
Characterization of Salmonella enterica Contamination in Pork and Poultry Meat from São Paulo/Brazil: Serotypes, Genotypes and Antimicrobial Resistance Profiles

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Salmonella Isolation and Confirmation
2.3. Serotyping
2.4. Antimicrobial Resistance Profiling
2.5. Genotyping
2.6. Statistical Analysis
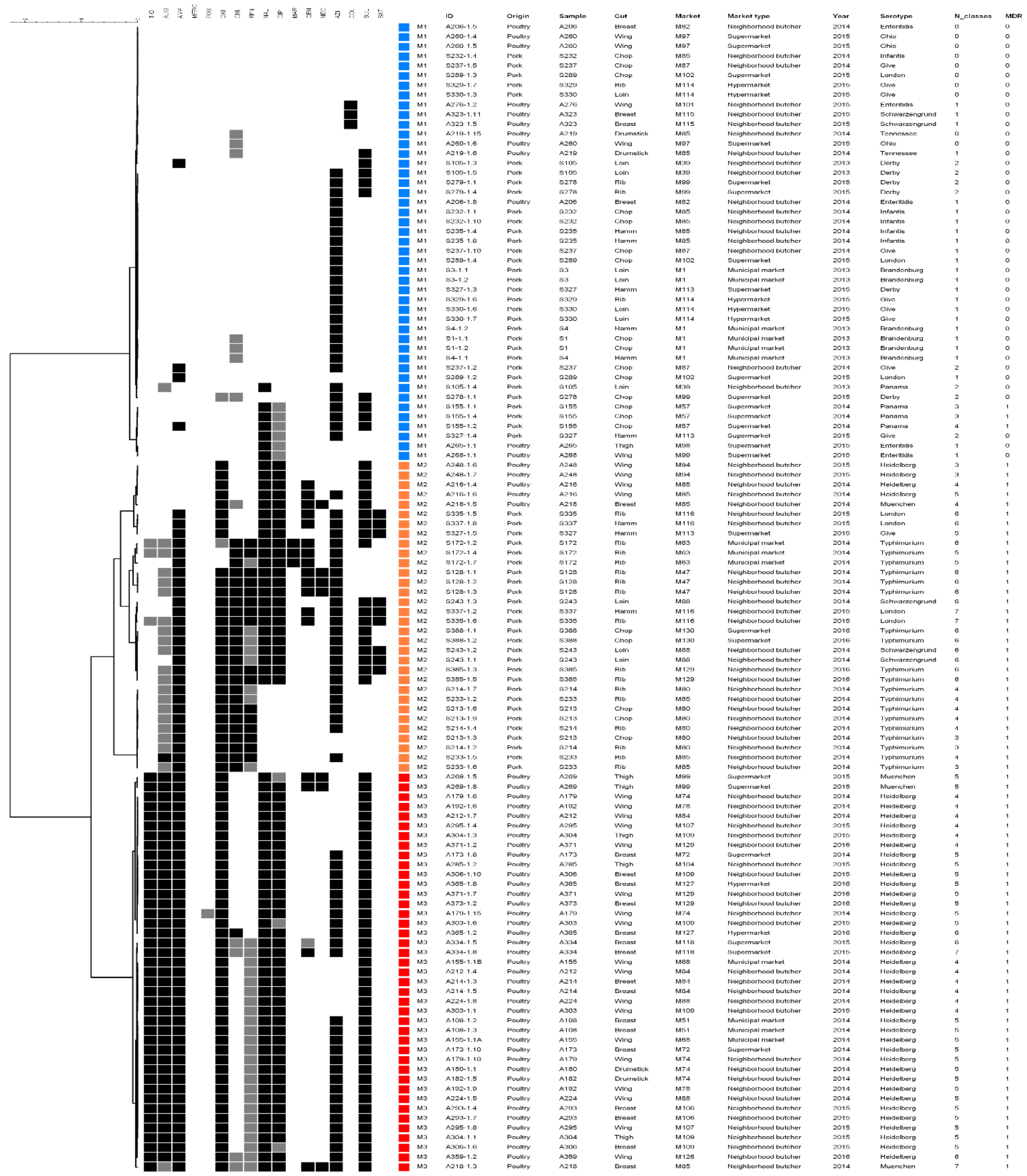
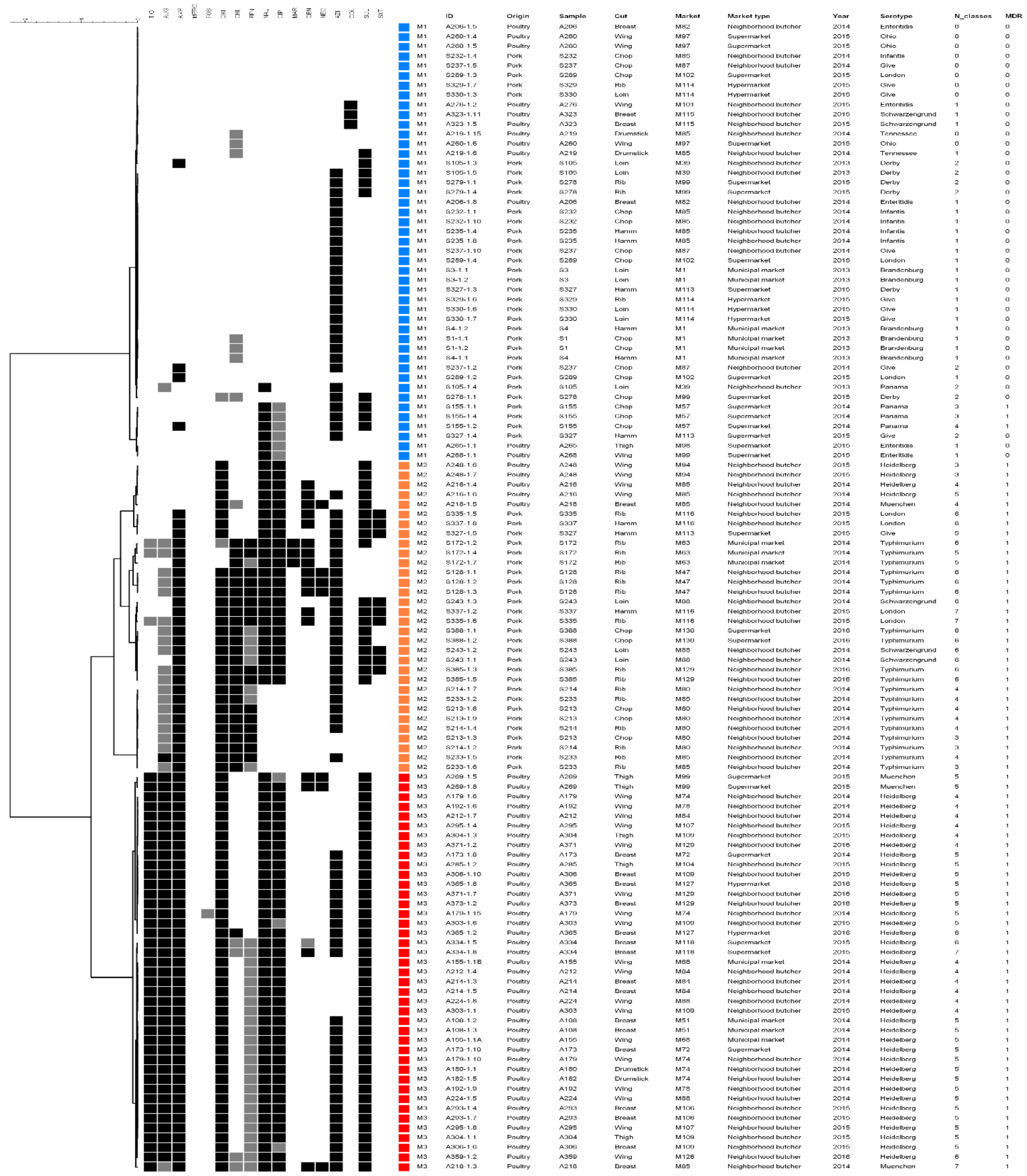
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- WHO—World Health Organization. Salmonella (Non-Typhoidal). 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) (accessed on 8 February 2022).
