
Somatic Dimorphism in Cercariae of a Bird Schistosome

 ,
,
Abstract

1. Introduction
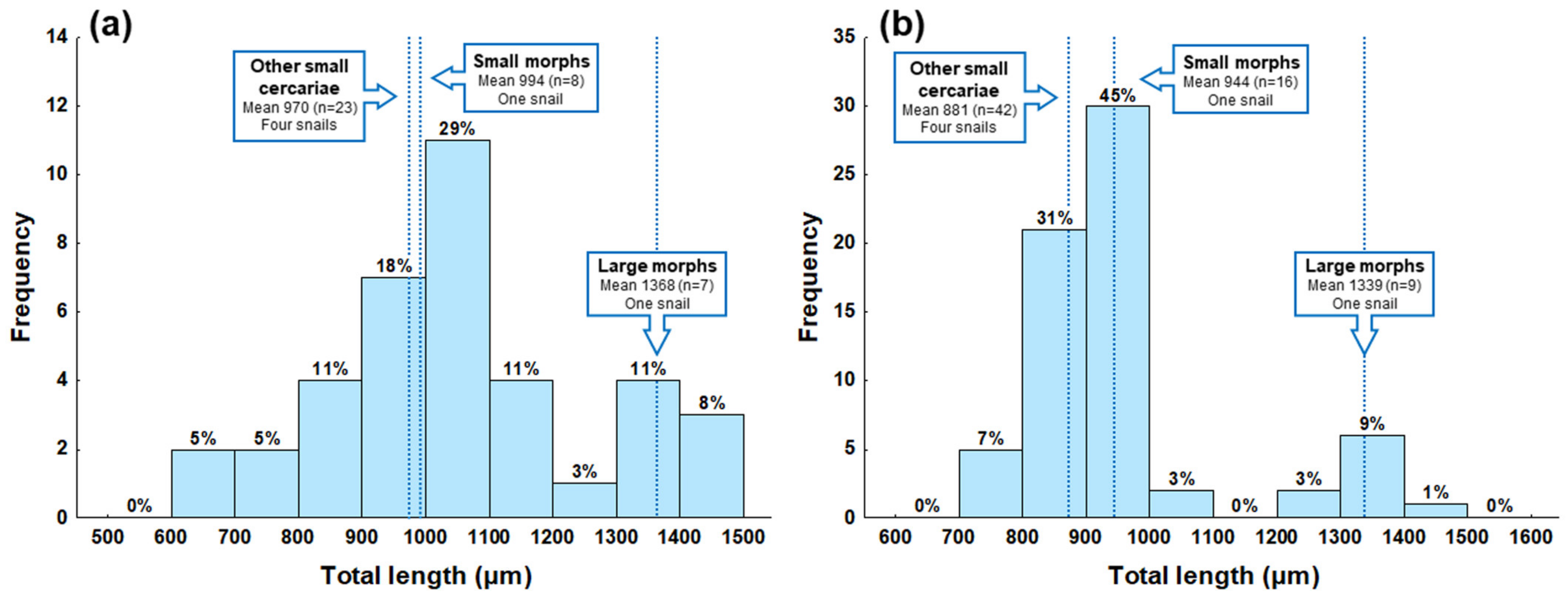
2. Results
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Species Identification
4.2. Morphometric Characterization
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group of Cercariae | Large Morphs 1 | Small Morphs 1 | Other Small Cercariae 2 | |||
|---|---|---|---|---|---|---|
| No. of Cercariae Measured | 7 | 8 | 23 | |||
| Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | Range | |
| Total length | 1368 ± 49 | 1302–1420 | 994 ± 65 | 901–1067 | 970 ± 157 | 668–1208 |
| Body length | 340 ± 42 | 304–427 | 276 ± 20 | 242–305 | 272 ± 36 | 200–332 |
| Body width | 110 ± 11 | 94–127 | 90 ± 4 | 85–97 | 93 ± 10 | 63–105 |
| Tail stem length | 702 ± 41 | 621–745 | 449 ± 39 | 401–497 | 432 ± 99 | 241–564 |
| Tail stem width | 62 ± 6 | 54–70 | 57 ± 4 | 50–62 | 52 ± 10 | 22–68 |
| Furca length | 326 ± 28 | 276–353 | 270 ± 23 | 236–292 | 265 ± 34 | 184–322 |
| Head organ length | 119 ± 9 | 105–130 | 93 ± 4 | 90–98 | 97 ± 9 | 72–108 |
| Head organ width | 76 ± 7 | 65–84 | 64 ± 2 | 61–66 | 64 ± 6 | 52–80 |
| Diameter of acetabulum | 34 ± 9 | 26–48 | 27 ± 7 | 21–39 | 34 ± 5 | 22–43 |
| Center of acetabulum to anterior body end | 212 ± 29 | 178–260 | 165 ± 17 | 141–191 | 161 ± 27 | 120–215 |
| Diameter of eye spot | 9 ± 1 | 8–10 | 8 ± 1 | 7–9 | 7 ± 1 | 6–9 |
| Eye spot to anterior body end | 145 ± 15 | 123–174 | 124 ± 13 | 111–151 | 120 ± 17 | 81–160 |
| Body length/Body width | 3.08 | 2.81–3.36 | 3.07 | 2.69–3.47 | 2.96 | 2.27–3.86 |
| Body length/Tail stem length | 0.49 | 0.43–0.60 | 0.62 | 0.53–0.76 | 0.65 | 0.49–1.01 |
| Tail stem length/Furca length | 2.17 | 1.84–2.60 | 1.67 | 1.57–1.88 | 1.62 | 0.97–1.91 |
| Group of Cercariae | Large Morphs 1 | Small Morphs 1 | Other Small Cercariae 2 | |||
|---|---|---|---|---|---|---|
| No. of Cercariae Measured | 9 | 16 | 42 | |||
| Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | Range | |
| Total length | 1339 ± 51 | 1270–1436 | 944 ± 37 | 874–1023 | 881 ± 66 | 733–991 |
| Body length | 278 ± 25 | 241–317 | 241 ± 30 | 201–332 | 223 ± 19 | 190–266 |
| Body width | 75 ± 10 | 65–87 | 66 ± 6 | 54–75 | 62 ± 9 | 45–78 |
| Tail stem length | 741 ± 39 | 680–789 | 461 ± 14 | 426–481 | 421 ± 50 | 320–499 |
| Tail stem width | 54 ± 5 | 49–64 | 47 ± 4 | 41–56 | 44 ± 5 | 34–54 |
| Furca length | 320 ± 17 | 294–340 | 242 ± 15 | 217–263 | 237 ± 22 | 169–281 |
| Head organ length | 85 ± 7 | 72–94 | 74 ± 8 | 62–91 | 73 ± 6 | 59–84 |
| Head organ width | 50 ± 6 | 42–57 | 45 ± 2 | 42–50 | 43 ± 4 | 38–55 |
| Diameter of acetabulum | 29 ± 2 | 26–33 | 26 ± 3 | 20–31 | 25 ± 3 | 20–33 |
| Center of acetabulum to anterior body end | 167 ± 29 | 105–196 | 137 ± 17 | 108–168 | 134 ± 13 | 107–160 |
| Diameter of eye spot | 9 ± 1 | 8–11 | 8 ± 1 | 7–9 | 8 ± 1 | 7–9 |
| Eye spot to anterior body end | 123 ± 14 | 104–148 | 112 ± 9 | 93–130 | 103 ± 11 | 80–126 |
| Body length/Body width | 3.76 | 2.97–4.65 | 3.70 | 2.91–5.19 | 3.71 | 2.56–5.56 |
| Body length/Tail stem length | 0.38 | 0.31–0.45 | 0.52 | 0.45–0.74 | 0.54 | 0.41–0.77 |
| Tail stem length/Furca length | 2.32 | 2.12–2.49 | 1.91 | 1.77–2.11 | 1.78 | 1.45–2.20 |
