
Molecular Detection and Phylogenetic Analyses of Diverse Bartonella Species in Bat Ectoparasites Collected from Yunnan Province, China

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Bat Ectoparasites Sampling, Mixing, and Species Identification
2.2. Bartonella Detection
2.3. Molecular Biological Characteristics of Newly Discovered Bartonella
2.3.1. Identification of Bartonella
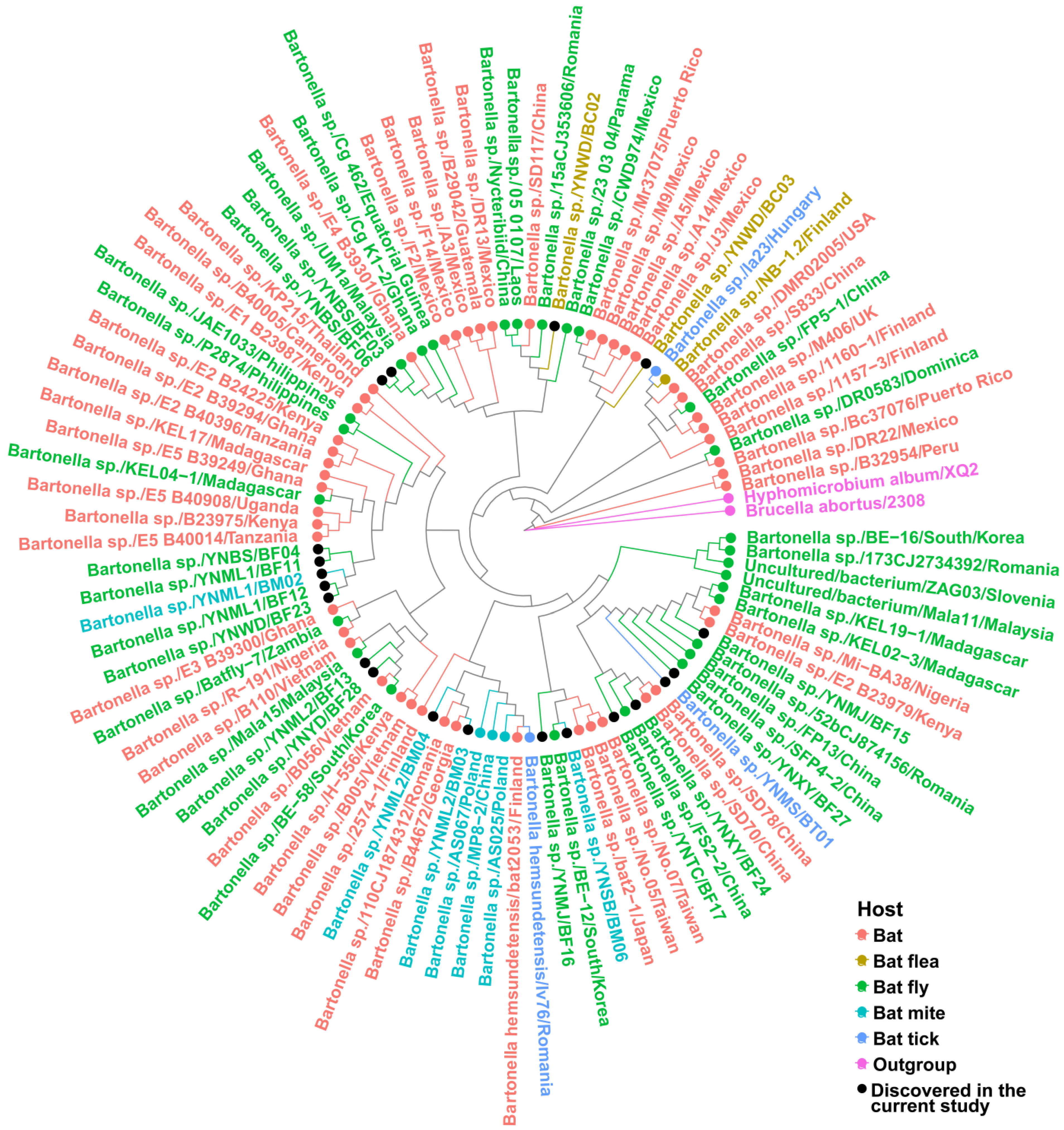
2.3.2. Phylogenetic Analyses
3. Discussion
4. Material and Methods
4.1. Ethics Statement
4.2. Sample Collection
4.3. Sample Mixing, Deoxyribonucleic Acid Extraction, and Host Species Identification
4.4. Bartonella Detection and Identification
4.5. Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dvm, E.B.B.; Maggi, R.; Chomel, B.B.; Lappin, M.R. Bartonellosis: An emerging infectious disease of zoonotic importance to animals and human beings. J. Veter. Emerg. Crit. Care 2010, 20, 8–30. [Google Scholar] [CrossRef]
- Harms, A.; Dehio, C. Intruders below the Radar: Molecular Pathogenesis of Bartonella spp. Clin. Microbiol. Rev. 2012, 25, 42–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okaro, U.; Addisu, A.; Casanas, B.; Anderson, B. Bartonella Species, an Emerging Cause of Blood-Culture-Negative Endocarditis. Clin. Microbiol. Rev. 2017, 30, 709–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, H.; Li, S.; Lu, L.; Wang, R.; Song, X.; Sun, K.; Shi, Y.; Li, D.; Yu, J. Genetic diversity of Bartonella species in small mammals in the Qaidam Basin, western China. Sci. Rep. 2021, 11, 1735. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.Y.; Tsigrelis, C.; Baddour, L.M.; Lepidi, H.; Rolain, J.-M.; Patel, R.; Raoult, D. Candidatus Bartonella mayotimonensis and Endocarditis. Emerg. Infect. Dis. 2010, 16, 500–503. [Google Scholar] [CrossRef]
- Kosoy, M.; McKee, C.; Albayrak, L.; Fofanov, Y. Genotyping of Bartonella bacteria and their animal hosts: Current status and perspectives. Parasitology 2018, 145, 543–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important Reservoir Hosts of Emerging Viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Baker, M.L.; Kulcsar, K.; Misra, V.; Plowright, R.; Mossman, K. Novel Insights into Immune Systems of Bats. Front. Immunol. 2020, 11, 26. [Google Scholar] [CrossRef] [Green Version]
- Dacheux, L.; Cervantes-Gonzalez, M.; Guigon, G.; Thiberge, J.-M.; Vandenbogaert, M.; Maufrais, C.; Caro, V.; Bourhy, H. A Preliminary Study of Viral Metagenomics of French Bat Species in Contact with Humans: Identification of New Mammalian Viruses. PLoS ONE 2014, 9, e87194. [Google Scholar] [CrossRef]
- Kading, R.C.; Schountz, T. Flavivirus Infections of Bats: Potential Role in Zika Virus Ecology. Am. J. Trop. Med. Hyg. 2016, 95, 993–996. [Google Scholar] [CrossRef]