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. International Collaboration on Enteric Disease ‘Burden of Illness’ Studies. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.P.; Humphrey, T.J.; Maskell, D.J. Molecular insights into farm animal and zoonotic Salmonella infections. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2709–2723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC—Centers for Disease and Control Prevention. CDC Reports of Selected Salmonella Outbreak Investigations. 2020. Available online: https://www.cdc.gov/salmonella/outbreaks.html (accessed on 11 January 2022).
- CDC—Centers for Disease and Control Prevention. Advice to Clinicians. 2019. Available online: https://www.cdc.gov/salmonella/infantis-10-18/advice.html (accessed on 13 January 2022).
- EFSA—European Food Safety Authority; ECDC—European Centre for Disease Prevention and Control. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6406. [Google Scholar]
- Ministério da Agricultura, Pecuária e Abastecimento. Surtos de Doenças Transmitidas por Alimentos no Brasil. Brasil, 2018. Available online: http://portalarquivos2.saude.gov.br/images/pdf/2018/julho/02/Apresentacao-Surtos-DTA-Junho-2018.pdf (accessed on 8 February 2022).
- Campioni, F.; Cao, G.; Kastanis, G.; Janies, D.A.; Bergamini, A.M.M.; Rodrigues, D.P.; Stones, R.; Brown, E.; Allard, M.W.; Falcão, J.P. Changing of the genomic pattern of Salmonella Enteritidis strains isolated in Brazil over a 48 year-period revealed by whole genome SNP analyses. Sci. Rep. 2018, 8, 10478. [Google Scholar] [CrossRef]
- Cardoso, T.G.; Carvalho, V.M. Toxifecção alimentar por Salmonella spp. Rev. Inst. Ciênc. Saúde 2006, 24, 95–101. [Google Scholar]
- Castillo, A.C.G.; Martínez, L.H.P.; Apodaca, N.L.C. Salmonellosis and campylobacteriosis, the most prevalent zoonosis in the world. Vet. Méx. 2008, 39, 81–90. [Google Scholar]
- Perin, A.P.; Martins, B.T.F.; Barreiros, M.A.B.; Yamatogi, R.S.; Nero, L.A.; Dos Santos Bersot, L. Occurrence, quantification, pulse types, and antimicrobial susceptibility of Salmonella sp. isolated from chicken meat in the state of Paraná, Brazil. Braz. J. Microbiol. 2020, 51, 335–345. [Google Scholar] [CrossRef]
- Bersot, L.S.; Cavicchioli, V.Q.; Viana, C.; Burin, R.C.K.; Camargo, A.C.; Pinto, J.P.A.N.; Nero, L.A.; Destro, M.T. Prevalence, antimicrobial resistance, and diversity of Salmonella along the pig production chain in Southern Brazil. Pathogens 2019, 8, 204. [Google Scholar] [CrossRef] [Green Version]
- Viana, C.; Sereno, M.J.; Pegoraro, K.; Yamatogi, R.S.; Call, D.R.; Bersot, L.S.; Nero, L.A. Distribution, diversity, virulence genotypes and antibiotic resistance for Salmonella isolated from a Brazilian pork production chain. Int. J. Food Microbiol. 2019, 310, 108310. [Google Scholar] [CrossRef]
- Voss-Rech, D.; Vaz, C.S.L.; Alves, L.; Coldebella, A.; Leao, J.A.; Rodrigues, D.P.; Back, A. A temporal study of Salmonella enterica serotypes from broiler farms in Brazil. Poult. Sci. 2015, 94, 433–441. [Google Scholar] [CrossRef]