References
- Leimar, O. The evolution of phenotypic polymorphism: Randomized strategies versus evolutionary branching. Am. Nat. 2005, 165, 669–681. [Google Scholar] [CrossRef] [PubMed]
- West-Eberhard, M.J. Phenotypic Plasticity. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 2701–2707. [Google Scholar]
- Fox, R.J.; Donelson, J.M.; Schunter, C.; Ravasi, T.; Gaitán-Espitia., J.D. Beyond buying time: The role of plasticity in phenotypic adaptation to rapid environmental change. Phil. Trans. R. Soc. B 2019, 374, 20180174. [Google Scholar] [CrossRef]
- Jamie, G.A.; Meier, J.I. The persistence of polymorphisms across species radiations. Trends Ecol. Evol. 2020, 35, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Ford, E.B. Genetic Polymorphism; Faber & Faber: London, UK, 1965; p. 101. [Google Scholar]
- Fusco, G.; Minelli, A. Phenotypic plasticity in development and evolution: Facts and concepts. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Pigliucci, M.; Murren, C.J.; Schlichting, C.D. Phenotypic plasticity and evolution by genetic assimilation. J. Exp. Biol. 2006, 209, 2362–2367. [Google Scholar] [CrossRef]
- Wills, B.D.; Powell, S.; Rivera, M.D.; Suarez, A.V. Correlates and consequences of worker polymorphism in ants. Annu. Rev. Entomol. 2018, 63, 575–598. [Google Scholar] [CrossRef]
- Frances, P.; Burnie, D. Bird: The Definitive Visual Guide; Dorling Kindersley Inc.: London, UK, 2007; p. 512. [Google Scholar]
- Poulin, R. The evolution of life history strategies in parasitic animals. Adv. Parasitol. 1996, 37, 107–134. [Google Scholar] [CrossRef]
- Maizels, R.M.; Kurniawan-Atmadja, A. Variation and polymorphism in helminth parasites. Parasitology 2002, 125, 25–37. [Google Scholar] [CrossRef]
- Thompson, C.K.; Botero, A.; Wayne, A.F.; Godfrey, S.S.; Lymbery, A.J.; Thompson, R.A. Morphological polymorphism of Trypanosoma copemani and description of the genetically diverse T. vegrandis sp. nov. from the critically endangered Australian potoroid, the brush-tailed bettong (Bettongia penicillata (Gray, 1837)). Parasit. Vectors 2013, 6, 121. [Google Scholar] [CrossRef]
- Hanzelová, V.; Oros, M.; Barčák, D.; Miklisová, D.; Kirin, D.; Scholz, T. Morphological polymorphism in tapeworms: Redescription of Caryophyllaeus laticeps (Pallas, 1781) (Cestoda: Caryophyllidea) and characterisation of its morphotypes from different fish hosts. Syst. Parasitol. 2015, 90, 177–190. [Google Scholar] [CrossRef]
- Guo, Q.; Huang, M.; Liu, Y.; Zhang, X.; Gu, Z. Morphological plasticity in Myxobolus Bütschli, 1882: A taxonomic dilemma case and renaming of a parasite species of the common carp. Parasit. Vectors 2018, 11, 399. [Google Scholar] [CrossRef]
- Dobson, A.; Lafferty, K.D.; Kuris, A.M.; Hechinger, R.F.; Jetz, W. Homage to Linnaeus: How many parasites? How many hosts? Proc. Natl. Acad. Sci. USA 2008, 105, 11482–11489. [Google Scholar] [CrossRef]
- Poulin, R. Parasite biodiversity revisited: Frontiers and constraints. Int. J. Parasitol. 2014, 44, 581–589. [Google Scholar] [CrossRef]
- Pandian, T.J. Reproduction and Development in Platyhelminthes; CRC Press: Boca Raton, FL, USA, 2020; p. 320. [Google Scholar] [CrossRef]