- Afelt, A.; Lacroix, A.; Zawadzka-Pawlewska, U.; Pokojski, W.; Buchy, P.; Frutos, R. Distribution of bat-borne viruses and environment patterns. Infect. Genet. Evol. 2017, 58, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Mühldorfer, K.; Speck, S.; Wibbelt, G. Diseases in free-ranging bats from Germany. BMC Veter. Res. 2011, 7, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Urushadze, L.; Osikowicz, L.; McKee, C.; Kuzmin, I.; Kandaurov, A.; Babuadze, G.; Natradze, I.; Imnadze, P.; Kosoy, M. Molecular Survey of Bacterial Zoonotic Agents in Bats from the Country of Georgia (Caucasus). PLoS ONE 2017, 12, e0171175. [Google Scholar] [CrossRef] [PubMed]
- Corduneanu, A.; Sándor, A.D.; Ionică, A.M.; Hornok, S.; Leitner, N.; Bagó, Z.; Stefke, K.; Fuehrer, H.-P.; Mihalca, A.D. Bartonella DNA in heart tissues of bats in central and eastern Europe and a review of phylogenetic relations of bat-associated bartonellae. Parasites Vectors 2018, 11, 489. [Google Scholar] [CrossRef]
- Nabeshima, K.; Sato, S.; Kabeya, H.; Kato, C.; Suzuki, K.; Maruyama, S. Isolation and genetic properties of Bartonella in eastern bent-wing bats (Miniopterus fuliginosus) in Japan. Infect. Genet. Evol. 2020, 83, 104354. [Google Scholar] [CrossRef]
- Han, H.; Li, Z.; Li, X.; Liu, J.; Peng, Q.; Wang, R.; Gu, X.; Jiang, Y.; Zhou, C.; Li, D.; et al. Bats and their ectoparasites (Nycteribiidae and Spinturnicidae) carry diverse novel Bartonella genotypes, China. Transbound. Emerg. Dis. 2022, 69, e845–e858. [Google Scholar] [CrossRef]
- Poofery, J.; Narapakdeesakul, D.; Riana, E.; Arnuphapprasert, A.; Nugraheni, Y.R.; Ngamprasertwong, T.; Wangthongchaicharoen, M.; Soisook, P.; Bhodhibundit, P.; Kaewthamasorn, M. Molecular identification and genetic diversity of Bartonella spp. in 24 bat species from Thailand. Transbound. Emerg. Dis. 2022, 69, e717–e733. [Google Scholar] [CrossRef]
- Bai, Y.; Osinubi, M.O.V.; Osikowicz, L.; McKee, C.; Vora, N.M.; Rizzo, M.R.; Recuenco, S.; Davis, L.; Niezgoda, M.; Ehimiyein, A.M.; et al. Human Exposure to Novel Bartonella Species from Contact with Fruit Bats. Emerg. Infect. Dis. 2018, 24, 2317–2323. [Google Scholar] [CrossRef] [Green Version]
- McKee, C.D.; Bai, Y.; Webb, C.T.; Kosoy, M.Y. Bats are key hosts in the radiation of mammal-associated Bartonella bacteria. Infect. Genet. Evol. 2021, 89, 104719. [Google Scholar] [CrossRef]
- Breitschwerdt, E.B.; Kordick, D.L. Bartonella infection in animals: Carriership, reservoir potential, pathogenicity, and zoonotic potential for human infection. Clin. Microbiol. Rev. 2000, 13, 428–438. [Google Scholar] [CrossRef]
