | 0 | 2 (0.06%) | |
| Unidentified a | 0 | 11 | 88 | 81 (34) | 53 | 233 (7.5%) | |
| Total (%) | 1 (0.03%) | 60 (1.9%) | 1715 (55.1%) | 1285 (41.3%) | 53 (1.7%) | 3114 | |
| Pathogen Species | I. ricinus | I. frontalis | Ixodes spp. | H. concinna | H. punctata | Haemaphysalis spp. | Unidentified | Number of Birds with TBP-Positive Larvae | |
|---|---|---|---|---|---|---|---|---|---|
| Pathogen Species | |||||||||
| Genus Anaplasma | |||||||||
| A. phagocytophilum | 5.8 (60) | 0 | 0 | 0 | 6.7 (1) | 0 | 5.9 (2) | 45 | |
| Genus Babesia | 2.3 (24) | 0 | 0 | 0 | 0 | 0 | 2.9 (1) | 18 | |
| B. venatorum | 0.3 (3) | 0 | 0 | 0 | 0 | 0 | 0 | 3 | |
| B. spp. | 2 (21) | 0 | 0 | 0 | 0 | 0 | 2.9 (1) | 15 | |
| Genus Bbsl a | 11.9 (124) | 100 (2) | 50 (1) | 0 | 0 | 33.3 (1) | 2.9 (1) | 70 | |
| B. afzelii | 0.2 (2) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | |
| Bbss b | 0.2 (2) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | |
| B. garinii | 9.2 (96) | 50 (1) | 50 (1) | 0 | 0 | 33.3 (1) | 2.9 (1) | 54 | |
| B. turdi | 0.3 (3) | 50 (1) | 0 | 0 | 0 | 0 | 0 | 3 | |
| B. valaisiana | 1.9 (20) | 0 | 0 | 0 | 0 | 0 | 0 | 15 | |
| B. spp. | 0.1 (1) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| B. miyamotoi | 1.1 (11) | 0 | 0 | 0 | 0 | 0 | 0 | 10 | |
| Genus Ehrlichia | 1.4 (15) | 0 | 0 | 0 | 0 | 0 | 0 | 12 | |
| E. canis | 0.1 (1) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| close to E. chaffeensis | 0.5 (5) | 0 | 0 | 0 | 0 | 0 | 0 | 5 | |
| E. spp. | 0.9 (9) | 0 | 0 | 0 | 0 | 0 | 0 | 9 | |
| Genus Rickettsia | 6.5 (68) | 0 | 0 | 18.2 (2) | 6.7 (1) | 0 | 32.3 (11) | 59 | |
| R. aeschlimannii | 0.4 (4) | 0 | 0 | 0 | 0 | 0 | 0 | 4 | |
| R. helvetica | 4.1 (43) | 0 | 0 | 0 | 0 | 0 | 5.9 (2) | 36 | |
| R. slovaca | 0.1 (1) | 0 | 0 | 0 | 0 | 0 | 26.5 (9) | 2 | |
| R. spp. | 1.9 (20) | 0 | 0 | 18.2 (2) | 6.7 (1) | 0 | 0 | 21 | |
| Total larvae | 1039 | 2 | 2 | 11 | 15 | 3 | 34 | ||
| Total birds | 442 | ||||||||
| Pathogen Genus | Target Gene | Primer Name | Sequence (5’-3’) | Amplicon Size (bp) | T | Reference |
|---|---|---|---|---|---|---|
| Borrelia spp. | FlaB | FlaB280F FlaRL flaB_737F FlaLL | GCAGTTCARTCAGGTAACGG GCAATCATAGCCATTGCAGATTGT GCATCAACTGTRGTTGTAACATTAACAGG ACATATTCAGATGCAGACAGAGGT | 645 407 | 55 59 | [95] |
| Anaplasma spp./Ehrlichia spp. | 16S rRNA | EHR1 F EHR2 R EHR3 F EHR2 R | GAACGAACGCTGGCGGCAAGC AGTA(T/C)CG(A/G)ACCAGATAGCCGC TGCATAGGAATCTACCTAGTAG AGTA(T/C)CG(A/G)ACCAGATAGCCGC | 693 629 | 60 55 | [96] |
| Rickettsia spp. | gltA | Rsfg877 Rsfg1258 | GGG GGC CTG CTC ACG GCG G ATT GCA AAA AGT ACA GTG AAC A- | 381 | 56 | [97] |
| Babesia spp. | 18S rRNA | BTH 18S 1st F BTH 18S 1st R BTH 18S 2nd F BTH 18S 2nd R | GTGAAACTGCGAATGGCTCATTAC AAGTGATAAGGTTCACAAAACTTCCC GGCTCATTACAACAGTTATAGTTTATTTG CGGTCCGAATAATTCACCGGAT | 1500 | 58 | [98] |
| B. miyamotoi | IGS | Bospp-IGS-F Bospp-IGS-R Bospp-IGS-Fi Bospp-IGS-Ri | GTATGTTTAGTGAGGGGGGTG GGATCATAGCTCAGGTGGTTAG AGGGGGGTGAAGTCGTAACAAG GTCTGATAAACCTGAGGTCGGA | 1007 388–685 | 56 58 | [99] |
| Tick species | COI | HCO2198 LCO1490 | TAA ACT TCA GGG TGA CCA AAA AAT CA GGT CAA CAA ATC ATA AAG ATA TTG G | 710 | 48 | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rataud, A.; Galon, C.; Bournez, L.; Henry, P.-Y.; Marsot, M.; Moutailler, S. Diversity of Tick-Borne Pathogens in Tick Larvae Feeding on Breeding Birds in France. Pathogens 2022, 11, 946. https://doi.org/10.3390/pathogens11080946
Rataud A, Galon C, Bournez L, Henry P-Y, Marsot M, Moutailler S. Diversity of Tick-Borne Pathogens in Tick Larvae Feeding on Breeding Birds in France. Pathogens. 2022; 11(8):946. https://doi.org/10.3390/pathogens11080946
Chicago/Turabian StyleRataud, Amalia, Clemence Galon, Laure Bournez, Pierre-Yves Henry, Maud Marsot, and Sara Moutailler. 2022. "Diversity of Tick-Borne Pathogens in Tick Larvae Feeding on Breeding Birds in France" Pathogens 11, no. 8: 946. https://doi.org/10.3390/pathogens11080946
APA StyleRataud, A., Galon, C., Bournez, L., Henry, P.-Y., Marsot, M., & Moutailler, S. (2022). Diversity of Tick-Borne Pathogens in Tick Larvae Feeding on Breeding Birds in France. Pathogens, 11(8), 946. https://doi.org/10.3390/pathogens11080946