- Galaktionov, K.V.; Dobrovolskij, A. The Biology and Evolution of Trematodes: An Essay on the Biology, Morphology, Life Cycles, Transmissions, and Evolution of Digenetic Trematodes; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; p. 592. [Google Scholar]
- Soldánová, M.; Selbach, C.; Kalbe, M.; Kostadinova, A.; Sures, B. Swimmer’s itch: Etiology, impact, and risk factors in Europe. Trends Parasitol. 2013, 29, 65–74. [Google Scholar] [CrossRef]
- Jamieson, B.G.M. Schistosoma: Biology, Pathology and Control; CRC Press: Boca Raton, FL, USA, 2017; p. 523. [Google Scholar] [CrossRef]
- Kock, S. Investigations of intermediate host specificity help to elucidate the taxonomic status of Trichobilharzia ocellata (Digenea: Schistosomatidae). Parasitology 2001, 123, 67–70. [Google Scholar] [CrossRef]
- Poulin, R. Morphological diversification in different trematode lineages: Body size, host type, or time? Parasitology 2009, 136, 85–92. [Google Scholar] [CrossRef]
- Hechinger, R.F.; Wood, A.C.; Kuris, A.M. Social organization in a flatworm: Trematode parasites form soldier and reproductive castes. Proc. R. Soc. B 2011, 278, 656–665. [Google Scholar] [CrossRef]
- Garcia-Vedrenne, A.E.; Quintana, A.C.; DeRogatis, A.M.; Martyn, K.; Kuris, A.M.; Hechinger, R.F. Social organization in parasitic flatworms—four additional echinostomoid trematodes have a soldier caste and one does not. J. Parasitol. 2016, 102, 11–20. [Google Scholar] [CrossRef]
- Poulin, R.; Kamiya, T.; Lagrue, C. Evolution, phylogenetic distribution and functional ecology of division of labour in trematodes. Parasit. Vectors 2019, 12, 1–10. [Google Scholar] [CrossRef]
- Seppälä, O.; Karvonen, A.; Valtonen, E.T. Phenotypic variation in infectivity of Diplostomum spathaceum cercariae within a population. J. Parasitol. 2007, 93, 124–1246. [Google Scholar] [CrossRef]
- Koehler, A.V.; Springer, Y.P.; Keeney, D.B.; Poulin, R. Intra-and interclonal phenotypic and genetic variability of the trematode Maritrema novaezealandensis. Biol. J. Linn. Soc. Lond. 2011, 103, 106–116. [Google Scholar] [CrossRef]
- Prokofiev, V.V.; Levakin, I.A.; Losev, E.A.; Zavirsky Ya, A.; Galaktionov, K.V. Clonal variability in expression of geo-and photoorientation in cercariae of Himasthla elongata (Trematoda: Echinostomatidae). Parazitologiia 2011, 45, 345–357. (In Russian) [Google Scholar]
- Koehler, A.V.; Poulin, R. Clone-specific immune reactions in a trematode-crustacean system. Parasitology 2012, 139, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Koehler, A.V.; Springer, Y.P.; Randhawa, H.S.; Leung, T.L.F.; Keeney, D.B.; Poulin, R. Genetic and phenotypic influences on clone-level success and host specialization in a generalist parasite. J. Evol. Biol. 2012, 25, 66–79. [Google Scholar] [CrossRef]
- Levakin, I.A.; Losev, E.A.; Nikolaev, K.E.; Galaktionov, K.V. In vitro encystment of Himasthla elongata cercariae (Digenea, Echinostomatidae) in the haemolymph of blue mussels Mytilus edulis as a tool for assessing cercarial infectivity and molluscan susceptibility. J. Helminthol. 2013, 87, 180–188. [Google Scholar] [CrossRef]
- Louhi, K.R.; Karvonen, A.; Rellstab, C.; Jokela, J. Genotypic and phenotypic variation in transmission traits of a complex life cycle parasite. Ecol. Evol. 2013, 3, 2116–2127. [Google Scholar] [CrossRef]