- Billeter, S.A.; Levy, M.G.; Chomel, B.B.; Breitschwerdt, E.B. Vector transmission of Bartonella species with emphasis on the potential for tick transmission. Med. Veter. Èntomol. 2008, 22, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Reis, C.; Cote, M.; Le Rhun, D.; Lecuelle, B.; Levin, M.L.; Vayssier-Taussat, M.; Bonnet, S.I. Vector Competence of the Tick Ixodes ricinus for Transmission of Bartonella birtlesii. PLOS Negl. Trop. Dis. 2011, 5, e1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wechtaisong, W.; Bonnet, S.I.; Lien, Y.-Y.; Chuang, S.-T.; Tsai, Y.-L. Transmission of Bartonella henselae within Rhipicephalus sanguineus: Data on the Potential Vector Role of the Tick. PLOS Negl. Trop. Dis. 2020, 14, e0008664. [Google Scholar] [CrossRef]
- Szentiványi, T.; Christe, P.; Glaizot, O. Bat Flies and Their Microparasites: Current Knowledge and Distribution. Front. Veter. Sci. 2019, 6, 115. [Google Scholar] [CrossRef]
- Xu, Z.; Feng, Y.; Chen, X.; Shi, M.; Fu, S.; Yang, W.; Liu, W.J.; Gao, G.F.; Liang, G. Virome of Bat-Infesting Arthropods: Highly Divergent Viruses in Different Vectors. J. Virol. 2022, 96, e01464-21. [Google Scholar] [CrossRef]
- Stuckey, M.J.; Chomel, B.B.; de Fleurieu, E.C.; Aguilar-Setién, A.; Boulouis, H.J.; Chang, C.C. Bartonella, bats and bugs: A review. Comp. Immunol. Microbiol. Infect. Dis. 2017, 55, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Morse, S.F.; Olival, K.J.; Kosoy, M.; Billeter, S.; Patterson, B.D.; Dick, C.W.; Dittmar, K. Global distribution and genetic diversity of Bartonella in bat flies (Hippoboscoidea, Streblidae, Nycteribiidae). Infect. Genet. Evol. 2012, 12, 1717–1723. [Google Scholar] [CrossRef]
- Reeves, W.K.; Loftis, A.D.; Gore, J.A.; Dasch, G.A. Molecular evidence for novel Bartonella species in Trichobius major (Diptera: Streblidae) and Cimex adjunctus (Hemiptera: Cimicidae) from two southeastern bat caves, U.S.A. J. Vector Ecol. 2005, 30, 339–341. [Google Scholar]
- Billeter, S.A.; Hayman, D.T.; Peel, A.J.; Baker, K.; Wood, J.L.; Cunningham, A.; Suu-Ire, R.; Dittmar, K.; Kosoy, M.Y. Bartonella species in bat flies (Diptera: Nycteribiidae) from western Africa. Parasitology 2012, 139, 324–329. [Google Scholar] [CrossRef]
- Brook, C.E.; Bai, Y.; Dobson, A.P.; Osikowicz, L.M.; Ranaivoson, H.C.; Zhu, Q.; Kosoy, M.Y.; Dittmar, K. Bartonella spp. in Fruit Bats and Blood-Feeding Ectoparasites in Madagascar. PLOS Negl. Trop. Dis. 2015, 9, e0003532. [Google Scholar] [CrossRef]
- Qiu, Y.; Kajihara, M.; Nakao, R.; Mulenga, E.; Harima, H.; Hang’Ombe, B.M.; Eto, Y.; Changula, K.; Mwizabi, D.; Sawa, H.; et al. Isolation of Candidatus Bartonella rousetti and Other Bat-associated Bartonellae from Bats and Their Flies in Zambia. Pathogens 2020, 9, 469. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Seo, M.-G.; Lee, S.-H.; Oem, J.-K.; Kim, S.-H.; Jeong, H.; Kim, Y.; Jheong, W.-H.; Kwon, O.-D.; Kwak, D. Relationship among bats, parasitic bat flies, and associated pathogens in Korea. Parasites Vectors 2021, 14, 503. [Google Scholar] [CrossRef] [PubMed]
