
Repurposing Avasimibe to Inhibit Bacterial Glycosyltransferases


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
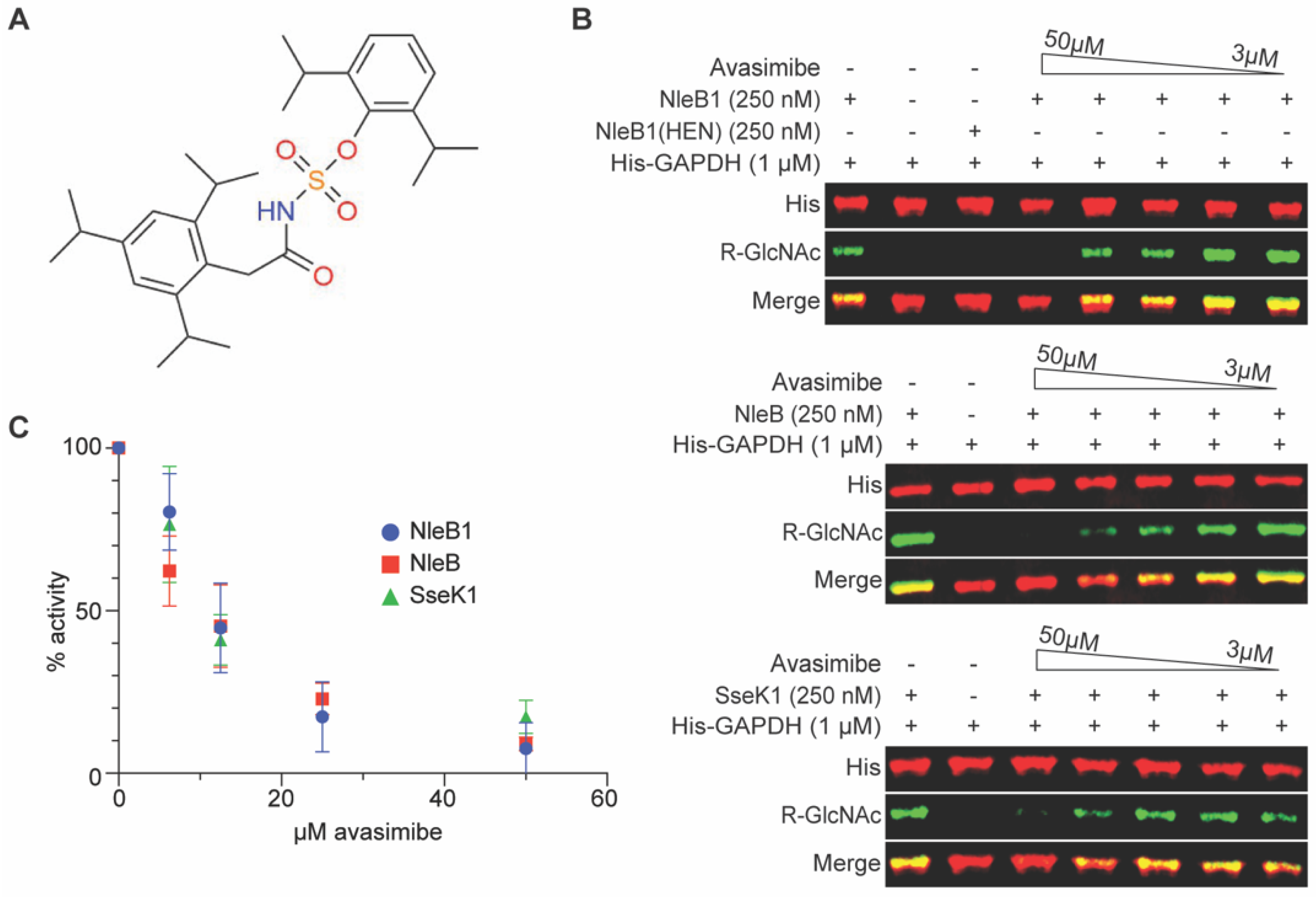
2.1. Avasimibe Inhibits NleB1 and Its Orthologs
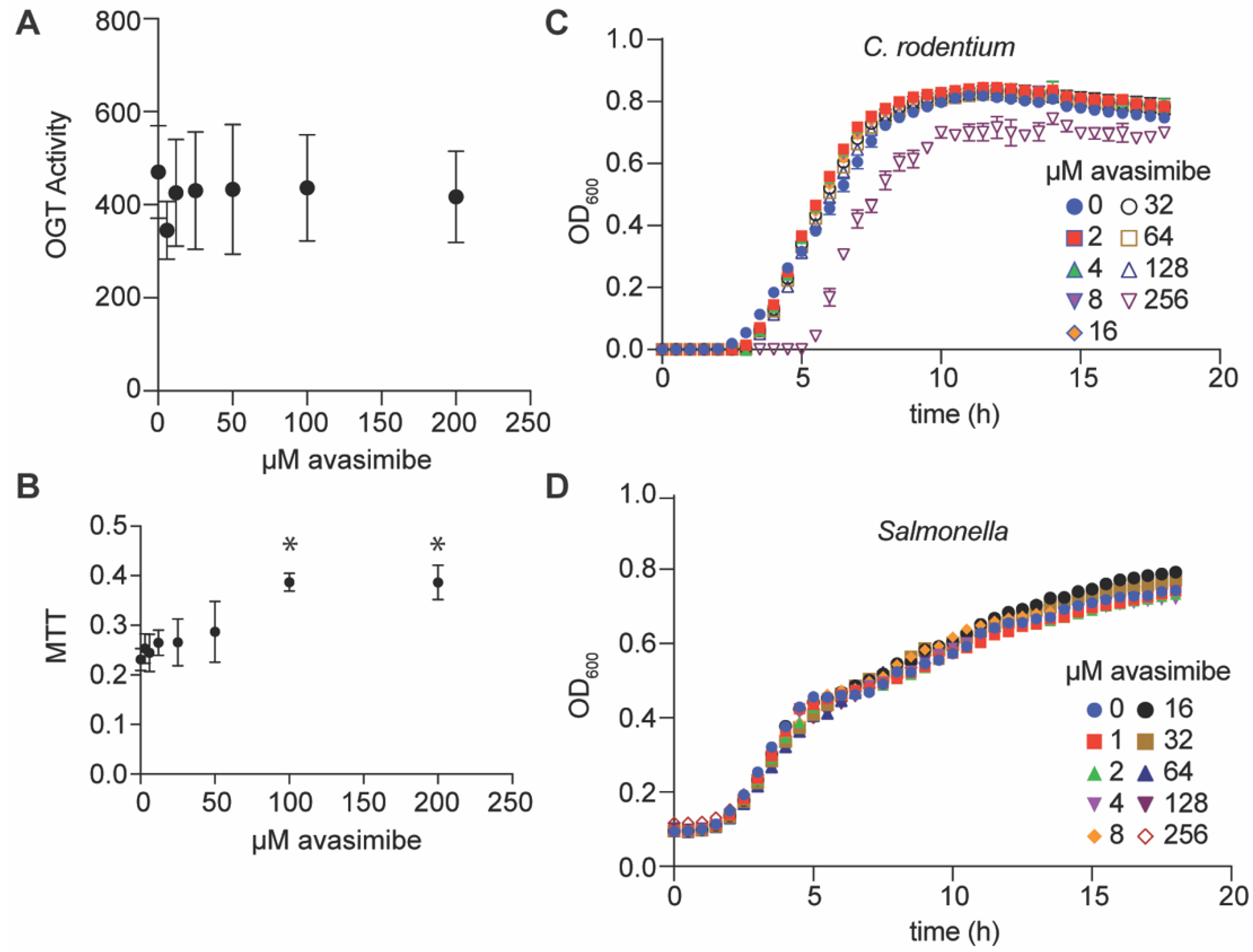
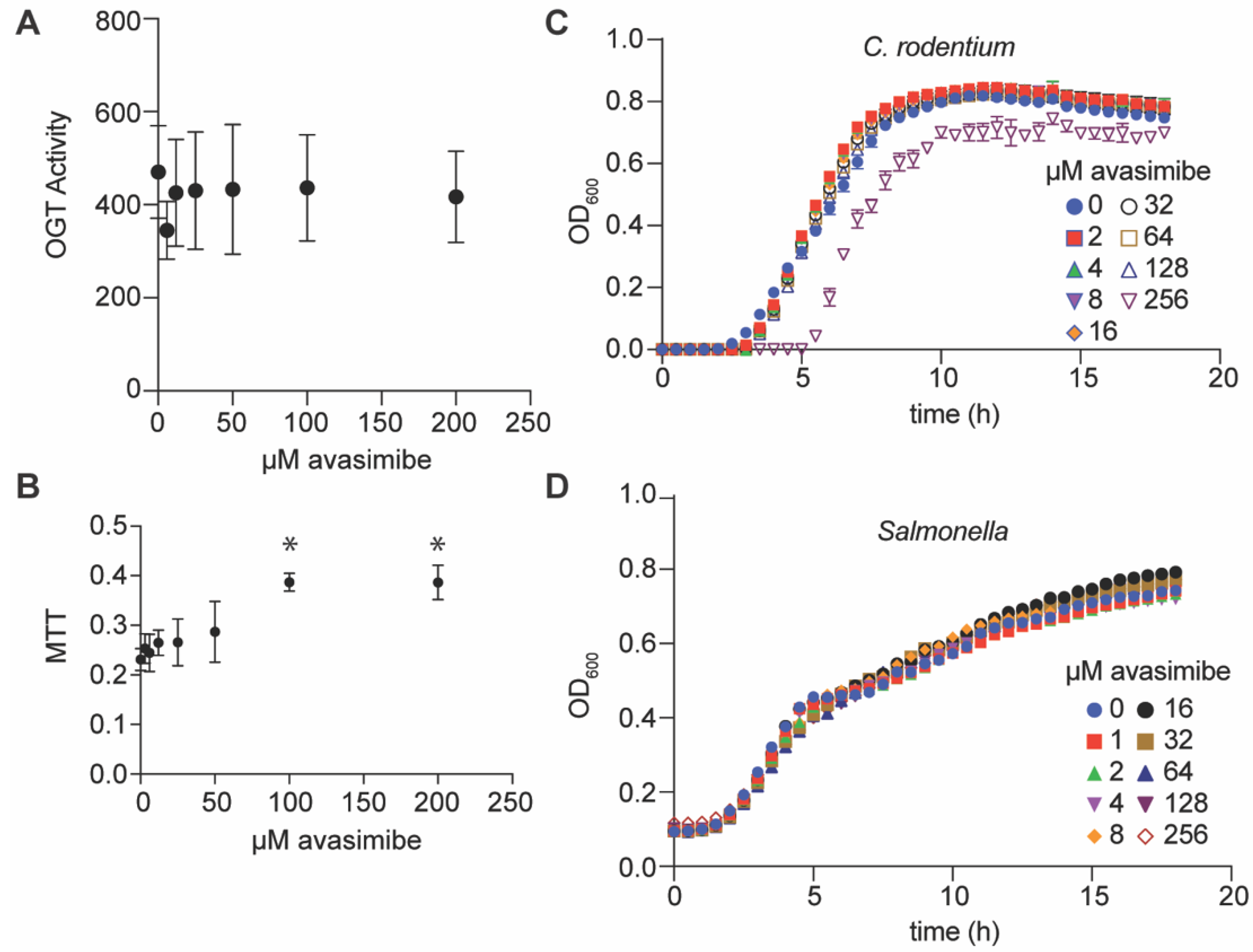
2.2. Avasimibe Does Not Inihibit the Mammalian OGT Enzyme, Is Not Toxic to Mammalian Cells, and Is Neither Bacteriostatic nor Bacteriocidal
2.3. Avasimibe Inhibits Salmonella and C. rodentium Survival In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Protein Purification
4.3. Glycosylation Assays
4.4. OGT Assays
4.5. MTT Assays
4.6. Bacterial Growth Assays
4.7. Macrophage Infections
4.8. Mouse Infections
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- El Qaidi, S.; Chen, K.; Halim, A.; Siukstaite, L.; Rueter, C.; Hurtado-Guerrero, R.; Clausen, H.; Hardwidge, P.R. Nleb/Ssek Effectors from Citrobacter Rodentium, Escherichia Coli, and Salmonella Enterica Display Distinct Differences in Host Substrate Specificity. J. Biol. Chem. 2017, 292, 11423–11430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Qaidi, S.; Scott, N.E.; Hays, M.P.; Geisbrecht, B.V.; Watkins, S.; Hardwidge, P.R. An Intra-Bacterial Activity for a T3ss Effector. Sci. Rep. 2020, 10, 1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Pham, T.H.; Feuerbacher, L.A.; Chen, K.; Hays, M.P.; Singh, G.; Rueter, C.; Hurtado-Guerrero, R.; Hardwidge, P.R. Citrobacter Rodentium Nleb Protein Inhibits Tumor Necrosis Factor (Tnf) Receptor-Associated Factor 3 (Traf3) Ubiquitination to Reduce Host Type I Interferon Production. J. Biol. Chem. 2016, 291, 18232–18238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Wang, X.; Pham, T.H.; Feuerbacher, L.A.; Lubos, M.-L.; Huang, M.; Olsen, R.; Mushegian, A.; Slawson, C.; Hardwidge, P.R. Nleb, a Bacterial Effector with Glycosyltransferase Activity, Targets Gapdh Function to Inhibit Nf-Kappab Activation. Cell Host Microbe 2013, 13, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Park, J.B.; Kim, Y.H.; Yoo, Y.; Kim, J.; Jun, S.-H.; Cho, J.W.; El Qaidi, S.; Walpole, S.; Monaco, S.; García-García, A.A.; et al. Structural Basis for Arginine Glycosylation of Host Substrates by Bacterial Effector Proteins. Nat. Commun. 2018, 9, 4283. [Google Scholar] [CrossRef]
- Li, S.; Zhang, L.; Yao, Q.; Li, L.; Dong, N.; Rong, J.; Gao, W.; Ding, X.; Sun, L.; Chen, X.; et al. Pathogen Blocks Host Death Receptor Signalling by Arginine Glcnacylation of Death Domains. Nature 2013, 501, 242–246. [Google Scholar] [CrossRef]
- Pearson, J.S.; Giogha, C.; Ong, S.Y.; Kennedy, C.L.; Kelly, M.; Robinson, K.S.; Lung, T.W.; Mansell, A.; Riedmaier, P.; Oates, C.V.; et al. A Type Iii Effector Antagonizes Death Receptor Signalling During Bacterial Gut Infection. Nature 2013, 501, 247–251. [Google Scholar] [CrossRef]
- Theuretzbacher, U.; Piddock, L.J.V. Non-Traditional Antibacterial Therapeutic Options and Challenges. Cell Host Microbe 2019, 26, 61–72. [Google Scholar] [CrossRef]