- Neves, R.H.; Costa-Silva, M.; Martinez, E.M.; Branquinho, T.B.; de Oliveira, R.M.; Lenzi, H.L.; Gomes, D.C.; Machado-Silva, J.R. Phenotypic plasticity in adult worms of Schistosoma mansoni (Trematoda: Schistosomatidae) evidenced by brightfield and confocal laser scanning microscopies. Mem. Inst. Oswaldo Cruz 2004, 99, 131–136. [Google Scholar] [CrossRef]
- Mati, V.L.T.; Freitas, R.M.; Bicalho, R.S.; Melo, A.L. Phenotypic plasticity of male Schistosoma mansoni from the peritoneal cavity and hepatic portal system of laboratory mice and hamsters. J. Helminthol. 2015, 89, 294–301. [Google Scholar] [CrossRef]
- Bayne, C.J.; Grevelding, C.G. Cloning of Schistosoma mansoni sporocysts in vitro and detection of genetic heterogeneity among individuals within clones. J. Parasitol. 2003, 89, 1056–1060. [Google Scholar] [CrossRef]
- Théron, A.; Sire, C.; Rognon, A.; Prugnolle, F.; Durand, P. Molecular ecology of Schistosoma mansoni transmission inferred from the genetic composition of larval and adult infrapopulations within intermediate and definitive hosts. Parasitology 2004, 129, 571–585. [Google Scholar] [CrossRef]
- Shalaby, I.; Gherbawy, Y.; Banaja, A. Genetic diversity among Schistosoma mansoni population in the western region of Saudi Arabia. Trop. Biomed. 2011, 28, 90–101. [Google Scholar]
- Korsunenko, A.; Chrisanfova, G.; Lopatkin, A.; Beer, S.A.; Voronin, M.; Ryskov, A.P.; Semyenova, S.K. Genetic differentiation of cercariae infrapopulations of the avian schistosome Trichobilharzia szidati based on RAPD markers and mitochondrial cox1 gene. Parasitol. Res. 2012, 110, 833–841. [Google Scholar] [CrossRef]
- Gu, M.J.; Li, Y.W.; Emery, A.M.; Li, S.Z.; Jiang, Y.Z.; Dong, H.F.; Zhao, Q.P. The genetic variation of different developmental stages of Schistosoma japonicum: Do the distribution in snails and pairing preference benefit the transmission? Parasit. Vectors 2020, 13, 360. [Google Scholar] [CrossRef]
- Mitta, G.; Adema, C.M.; Gourbal, B.; Loker, E.S.; Theron, A. Compatibility polymorphism in snail/schistosome interactions: From field to theory to molecular mechanisms. Dev. Comp. Immunol. 2012, 37, 1–8. [Google Scholar] [CrossRef]
- Lima, M.G.; Montresor, L.C.; Pontes, J.; Augusto, R.C.; da Silva, J.P.; Thiengo, S.C. Compatibility polymorphism based on long-term host-parasite relationships: Cross talking between Biomphalaria glabrata and the trematode Schistosoma mansoni from endemic areas in Brazil. Front. Immunol. 2019, 10, 328. [Google Scholar] [CrossRef]
- Pino, L.A.; Matinella, L.; Morales, G. The size polymorphism of the cercariae of a Venezuelan strain of Schistosoma mansoni. Rev. Soc. Bras. Med. Trop. 1999, 32, 443–446. [Google Scholar] [CrossRef]
- Jouet, D.; Skírnisson, K.; Kolářová, L.; Ferté, H. Molecular diversity of Trichobilharzia franki in two intermediate hosts (Radix auricularia and Radix peregra): A complex of species. Infect. Genet. Evol. 2010, 10, 1218–1227. [Google Scholar] [CrossRef]