- Low, V.L.; Tan, T.K.; Tohiran, K.A.; Lim, Y.A.L.; AbuBakar, S.; Nasir, D.M. A novel clade of bat-associated Bartonella detected in the bat fly Leptocyclopodia ferrari (Diptera: Nycteribiidae) parasitizing Cynopterus brachyotis (Chiroptera: Pteropodidae). Veter. Microbiol. 2022, 264, 109284. [Google Scholar] [CrossRef] [PubMed]
- Szubert-Kruszyńska, A.; Stańczak, J.; Cieniuch, S.; Podsiadły, E.; Postawa, T.; Michalik, J. Bartonella and Rickettsia Infections in Haematophagous Spinturnix myoti Mites (Acari: Mesostigmata) and their Bat Host, Myotis myotis (Yangochiroptera: Vespertilionidae), from Poland. Microb. Ecol. 2019, 77, 759–768. [Google Scholar] [CrossRef]
- Tahir, D.; Socolovschi, C.; Marié, J.-L.; Ganay, G.; Berenger, J.-M.; Bompar, J.-M.; Blanchet, D.; Cheuret, M.; Mediannikov, O.; Raoult, D.; et al. New Rickettsia species in soft ticks Ornithodoros hasei collected from bats in French Guiana. Ticks Tick-Borne Dis. 2016, 7, 1089–1096. [Google Scholar] [CrossRef]
- Hornok, S.; Szőke, K.; Meli, M.L.; Sándor, A.D.; Görföl, T.; Estók, P.; Wang, Y.; Tu, V.T.; Kováts, D.; Boldogh, S.A.; et al. Molecular detection of vector-borne bacteria in bat ticks (Acari: Ixodidae, Argasidae) from eight countries of the Old and New Worlds. Parasites Vectors 2019, 12, 50. [Google Scholar] [CrossRef] [Green Version]
- Reeves, W.K.; Rogers, T.E.; Durden, L.A.; Dasch, G. Association of Bartonella with the fleas (Siphonaptera) of rodents and bats using molecular techniques. J. Vector Ecol. 2007, 32, 118–122. [Google Scholar] [CrossRef]
- Veikkolainen, V.; Vesterinen, E.J.; Lilley, T.M.; Pulliainen, A.T. Bats as Reservoir Hosts of Human Bacterial Pathogen, Bartonella mayotimonensis. Emerg. Infect. Dis. 2014, 20, 960–967. [Google Scholar] [CrossRef]
- Bush, S.E.; Robbins, R.G. New host and locality records for Ixodes simplex Neumann and Ixodes vespertilionis Koch (Acari: Ixodidae) from bats (Chiroptera: Hipposideridae, Rhinolophidae and Vespertilionidae) in southern China. Int. J. Acarol. 2012, 38, 1–5. [Google Scholar] [CrossRef]