- El Qaidi, S.; Zhu, C.; McDonald, P.; Roy, A.; Maity, P.K.; Rane, D.; Perera, C.; Hardwidge, P.R. High-Throughput Screening for Bacterial Glycosyltransferase Inhibitors. Front. Cell Infect Microbiol. 2018, 8, 435. [Google Scholar] [CrossRef]
- Zhu, C.; El Qaidi, S.; McDonald, P.; Roy, A.; Hardwidge, P. Ym155 Inhibits Nleb and Ssek Arginine Glycosyltransferase Activity. Pathogens 2021, 10, 253. [Google Scholar] [CrossRef]
- Burnett, J.R.; Huff, M.W. Avasimibe Pfizer. Curr. Opin. Investig. Drugs 2002, 3, 1328–1333. [Google Scholar] [PubMed]
- Aquino-Gil, M.; Pierce, A.; Perez-Cervera, Y.; Zenteno, E.; Lefebvre, T. Ogt: A Short Overview of an Enzyme Standing out from Usual Glycosyltransferases. Biochem. Soc. Trans. 2017, 45, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.R.; Wilcox, L.J.; Telford, D.E.; Kleinstiver, S.J.; Barrett, P.H.; Newton, R.S.; Huff, M.W. Inhibition of Acat by Avasimibe Decreases Both Vldl and Ldl Apolipoprotein B Production in Miniature Pigs. J. Lipid Res. 1999, 40, 1317–1327. [Google Scholar] [CrossRef]
- Xiong, K.; Wang, G.; Peng, T.; Zhou, F.; Chen, S.; Liu, W.; Ju, L.; Xiao, Y.; Qian, K.; Wang, X. The Cholesterol Esterification Inhibitor Avasimibe Suppresses Tumour Proliferation and Metastasis Via the E2f-1 Signalling Pathway in Prostate Cancer. Cancer Cell Int. 2021, 21, 461. [Google Scholar] [CrossRef]
- Zhu, Y.; Kim, S.Q.; Zhang, Y.; Liu, Q.; Kim, K.H. Pharmacological Inhibition of Acyl-Coenzyme A:Cholesterol Acyltransferase Alleviates Obesity and Insulin Resistance in Diet-Induced Obese Mice by Regulating Food Intake. Metabolism 2021, 123, 154861. [Google Scholar] [CrossRef]