- Born-Torrijos, A.; Paterson, R.A.; van Beest, G.S.; Schwelm, J.; Vyhlídalová, T.; Henriksen, E.H.; Knudsen, R.; Kristoffersen, R.; Amundsen, P.-A.; Soldánová, M. Temperature does not influence functional response of amphipods consuming different trematode prey. Parasitol. Res. 2020, 119, 4271–4276. [Google Scholar] [CrossRef]
- Born-Torrijos, A.; Paterson, R.A.; van Beest, G.S.; Vyhlídalová, T.; Henriksen, E.H.; Knudsen, R.; Kristoffersen, R.; Amundsen, P.-A.; Soldánová, M. Cercarial behaviour alters the consumer functional response of three-spined sticklebacks. J. Anim. Ecol. 2021, 90, 978–988. [Google Scholar] [CrossRef]
- Esch, G.W.; Curtis, L.A.; Barger, M.A. A perspective on the ecology of trematode communities in snails. Parasitology 2001, 123, 57–75. [Google Scholar] [CrossRef]
- Combes, C.; Bartoli, P.; Théron, A. Trematode Transmission Strategies. In The Behavioural Ecology of Parasites; Lewis, E.E., Campbell, J.F., Sukhdeo, M.V.K., Eds.; CABI: Wallingford, UK, 2002; pp. 1–12. [Google Scholar] [CrossRef]
- Morley, N.J. Cercariae (Platyhelminthes: Trematoda) as neglected components of zooplankton communities in freshwater habitats. Hydrobiologia 2012, 691, 7–19. [Google Scholar] [CrossRef]
- Pietrock, M.; Marcogliese, D.J. Free-living endohelminth stages: At the mercy of environmental conditions. Trends Parasitol. 2003, 19, 293–299. [Google Scholar] [CrossRef]
- Thieltges, D.W.; Jensen, K.T.; Poulin, R. The role of biotic factors in the transmission of free-living endohelminth stages. Parasitology 2008, 135, 407–426. [Google Scholar] [CrossRef]
- Combes, C.; Fournier, A.; Moné, H.; Théron, A. Behaviours in trematode cercariae that enhance parasite transmission: Patterns and processes. Parasitology 1994, 109, 3–13. [Google Scholar] [CrossRef]
- Haas, W. Parasitic worms: Strategies of host finding, recognition and invasion. Zoology 2003, 106, 349–364. [Google Scholar] [CrossRef]
- Horák, P.; Mikeš, L.; Lichtenbergová, L.; Skála, V.; Soldánová, M.; Brant, S.V. Avian schistosomes and outbreaks of cercarial dermatitis. Clin. Microbiol. Rev. 2015, 28, 165–190. [Google Scholar] [CrossRef]
- Soldánová, M.; Selbach, C.; Sures, B. The early worm catches the bird? Productivity and patterns of Trichobilharzia szidati cercarial emission from Lymnaea stagnalis. PLoS ONE 2016, 11, e0149678. [Google Scholar] [CrossRef]
- Ginetsinskaya, T.A. Glycogen in cercariae, and the dependence of its distribution on the specific characters of the parasite. Dokl. Akad. Nauk SSSR 1960, 135, 1012–1015. [Google Scholar]
- Ginetsinskaya, T.A. Trematodes; Their Life-Cycles, Biology and Evolution; Amerind Publ. Co., Pvt. Ltd.: New Delhi, India, 1988; p. 559. [Google Scholar]
- Morley, N.J. Cercarial swimming performance and its potential role as a key variable of trematode transmission. Parasitology 2020, 147, 1369–1374. [Google Scholar] [CrossRef]
- Lawson, R.J.; Wilson, R.A. The survival of the cercariae of Schistosoma mansoni in relation to water temperature and glycogen utilization. Parasitology 1980, 81, 337–348. [Google Scholar] [CrossRef]
- Lowenberger, C.A.; Rau, M.E. Plagiorchis elegans: Emergence, longevity and infectivity of cercariae, and host behavioural modifications during cercarial emergence. Parasitology 1994, 109, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Pechenik, J.A.; Fried, B. Effect of temperature on survival and infectivity of Echinostoma trivolvis cercariae: A test of the energy limitation hypothesis. Parasitology 1995, 111, 373–378. [Google Scholar] [CrossRef]