- Hornok, S.; Görföl, T.; Estók, P.; Tu, V.T.; Kontschán, J. Description of a new tick species, Ixodes collaris n. sp. (Acari: Ixodidae), from bats (Chiroptera: Hipposideridae, Rhinolophidae) in Vietnam. Parasites Vectors 2016, 9, 332. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.-W.; Hsu, Y.-M.; Chomel, B.B.; Lin, L.-K.; Pei, J.-C.; Wu, S.-H.; Chang, C.-C. Identification of novel Bartonella spp. in bats and evidence of Asian gray shrew as a new potential reservoir of Bartonella. Veter. Microbiol. 2012, 156, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Han, H.-J.; Wen, H.-L.; Zhao, L.; Liu, J.-W.; Luo, L.-M.; Zhou, C.-M.; Qin, X.-R.; Zhu, Y.-L.; Zheng, X.-X.; Yu, X.-J. Novel Bartonella Species in Insectivorous Bats, Northern China. PLoS ONE 2017, 12, e0167915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birtles, R.; Raoult, D. Comparison of Partial Citrate Synthase Gene (gltA) Sequences for Phylogenetic Analysis of Bartonella Species. Int. J. Syst. Bacteriol. 1996, 46, 891–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Scola, B.; Zeaiter, Z.; Khamis, A.; Raoult, D. Gene-sequence-based criteria for species definition in bacteriology: The Bartonella paradigm. Trends Microbiol. 2003, 11, 318–321. [Google Scholar] [CrossRef]
- Vayssier-Taussat, M.; Le Rhun, D.; Bonnet, S.; Cotté, V. Insights in Bartonella Host Specificity. Ann. N. Y. Acad. Sci. 2009, 1166, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Lei, B.R.; Olival, K.J. Contrasting Patterns in Mammal–Bacteria Coevolution: Bartonella and Leptospira in Bats and Rodents. PLOS Negl. Trop. Dis. 2014, 8, e2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dick, C.W. High host specificity of obligate ectoparasites. Ecol. Èntomol. 2007, 32, 446–450. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]
- Bininda-Emonds, O.R.P.; Cardillo, M.; Jones, K.E.; MacPhee, R.D.E.; Beck, R.M.D.; Grenyer, R.; Price, S.A.; Vos, R.A.; Gittleman, J.L.; Purvis, A. The delayed rise of present-day mammals. Nature 2007, 446, 507–512. [Google Scholar] [CrossRef]
- Feng, Y.; Li, Y.; Fu, S.; Li, X.; Song, J.; Zhang, H.; Yang, W.; Zhang, Y.; Pan, H.; Liang, G. Isolation of Kaeng Khoi virus (KKV) from Eucampsipoda sundaica bat flies in China. Virus Res. 2017, 238, 94–100. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Norman, A.F.; Regnery, R.; Jameson, P.; Greene, C.; Krause, D.C. Differentiation of Bartonella-like isolates at the species level by PCR-restriction fragment length polymorphism in the citrate synthase gene. J. Clin. Microbiol. 1995, 33, 1797–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Hayman, D.T.S.; McKee, C.; Kosoy, M.Y. Classification of Bartonella Strains Associated with Straw-Colored Fruit Bats (Eidolon helvum) across Africa Using a Multi-locus Sequence Typing Platform. PLOS Negl. Trop. Dis. 2015, 9, e0003478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, J.G.; Rozental, T.; Guterres, A.; Teixeira, B.R.; Andrade-Silva, B.E.; da Costa-Neto, S.F.; Furtado, M.C.; Moratelli, R.; D’Andrea, P.S.; Lemos, E.R.S. Investigation of Bartonella spp. in brazilian mammals with emphasis on rodents and bats from the Atlantic Forest. Int. J. Parasitol. Parasites Wildl. 2020, 13, 80–89. [Google Scholar] [CrossRef]