- Xavier, M.A.; Brust, F.R.; Waldman, J.; Macedo, A.J.; Juliano, M.A.; da Silva Vaz, I., Jr.; Termignoni, C. Interfering with Cholesterol Metabolism Impairs Tick Embryo Development and Turns Eggs Susceptible to Bacterial Colonization. Ticks Tick Borne Dis. 2021, 12, 101790. [Google Scholar] [CrossRef]
- Luo, Y.; Liu, L.; Li, X.; Shi, Y. Avasimibe Inhibits the Proliferation, Migration and Invasion of Glioma Cells by Suppressing Linc00339. Biomed. Pharmacother. 2020, 130, 110508. [Google Scholar] [CrossRef]
- Hu, L.; Li, J.; Cai, H.; Yao, W.; Xiao, J.; Li, Y.-P.; Qiu, X.; Xia, H.; Peng, T. Avasimibe: A Novel Hepatitis C Virus Inhibitor That Targets the Assembly of Infectious Viral Particles. Antiviral Res. 2017, 148, 5–14. [Google Scholar] [CrossRef]
- Bi, M.; Qiao, X.; Zhang, H.; Wu, H.; Gao, Z.; Zhou, H.; Shi, M.; Wang, Y.; Yang, J.; Hu, J.; et al. Effect of Inhibiting Acat-1 Expression on the Growth and Metastasis of Lewis Lung Carcinoma. Oncol. Lett. 2019, 18, 1548–1556. [Google Scholar] [CrossRef]
- Li, J.; Qu, X.; Tian, J.; Zhang, J.-T.; Cheng, J.-X. Cholesterol Esterification Inhibition and Gemcitabine Synergistically Suppress Pancreatic Ductal Adenocarcinoma Proliferation. PLoS ONE 2018, 13, e0193318. [Google Scholar] [CrossRef] [Green Version]