- Karvonen, A.; Paukku, S.; Valtonen, E.T.; Hudson, P.J. Transmission, infectivity and survival of Diplostomum spathaceum cercariae. Parasitology 2003, 127, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.J.; Thieltges, D.W. Diversity, decoys and the dilution effect: How ecological communities affect disease risk. J. Exp. Biol. 2010, 213, 961–970. [Google Scholar] [CrossRef]
- Johnson, P.T.J.; Dobson, A.; Lafferty, K.D.; Marcogliese, D.J.; Memmott, J.; Orlofske, S.A.; Poulin, R.; Thieltges, D.W. When parasites become prey: Ecological and epidemiological significance of eating parasites. Trends Ecol. Evol. 2010, 25, 362–371. [Google Scholar] [CrossRef]
- Keesing, F.; Belden, L.K.; Daszak, P.; Dobson, A.; Harvell, C.D.; Holt, R.D.; Hudson, P.; Jolles, A.; Jones, K.E.; Mitchell, C.E.; et al. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature 2010, 468, 647–652. [Google Scholar] [CrossRef]
- Goedknegt, A.; Welsh, J.; Thieltges, D.W. Parasites as Prey; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; p. 7. [Google Scholar] [CrossRef]
- Stanicka, A.; Migdalski, Ł.; Szopieray, K.; Cichy, A.; Jermacz, Ł.; Lombardo, P.; Żbikowska, E. Invaders as diluents of the cercarial dermatitis etiological agent. Pathogens 2021, 10, 740. [Google Scholar] [CrossRef]
- Kaplan, A.T.; Rebhal, S.; Lafferty, K.D.; Kuris, A.M. Small estuarine fishes feed on large trematode cercariae: Lab and field investigations. J. Parasitol. 2009, 95, 477–480. [Google Scholar] [CrossRef]
- Orlofske, S.A.; Jadin, R.C.; Johnson, P.T.J. It’s a predator-eat-parasite world: How characteristics of predator, parasite and environment affect consumption. Oecologia 2015, 178, 537–547. [Google Scholar] [CrossRef]
- Catania, S.V.; Koprivnikar, J.; McCauley, S.J. Size-dependent predation alters interactions between parasites and predators. Can. J. Zool. 2016, 94, 631–635. [Google Scholar] [CrossRef]
- Welsh, J.E.; Hempel, A.; Markovic, M.; Van der Meer, J.; Thieltges, D.W. Consumer and host body size effects on the removal of trematode cercariae by ambient communities. Parasitology 2019, 146, 342–347. [Google Scholar] [CrossRef]
- Gilbert, S.F. Ecological developmental biology: Developmental biology meets the real world. Dev. Biol. 2001, 233, 1–12. [Google Scholar] [CrossRef]
- Loker, E.S. A comparative study of the life-histories of mammalian schistosomes. Parasitology 1983, 87, 343–369. [Google Scholar] [CrossRef]
- Gérard, C.J.; Moné, H.; Théron, A. Schistosoma mansoni-Biomphalaria glabrata: Dynamics of the sporocyst population in relation to the miracidial dose and the host size. Can. J. Zool. 1993, 71, 1880–1885. [Google Scholar] [CrossRef]
- Podhorský, M.; Hůzová, Z.; Mikeš, L.; Horák, P. Cercarial dimensions and surface structures as a tool for species determination of Trichobilharzia spp. Acta Parasitol. 2009, 54, 28–36. [Google Scholar] [CrossRef]
- Poulin, R. Global warming and temperature-mediated increases in cercarial emergence in trematode parasites. Parasitology 2006, 132, 143–151. [Google Scholar] [CrossRef]