- Roux, V.; Raoult, D. The 16S-23S rRNA intergenic spacer region of Bartonella (Rochalimaea) species is longer than usually described in other bacteria. Gene 1995, 156, 107–111. [Google Scholar] [CrossRef]
- Paziewska, A.; Harris, P.D.; Zwolińska, L.; Bajer, A.; Siński, E. Recombination Within and Between Species of the Alpha Proteobacterium Bartonella Infecting Rodents. Microb. Ecol. 2010, 61, 134–145. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ectoparasite Family | Location | Number of Ectoparasites | Number of Mixed Pools | Bartonella PCR | |||
|---|---|---|---|---|---|---|---|
| gltA | ftsZ | rpoB | ITS | ||||
| Ischnopsyllidae | Baoshan | 1 | 1 | 0 | 0 | 0 | 0 |
| (Bat flea) | Wanding | 20 | 2 | 2 | 2 | 2 | 1 |
| Ixodidae | Mangshi | 1 | 1 | 1 | 1 | 1 | 1 |
| (Bat tick) | Mojiang | 2 | 1 | 0 | 0 | 0 | 0 |
| Tengchong | 2 | 1 | 0 | 0 | 0 | 0 | |
| Spinturnicidae | Mengla | 33 | 2 | 1 | 1 | 1 | 1 |
| (Bat mite) | Menglian | 55 | 2 | 2 | 2 | 2 | 1 |
| Mojiang | 15 | 1 | 0 | 0 | 0 | 0 | |
| Shuangbai | 10 | 1 | 1 | 1 | 0 | 1 | |
| Streblidae | Baoshan | 1 | 1 | 0 | 0 | 0 | 0 |
| (Bat fly) | Mojiang | 8 | 1 | 0 | 0 | 0 | 0 |
| Wanding | 2 | 1 | 0 | 0 | 0 | 0 | |
| Nycteribiidae | Baoshan | 79 | 6 | 3 | 3 | 3 | 3 |
| (Wingless bat fly) | Mangshi | 19 | 3 | 0 | 0 | 0 | 0 |
| Mengla | 43 | 2 | 2 | 2 | 2 | 2 | |
| Menglian | 4 | 1 | 1 | 1 | 1 | 1 | |
| Mojiang | 26 | 2 | 2 | 2 | 1 | 2 | |
| Tengchong | 15 | 2 | 1 | 1 | 0 | 1 | |
| Wanding | 44 | 4 | 1 | 1 | 1 | 1 | |
| Xiangyun | 44 | 4 | 2 | 2 | 2 | 2 | |
| Yongde | 10 | 1 | 1 | 1 | 1 | 1 | |
| NO. | Pools | Classification | Strains | Host Species | BLASTn Hits on Known Bartonella (BLAST Nucleotide Identity) |
|---|---|---|---|---|---|
| 1 | YNML1/BM02 | Bartonella sp. | YNML1/BM02 | Spinturnicidae sp. | KM030506/Bartonella sp./B23975 (93.42%) |
| 2 | YNML2/BM03 | Bartonella sp. | YNML2/BM03 | Spinturnicidae sp. | MK140370/Bartonella sp./B44672 (100%) |
| 3 | YNML2/BM04 | Bartonella sp. | YNML2/BM04 | Spinturnicidae sp. | MK140216/Bartonella sp./110CJ1874312 (100%) |
| 4 | YNSB/BM06 | Bartonella sp. | YNSB/BM06 | Spinturnicidae sp. | MT362931/Bartonella sp./BE-12 (99.47%) |
| 5 | YNMS/BT01 | Bartonella sp. | YNMS/BT01 | Ixodes vespertilionis | KX655829/Bartonella sp./SD-70/2015 (98.42%) |
| 6 | YNWD/BC02 | Bartonella sp. | YNWD/BC02 | Thaumapsylla sp. | KM215691/Bartonella chomelii/Ru55 (88.42%) |
| 7 | YNWD/BC03 | Bartonella sp. | YNWD/BC03 | Thaumapsylla sp. | FJ589054/Bartonella sp./RT230YN (94.74%) |
| 8 | YNBS/BF03 | Bartonella sp. | YNBS/BF03 | Eucampsipoda africana | MZ388461/Bartonella sp./UM1a (95.26%) |