- Voss-Rech, D.; Kramer BSilva, V.S.; Rebelatto, R.; Abreu, P.G.; Coldebella, A.; Vaz, C.S.L. Longitudinal study reveals persistent environmental Salmonella Heidelberg in Brazilian broiler farms. Vet. Microbiol. 2019, 233, 118–123. [Google Scholar] [CrossRef]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A. Facultative anaerobic Gram-negative rods. In Bergey’s Manual of Determinative Bacteriology, 9th ed.; Williams & Wilkins: Baltimore, MA, USA, 1994; pp. 175–189. [Google Scholar]
- Hijazin, M.; Alber, J.; Lämmler, C.; Weitzel, T.; Hassan, A.A.; Timke, M.; Kostrzewa, M.; Prenger-Berninghoff, E.; Zschöck, M. Identification of Trueperella (Arcanobacterium) bernardiae by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry analysis and by species-specific PCR. J. Med. Microbiol. 2012, 61, 457–459. [Google Scholar] [CrossRef] [Green Version]
- Boom, R.; Sol, C.J.A.; Salimans, M.M.M.; Jansen, C.L.; Wertheim-Van Dillen, P.M.E.; Van Der Noordaa, J. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Rahn, K.; De Grandis, S.A.; Clarke, R.C.; McEwen, S.A.; Galán, J.E.; Ginocchio, C.; Curtiss, R.; Gyles, C.L. Amplification of an invA gene sequence of Salmonella Typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Mol. Cell. Probes 1992, 6, 271–279. [Google Scholar] [CrossRef]
- Popoff, M.Y.; Le Minor, L. Antigenic Formulas of the Salmonella Serovars; WHO Collaborating Centre for Reference and Research on Salmonella, World Health Organization: Geneva, Switzerland, 2001. [Google Scholar]
- CLSI—Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; CLSI supplement VET08; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- CLSI—Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Assessing the antimicrobial susceptibility of bacteria obtained from animals. Vet. Microbiol. 2010, 141, 601–604. [Google Scholar] [CrossRef]
- McLauchlin, J.; Ripabelli, G.; Brett, M.M.; Threlfall, E.J. Amplified fragment length polymorphism (AFLP) analysis of Clostridium perfringens for epidemiological typing. Int. J. Food Microbiol. 2000, 56, 21–28. [Google Scholar] [CrossRef]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Timothy, J.B. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Van Belkum, A.; Tassios, P.T.; Dijkshoorn, L.; Haeggman, S.; Cookson, B.; Fry, N.K.; Fussing, V.; Green, J.; Feil, E.; Gerner-Smidt, P.; et al. Guidelines for the validation and application of typing methods for use in bacterial epidemiology. Clin. Microbiol. Infect. Dis. 2007, 13, 1–46. [Google Scholar] [CrossRef]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef] [Green Version]
- Ristori, C.A.; Rowlands, R.E.G.; Martins, C.G.; Barbosa, M.L.; Dos Santos, L.F.; Jakabi, M.; de Melo Franco, B.D.G. Assessment of Consumer Exposure to Salmonella spp., Campylobacter spp., and Shiga Toxin-Producing Escherichia coli in Meat Products at Retail in the City of Sao Paulo, Brazil. Foodborne Pathog. Dis. 2017, 14, 447–453. [Google Scholar] [CrossRef]
- Mürmann, L.; dos Santos, M.C.; Cardoso, M. Prevalence, genetic characterization, and antimicrobial resistance of Salmonella isolated from fresh pork sausages in Porto Alegre, Brazil. Food Control 2009, 20, 191–195. [Google Scholar] [CrossRef]