- Horák, P.; Kolářová, L.; Adema, C.M. Biology of the schistosome genus Trichobilharzia. Adv. Parasitol. 2002, 52, 155–233. [Google Scholar] [CrossRef]
- Grevelding, C.G. Genomic instability in Schistosoma mansoni. Mol. Biochem. Parasitol. 1999, 101, 207–216. [Google Scholar] [CrossRef]
- Semyenova, S.K.; Khrisanfova, G.G.; Korsunenko, A.V.; Voronin, M.V.; Beer, S.V.; Vodyanitskaya, S.V.; Serbina, E.A.; Yurlova, N.I.; Ryskov, A.P. Multilocus variation in cercariae, parthenogenetic progeny of different species of the class Trematoda. Dokl. Biol. Sci. 2007, 414, 235–238. [Google Scholar] [CrossRef]
- Galaktionov, N.K.; Podgornaya, O.I.; Strelkov, P.P.; Galaktionov, K.V. Genomic diversity of cercarial clones of Himasthla elongata (Trematoda, Echinostomatidae) determined with AFLP technique. Parasitol. Res. 2016, 115, 4587–4593. [Google Scholar] [CrossRef]
- Minchella, D.J.; Sollenberger, K.M.; Pereira de Souza, C. Distribution of schistosome genetic diversity within molluscan intermediate hosts. Parasitology 1995, 111, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.M.; Webster, J.P.; Krüger, O.; Munatsi, A.; Ndamba, J.; Woolhouse, M.E. Host–parasite population genetics: A cross-sectional comparison of Bulinus globosus and Schistosoma haematobium. Parasitology 1999, 119, 295–302. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Soldánová, M.; Georgieva, S.; Roháčová, J.; Knudsen, R.; Kuhn, J.A.; Henriksen, E.H.; Siwertsson, A.; Shaw, J.C.; Kuris, A.M.; Amundsen, P.-A.; et al. Molecular analyses reveal high species diversity of trematodes in a sub-Arctic lake. Int. J. Parasitol. 2017, 47, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Reier, S.; Haring, E.; Billinger, F.; Blatterer, H.; Duda, M.; Gorofsky, C.; Grasser, H.P.; Heinisch, W.; Hörweg, C.; Kruckenhauser, L.; et al. First confirmed record of Trichobilharzia franki Müller & Kimmig, 1994, from Radix auricularia (Linnaeus, 1758) for Austria. Parasitol. Res. 2020, 119, 4135–4141. [Google Scholar] [CrossRef]
- Helmer, N.; Blatterer, H.; Hörweg, C.; Reier, S.; Sattmann, H.; Schindelar, J.; Szucsich, N.U.; Haring, E. First record of Trichobilharzia physellae (Talbot, 1936) in Europe, a possible causative agent of cercarial dermatitis. Pathogens 2021, 10, 1473. [Google Scholar] [CrossRef]
- Brant, S.V.; Loker, E.S. Molecular systematics of the avian schistosome genus Trichobilharzia (Trematoda: Schistosomatidae) in North America. J. Parasitol. 2009, 95, 941–963. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]


| Snail Host | Radix peregra | Radix balthica | Radix balthica | Radix balthica | Radix balthica | Radix balthica | Radix balthica |
|---|---|---|---|---|---|---|---|
| Source | Jouet et al. [43] | Present study | Present study | Present study | Present study | Present study | Present study |
| Group of cercariae | – | Large morphs 1 | Small morphs 1 | Other small cercariae 2 | Large morphs 1 | Small morphs 1 | Other small cercariae 2 |
| Fixation method | Live and formalin | Live | Live | Live | Hot formalin | Hot formalin | Hot formalin |
| No. of cercariae measured | 31 | 7 | 8 | 23 | 9 | 16 | 42 |
| Total length | 864 | 1368 | 994 | 970 | 1339 | 944 | 881 |