- Delsing, D.J.; Offerman, E.H.; van Duyvenvoorde, W.; van Der Boom, H.; de Wit, E.C.; Gijbels, M.J.; van Der Laarse, A.; Jukema, J.W.; Havekes, L.M.; Princen, H.M. Acyl-Coa:Cholesterol Acyltransferase Inhibitor Avasimibe Reduces Atherosclerosis in Addition to Its Cholesterol-Lowering Effect in Apoe*3-Leiden Mice. Circulation 2001, 103, 1778–1786. [Google Scholar] [CrossRef] [PubMed]
- Sahi, J.; Milad, M.A.; Zheng, X.; Rose, K.A.; Wang, H.; Stilgenbauer, L.; Gilbert, D.; Jolley, S.; Stern, R.H.; Lecluyse, E.L. Avasimibe Induces Cyp3a4 and Multiple Drug Resistance Protein 1 Gene Expression through Activation of the Pregnane X Receptor. J. Pharmacol. Exp. Ther. 2003, 306, 1027–1034. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasan, M.K.; El Qaidi, S.; McDonald, P.; Roy, A.; Hardwidge, P.R. Repurposing Avasimibe to Inhibit Bacterial Glycosyltransferases. Pathogens 2022, 11, 370. https://doi.org/10.3390/pathogens11030370
Hasan MK, El Qaidi S, McDonald P, Roy A, Hardwidge PR. Repurposing Avasimibe to Inhibit Bacterial Glycosyltransferases. Pathogens. 2022; 11(3):370. https://doi.org/10.3390/pathogens11030370
Chicago/Turabian StyleHasan, Md Kamrul, Samir El Qaidi, Peter McDonald, Anuradha Roy, and Philip R. Hardwidge. 2022. "Repurposing Avasimibe to Inhibit Bacterial Glycosyltransferases" Pathogens 11, no. 3: 370. https://doi.org/10.3390/pathogens11030370
APA StyleHasan, M. K., El Qaidi, S., McDonald, P., Roy, A., & Hardwidge, P. R. (2022). Repurposing Avasimibe to Inhibit Bacterial Glycosyltransferases. Pathogens, 11(3), 370. https://doi.org/10.3390/pathogens11030370