- EFSA—European Food Safety Authority; ECDC—European Centre for Disease Prevention and Control. ECDC/EFSA/EMA second joint report on the integrated analysis of the consumption of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from humans and food producing animals. EFSA J. 2017, 15, 4872. [Google Scholar]
- Regalado-Pineda, I.D.; Rodarte-Medina, R.; Resendiz-Nava, C.N.; Saenz-Garcia, C.E.; Castañeda-Serrano, P.; Nava, G.M. Three-Year Longitudinal Study: Prevalence of Salmonella enterica in Chicken Meat is Higher in Supermarkets than Wet Markets from Mexico. Foods 2020, 9, 264. [Google Scholar] [CrossRef] [PubMed]
- Rortana, C.; Nguyen-Viet, H.; Tum, S.; Unger, F.; Boqvist, S.; Dang-Xuan, S.; Koam, S.; Grace, D.; Osbjer, K.; Heng, T.; et al. Prevalence of Salmonella spp. and Staphylococcus aureus in Chicken Meat and Pork from Cambodian Markets. Pathogens 2021, 10, 556. [Google Scholar] [CrossRef]
- Yang, X.; Huang, J.; Zhang, Y.; Liu, S.; Chen, L.; Xiao, C.; Zeng, H.; Wei, X.; Gu, Q.; Li, Y.; et al. Prevalence, abundance, serovars and antimicrobial resistance of Salmonella isolated from retail raw poultry meat in China. Sci. Total Environ. 2020, 713, 136385. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Zhi, W.; Qiu, Y.; Wei, L.; Tian, J.; Pan, Z.; Kang, X.; Gu, W.; Duan, L. Surveillance study of the prevalence and antimicrobial resistance of Salmonella in pork from open markets in Xuzhou, China. Food Control 2019, 98, 474–480. [Google Scholar] [CrossRef]
- Khan, A.S.; Georges, K.; Rahaman, S.; Abdela, W.; Adesiyun, A.A. Prevalence and serotypes of Salmonella spp. on chickens sold at retail outlets in Trinidad. PLoS ONE 2018, 13, e0202108. [Google Scholar] [CrossRef]
- Scheinberg, J.; Doores, S.; Cutter, C.N. A Microbiological Comparison of Poultry Products Obtained from Farmers’ Markets and Supermarkets in Pennsylvania. J. Food Saf. 2013, 33, 259–264. [Google Scholar] [CrossRef]
- de Melo, A.N.F.; Monte, D.F.M.; de Souza Pedrosa, G.T.; Balkey, M.; Jin, Q.; Brown, E.; Allard, M.; de Oliveira, T.C.R.M.; Cao, G.; Magnani, M.; et al. Genomic investigation of antimicrobial resistance determinants and virulence factors in Salmonella enterica serovars isolated from contaminated food and human stool samples in Brazil. Int. J. Food Microbiol. 2021, 343, 109091. [Google Scholar] [CrossRef]
- Kich, J.D.; Souza, A.I.A.; Montes, J.; Meneguzzi, M.; Costa, E.F.; Coldebella, A.; Corbellini, L.G.; Cardoso, M. Investigation of Listeria monocytogenes, Salmonella enterica and Yersinia enterocolitica in pig carcasses in Southern Brazil. Pesqui. Veterinária Bras. 2020, 40, 781–790. [Google Scholar] [CrossRef]
- Rodrigues, G.L.; Panzenhagen, P.; Ferrari, R.G.; Paschoalin, V.M.F.; Conte-Junior, C.A. Antimicrobial Resistance in Nontyphoidal Salmonella Isolates from Human and Swine Sources in Brazil: A Systematic Review of the Past Three Decades. Microb. Drug Resist. 2020, 26, 1260–1270. [Google Scholar] [CrossRef]