| 9 | YNBS/BF04 | Bartonella sp. | YNBS/BF04 | Eucampsipoda africana | KM030506/Bartonella sp./B23975 (93.42%) |
| 10 | YNBS/BF06 | Bartonella sp. | YNBS/BF06 | Eucampsipoda africana | MZ388461/Bartonella sp./UM1a (94.99%) |
| 11 | YNML1/BF11 | Bartonella sp. | YNML1/BF11 | Eucampsipoda sundaica | KM030526/Bartonella sp./B40908 (93.40%) |
| 12 | YNML1/BF12 | Bartonella sp. | YNML1/BF12 | Eucampsipoda sundaica | KP010193/Bartonella sp./KEL17 (93.16%) |
| 13 | YNML2/BF13 | Bartonella sp. | YNML2/BF13 | Phthiridium sp. | KP100360/Bartonella sp./B110 (95.26%) |
| 14 | YNMJ/BF15 | Bartonella sp. | YNMJ/BF15 | Nycteribiidae sp. | KM030503/Bartonella sp./B23797 (97.89%) |
| 15 | YNMJ/BF16 | Bartonella sp. | YNMJ/BF16 | Nycteribiidae sp. | MT362931/Bartonella sp./BE-12 (98.94%) |
| 16 | YNTC/BF17 | Bartonella sp. | YNTC/BF17 | Nycteribia sp. | KX655829/Bartonella sp./SD-70/2015 (98.68%) |
| 17 | YNWD/BF23 | Bartonella sp. | YNWD/BF23 | Eucampsipoda africana | KM030506/Bartonella sp./B23975 (93.42%) |
| 18 | YNXY/BF24 | Bartonella sp. | YNXY/BF24 | Penicillidia monoceros | KX655829/Bartonella sp./SD-70/2015 (99.47%) |
| 19 | YNXY/BF27 | Bartonella sp. | YNXY/BF27 | Nycteribia sp. | KX655829/Bartonella sp./SD-70/2015 (94.99%) |
| 20 | YNYD/BF28 | Bartonella sp. | YNYD/BF28 | Phthiridium sp. | KP100348/Bartonella sp./B056 (95.79%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuang, G.; Zhang, J.; Yang, W.; Pan, H.; Han, X.; Yang, L.; Wang, J.; Yang, T.; Song, Z.; Feng, Y.; et al. Molecular Detection and Phylogenetic Analyses of Diverse Bartonella Species in Bat Ectoparasites Collected from Yunnan Province, China. Pathogens 2022, 11, 1283. https://doi.org/10.3390/pathogens11111283
Kuang G, Zhang J, Yang W, Pan H, Han X, Yang L, Wang J, Yang T, Song Z, Feng Y, et al. Molecular Detection and Phylogenetic Analyses of Diverse Bartonella Species in Bat Ectoparasites Collected from Yunnan Province, China. Pathogens. 2022; 11(11):1283. https://doi.org/10.3390/pathogens11111283
Chicago/Turabian StyleKuang, Guopeng, Jing Zhang, Weihong Yang, Hong Pan, Xi Han, Lifen Yang, Juan Wang, Tian Yang, Zhizhong Song, Yun Feng, and et al. 2022. "Molecular Detection and Phylogenetic Analyses of Diverse Bartonella Species in Bat Ectoparasites Collected from Yunnan Province, China" Pathogens 11, no. 11: 1283. https://doi.org/10.3390/pathogens11111283
APA StyleKuang, G., Zhang, J., Yang, W., Pan, H., Han, X., Yang, L., Wang, J., Yang, T., Song, Z., Feng, Y., & Liang, G. (2022). Molecular Detection and Phylogenetic Analyses of Diverse Bartonella Species in Bat Ectoparasites Collected from Yunnan Province, China. Pathogens, 11(11), 1283. https://doi.org/10.3390/pathogens11111283