| Body length | 257 | 340 | 276 | 272 | 278 | 241 | 223 |
| Body width | 73 | 110 | 90 | 93 | 75 | 66 | 62 |
| Tail stem length | 379 | 702 | 449 | 432 | 741 | 461 | 421 |
| Tail stem width | 50 | 62 | 57 | 52 | 54 | 47 | 44 |
| Furca length | 227 | 326 | 270 | 265 | 320 | 242 | 237 |
| Head organ length | – | 119 | 93 | 97 | 85 | 74 | 73 |
| Head organ width | – | 76 | 64 | 64 | 50 | 45 | 43 |
| Diameter of acetabulum | 28 | 34 | 27 | 34 | 29 | 26 | 25 |
| Center of acetabulum to anterior body end | 182 | 212 | 165 | 161 | 167 | 137 | 134 |
| Diameter of eye spot | – | 9 | 8 | 7 | 9 | 8 | 8 |
| Eye spot to anterior body end | 134 | 145 | 124 | 120 | 123 | 112 | 103 |
| Body length/Body width | 3.52 3 | 3.08 | 3.07 | 2.96 | 3.76 | 3.70 | 3.71 |
| Body length/Tail stem length | 0.68 3 | 0.49 | 0.62 | 0.65 | 0.38 | 0.52 | 0.54 |
| Tail stem length/Furca length | 1.69 3 | 2.17 | 1.67 | 1.62 | 2.32 | 1.91 | 1.78 |
| Data Set | Live Cercariae | Formalin-Fixed Cercariae | ||||||
|---|---|---|---|---|---|---|---|---|
| Parameter Tested | Df 1 | MS 2 | F 3 | p 4 | Df | MS 2 | F 3 | p 4 |
| Total length | 2 | 0.348 | 17.84 | <0.001 | 2 | 0.656 | 153.1 | <0.001 |
| Body length | 2 | 0.138 | 9.13 | <0.001 | 2 | 0.189 | 21.3 | <0.001 |
| Body width | 2 | 0.095 | 9.18 | <0.001 | 2 | 0.147 | 8.49 | <0.001 |
| Tail stem length | 2 | 0.713 | 16.11 | <0.001 | 2 | 1.210 | 115.7 | <0.001 |
| Tail stem width | 2 | 0.106 | 3.14 | 0.056 | 2 | 0.129 | 13.30 | <0.001 |
| Furca length | 2 | 0.123 | 8.63 | <0.001 | 2 | 0.343 | 45.6 | <0.001 |
| Head organ length | 2 | 0.137 | 18.17 | <0.001 | 2 | 0.093 | 11.5 | <0.001 |
| Head organ width | 2 | 0.086 | 12.27 | <0.001 | 2 | 0.074 | 10.94 | <0.001 |
| Diameter of acetabulum | 2 | 0.153 | 4.36 | <0.05 | 2 | 0.075 | 6.17 | <0.01 |
| Center of acetabulum to anterior body end | 2 | 0.216 | 9.52 | <0.001 | 2 | 0.160 | 11.31 | <0.001 |
| Diameter of eye spot | 2 | 0.093 | 13.55 | <0.001 | 2 | 0.086 | 14.33 | <0.001 |
| Eye spot to anterior body end | 2 | 0.101 | 5.99 | <0.01 | 2 | 0.137 | 12.93 | <0.001 |
| Body length/Body width | 2 | 0.005 | 0.57 | 0.570 | 2 | 0.001 | 0.06 | 0.94 |
| Body length/Tail stem length | 2 | 0.030 | 6.68 | <0.01 | 2 | 0.046 | 17.39 | <0.001 |
| Tail stem length/Furca length | 2 | 0.101 | 13.08 | <0.001 | 2 | 0.121 | 32.25 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soldánová, M.; Kundid, P.; Scholz, T.; Kristoffersen, R.; Knudsen, R. Somatic Dimorphism in Cercariae of a Bird Schistosome. Pathogens 2022, 11, 290. https://doi.org/10.3390/pathogens11030290
Soldánová M, Kundid P, Scholz T, Kristoffersen R, Knudsen R. Somatic Dimorphism in Cercariae of a Bird Schistosome. Pathogens. 2022; 11(3):290. https://doi.org/10.3390/pathogens11030290
Chicago/Turabian StyleSoldánová, Miroslava, Petra Kundid, Tomáš Scholz, Roar Kristoffersen, and Rune Knudsen. 2022. "Somatic Dimorphism in Cercariae of a Bird Schistosome" Pathogens 11, no. 3: 290. https://doi.org/10.3390/pathogens11030290
APA StyleSoldánová, M., Kundid, P., Scholz, T., Kristoffersen, R., & Knudsen, R. (2022). Somatic Dimorphism in Cercariae of a Bird Schistosome. Pathogens, 11(3), 290. https://doi.org/10.3390/pathogens11030290