- Sjölund-Karlsson, M.; Joyce, K.; Blickenstaff, K.; Ball, T.; Haro, J.; Medalla, F.M.; Fedorka-Cray, P.; Zhao, S.; Crump, J.A.; Whichard, J.M. Antimicrobial susceptibility to azithromycin among Salmonella enterica isolates from the United States. Antimicrob. Agents Chemother. 2011, 55, 3985–3989. [Google Scholar] [CrossRef] [Green Version]
- Dutra, M.C.; Moreno, L.Z.; Dias, R.A.; Moreno, A.M. Antimicrobial Use in Brazilian Swine Herds: Assessment of Use and Reduction Examples. Microorganisms 2021, 9, 881. [Google Scholar] [CrossRef]
- Lemos, M.P.L.; Saraiva, M.M.S.; Leite, E.L.; Silva, N.M.V.; Vasconcelos, P.C.; Giachetto, P.F.; Freitas Neto, O.C.; Givisiez, P.E.N.; Gebreyes, W.A.; Oliveira, C.J.B. The post hatch prophylactic use of ceftiofur affects the cecal microbiota similar to the dietary sanguinarine supplementation in broilers. Poult. Sci. 2020, 99, 6013–6021. [Google Scholar] [CrossRef]
- Mellor, K.C.; Petrovska, L.; Thomson, N.R.; Harris, K.; Reid, S.W.J.; Mather, A.E. Antimicrobial resistance diversity suggestive of distinct Salmonella Typhimurium sources or selective pressures in food-production animals. Front. Microbiol. 2019, 10, 708. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| City Region | Nº Positive Markets | Pork | Poultry | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Chop | Rib | Loin | Hamm | Breast | Thigh | Drumstick | Wing | |||
| Center | 7/22 | 1/17 | 1/17 | 0/17 | 0/13 | 2/17 | 2/17 | 1/17 | 4/15 | 11/130 |
| East | 4/18 | 1/11 | 1/13 | 0/12 | 1/11 | 1/12 | 0/11 | 0/12 | 0/12 | 4/94 |
| North | 8/26 | 0/22 | 2/24 | 1/22 | 1/20 | 3/22 | 1/21 | 1/22 | 3/18 | 12/171 |
| West | 8/37 | 3/21 | 2/25 | 2/24 | 1/22 | 0/22 | 2/22 | 0/22 | 4/22 | 14/180 |
| South | 8/29 | 3/26 | 2/24 | 2/26 | 1/27 | 4/25 | 0/26 | 1/26 | 3/25 | 16/205 |
| Total | 35/132 | 8/97 | 8/103 | 5/101 | 4/93 | 10/98 | 5/97 | 3/99 | 14/92 | 57/780 |
| Serotype | Pork | Poultry | Total |
|---|---|---|---|
| Heidelberg | - | 42 (72.4) | 42 (35.6) |
| Typhimurium | 19 (31.7) | - | 19 (16.1) |
| Give | 10 (16.7) | - | 10 (8.5) |
| London | 7 (11.7) | - | 7 (5.9) |
| Brandenburg | 6 (10.0) | - | 6 (5.1) |
| Derby | 6 (10.0) | - | 6 (5.1) |
| Enteritidis | - | 5 (8.6) | 5 (4.2) |
| Infantis | 5 (8.3) | - | 5 (4.2) |
| Schwarzengrund | 3 (5.0) | 2 (3.4) | 5 (4.2) |
| Muenchen | - | 4 (6.9) | 4 (3.4) |
| Panama | 4 (6.7) | - | 4 (3.4) |
| Ohio | - | 3 (5.2) | 3 (2.5) |
| Tennessee | - | 2 (3.4) | 2 (1.7) |
| Total | 60 (100) | 58 (100) | 118 (100) |
| Antimicrobial | Range (µg/mL) | Pork | Poultry | ||||
|---|---|---|---|---|---|---|---|
| S N (%) | I N (%) | R * N (%) | S N (%) | I N (%) | R * N (%) | ||
| Ceftiofur | 0.25–8 | 57 (95.0) | - | 3 (5.0) | 17 (29.3) | - | 41 (70.7) |
| Amoxicillin/Clavulanate | 1/0.5–32/64 | 39 (65.0) | 20 (33.3) | 1 (1.7) | 17 (29.3) | 1 (1.7) | 40 (69.0) |
| Ampicillin | 1–64 | 29 (48.3) | - | 31 (51.7) | 17 (29.3) | - | 41 (70.7) |
| Meropenem | 0.25–8 | 60 (100) | - | - | 58 (100) | - | - |
| Fosfomycin | 8–512 | 60 (100) | - | - | 57 (98.3) | - | 1 (1.7) |
| Oxytetracycline | 2–32 | 34 (56.7) | 2 (3.3) | 24 (40.0) | 12 (20.7) | - | 46 (79.3) |
| Chloramphenicol | 4–64 | 32 (53.3) | 4 (6.7) | 24 (40.0) | 49 (84.5) | 8 (13.8) | 1 (1.7) |
| Florfenicol | 0.5–8 | 36 (60.0) | 8 (13.3) | 16 (26.7) | 34 (58.6) | - | 24 (41.4) |
| Nalidixic Acid | 8–128 | 37 (61.7) | - | 23 (38.3) | 10 (17.2) | - | 48 (82.8) |
| Ciprofloxacin | 0.06–8 | 38 (63.3) | 4 (6.7) | 18 (30.0) | 10 (17.2) | 5 (8.6) | 43 (74.1) |
| Marbofloxacin | 0.06–8 | 57 (95.0) | - | 3 (5.0) | 58 (100) | - | - |
| Gentamicin | 0.5–32 | 50 (83.3) | - | 10 (16.7) | 50 (86.2) | 1 (1.7) | 7 (12.1) |
| Neomycin | 4–16 | 57 (95.0) | - | 3 (5.0) | 54 (93.1) | - | 4 (6.9) |
| Azithromycin | 4–64 | 3 (5.0) | - | 57 (95.0) | 29 (50.0) | - | 29 (50.0) |
| Colistin | 1–16 | 60 (100) | - | - | 55 (94.8) | - | 3 (5.2) |
| Sulfamethoxazole | 256–1024 | 39 (65.0) | - | 21 (35.0) | 11 (19.0) | - | 47 (81.0) |
| Trimethoprim/Sulfamethoxazole | 2/18–4/76 | 51 (85.0) | - | 9 (15.0) | 58 (100) | - | - |
| Origin | Serotype | Nº Resistant Antimicrobial Classes | Total | ||
|---|---|---|---|---|---|
| 0–2 | 3–5 | >6 | |||
| Pork | Typhimurium | - | 11 (57.9) | 8 (42.1) | 19 (100) |
| Give | 9 (90.0) | 1 (10.0) | - | 10 (100) | |
| London | 3 (42.9) | - | 4 (57.1) | 7 (100) | |
| Brandenburg | 6 (100) | - | - | 6 (100) | |
| Derby | 6 (100) | - | - | 6 (100) | |
| Infantis | 5 (100) | - | - | 5 (100) | |
| Panama | 1 (25.0) | 3 (75.0) | - | 4 (100) | |
| Schwarzengrund | - | - | 3 (100) | 3 (100) | |
| Total | 30 (50.0) | 15 (25.0) | 15 (25.0) | 60 (100) | |
| Poultry | Heidelberg | - | 38 (90.5) | 4 (9.5) | 42 (100) |
| Enteritidis | 5 (100) | - | - | 5 (100) | |
| Muenchen | - | 3 (75.0) | 1 (25.0) | 4 (100) | |
| Ohio | 3 (100) | - | - | 3 (100) | |
| Schwarzengrund | 2 (100) | - | - | 2 (100) | |
| Tennessee | 2 (100) | - | - | 2 (100) | |
| Total | 12 (20.7) | 41 (70.7) | 5 (8.6) | 58 (100) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, V.T.M.; Moreno, L.Z.; Silva, A.P.S.; Thakur, S.; La Ragione, R.M.; Mather, A.E.; Moreno, A.M. Characterization of Salmonella enterica Contamination in Pork and Poultry Meat from São Paulo/Brazil: Serotypes, Genotypes and Antimicrobial Resistance Profiles. Pathogens 2022, 11, 358. https://doi.org/10.3390/pathogens11030358
Gomes VTM, Moreno LZ, Silva APS, Thakur S, La Ragione RM, Mather AE, Moreno AM. Characterization of Salmonella enterica Contamination in Pork and Poultry Meat from São Paulo/Brazil: Serotypes, Genotypes and Antimicrobial Resistance Profiles. Pathogens. 2022; 11(3):358. https://doi.org/10.3390/pathogens11030358
Chicago/Turabian StyleGomes, Vasco T. M., Luisa Z. Moreno, Ana Paula S. Silva, Siddhartha Thakur, Roberto M. La Ragione, Alison E. Mather, and Andrea M. Moreno. 2022. "Characterization of Salmonella enterica Contamination in Pork and Poultry Meat from São Paulo/Brazil: Serotypes, Genotypes and Antimicrobial Resistance Profiles" Pathogens 11, no. 3: 358. https://doi.org/10.3390/pathogens11030358
APA StyleGomes, V. T. M., Moreno, L. Z., Silva, A. P. S., Thakur, S., La Ragione, R. M., Mather, A. E., & Moreno, A. M. (2022). Characterization of Salmonella enterica Contamination in Pork and Poultry Meat from São Paulo/Brazil: Serotypes, Genotypes and Antimicrobial Resistance Profiles. Pathogens, 11(3), 358. https://doi.org/10.3390/pathogens11030358