
Avian Bornaviruses in Wild Aquatic Birds of the Anseriformes Order in Poland


Abstract
1. Introduction
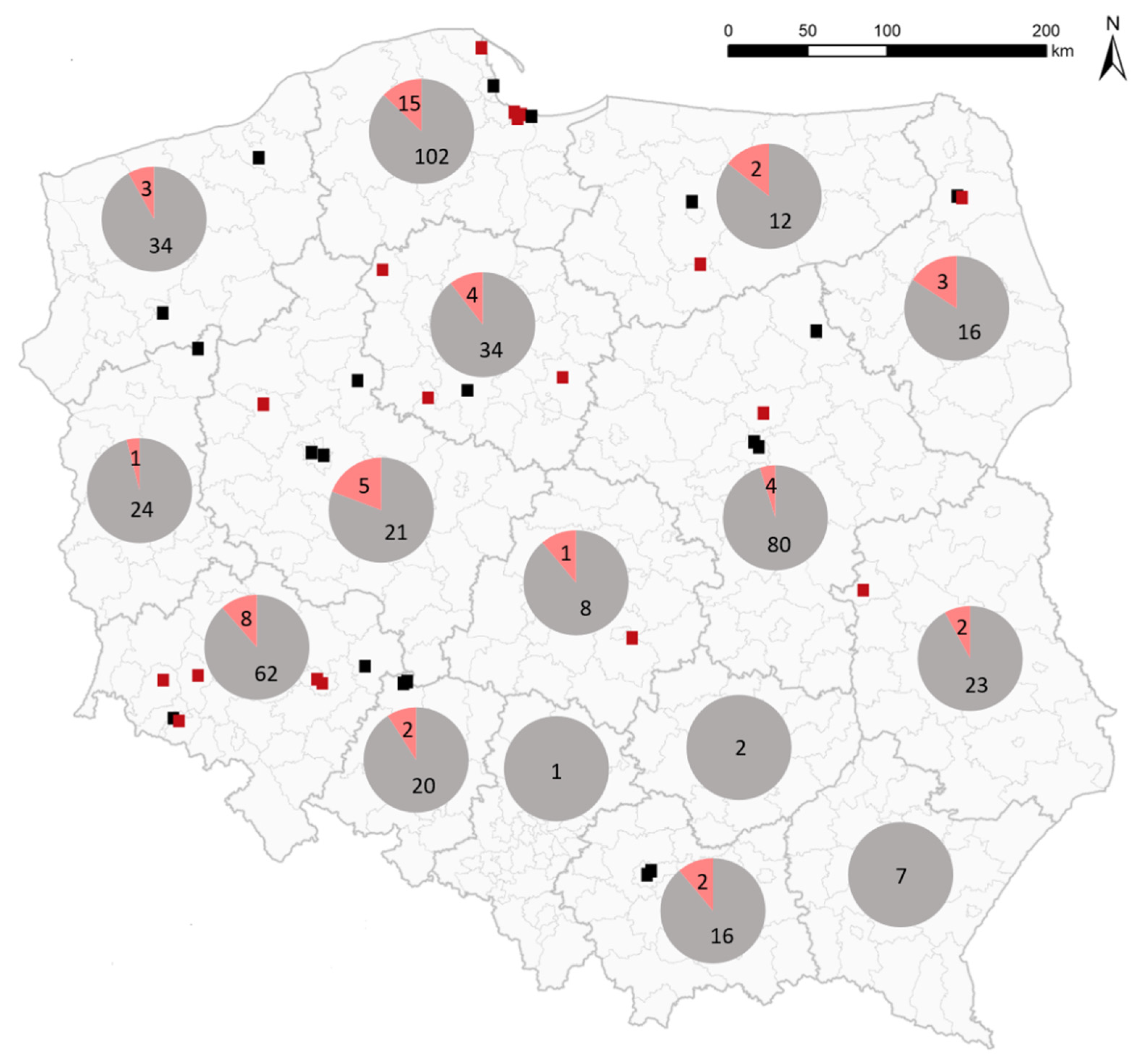
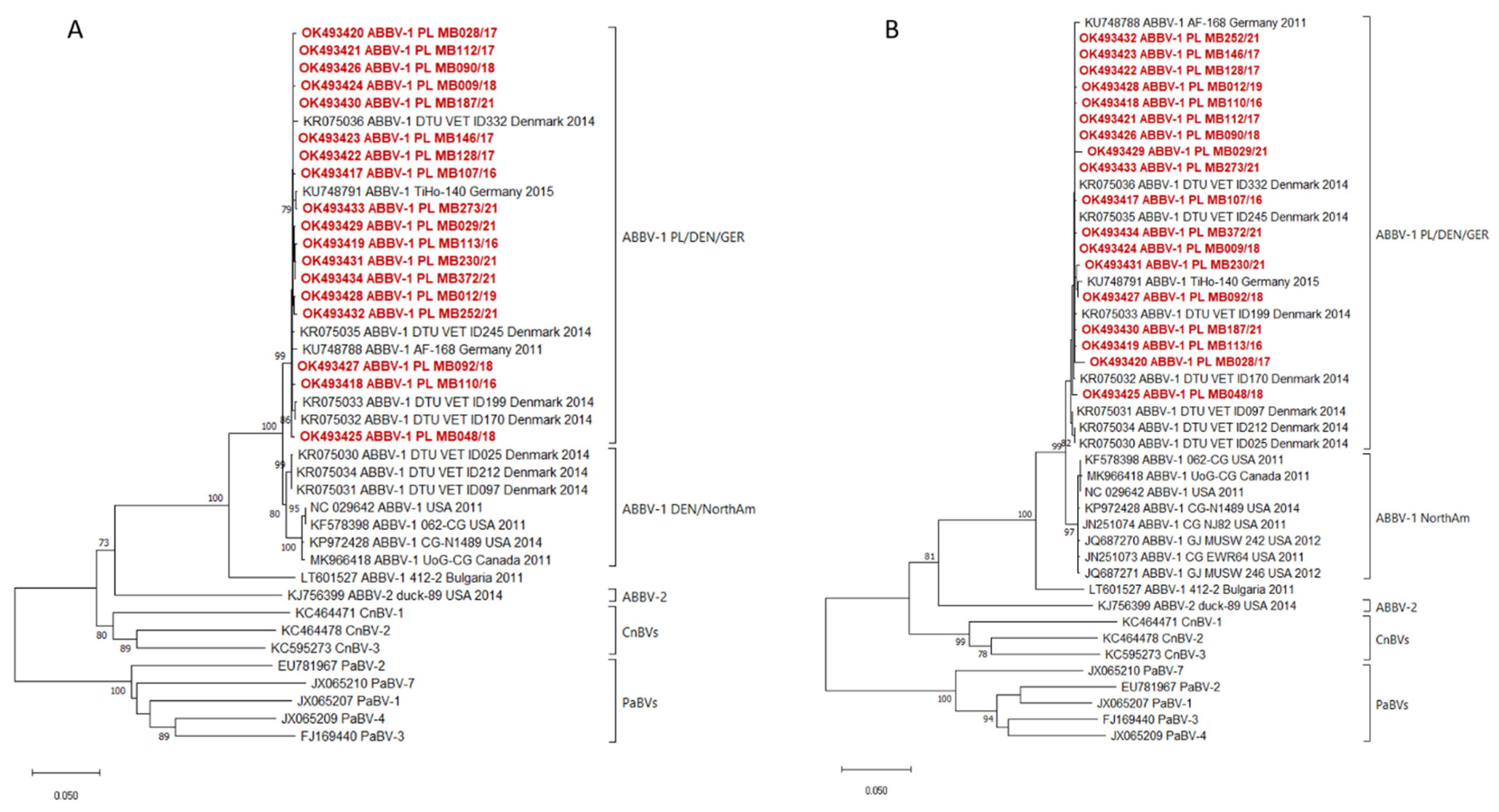
2. Results
3. Discussion
4. Materials and Methods
4.1. Samples from Wild Birds
4.2. Detection of ABBVs RNA with Real Time RT-PCR
4.3. Sequencing and Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rubbenstroth, D.; Briese, T.; Dürrwald, R.; Horie, M.; Hyndman, T.H.; Kuhn, J.H.; Nowotny, N.; Payne, S.; Stenglein, M.D.; Tomonaga, K.; et al. ICTV Virus Taxonomy Profile: Bornaviridae. J. Gen. Virol. 2021, 102, 001613. [Google Scholar] [CrossRef] [PubMed]
- Gancz, A.Y.; Clubb, S.; Shivaprasad, H.L. Advanced diagnostic approaches and current management of proventricular dilatation disease. Vet. Clin. N. Am. Exot. Anim. Pract. 2010, 13, 471–494. [Google Scholar] [CrossRef] [PubMed]
- Staeheli, P.; Rinder, M.; Kaspers, B. Avian bornavirus associated with fatal disease in psittacine birds. J. Virol. 2010, 84, 6269–6275. [Google Scholar] [CrossRef] [PubMed]
- Weissenböck, H.; Sekulin, K.; Bakonyi, T.; Högler, S.; Nowotny, N. Novel avian bornavirus in a nonpsittacine species (Canary; Serinus canaria) with enteric ganglioneuritis and encephalitis. J. Virol. 2009, 83, 11367–11371. [Google Scholar] [CrossRef] [PubMed]
- Delnatte, P.; Berkvens, C.; Kummrow, M.; Smith, D.A.; Campbell, D.; Crawshaw, G.; Ojkic, D.; DeLay, J. New genotype of avian bornavirus in wild geese and trumpeter swans in Canada. Vet. Rec. 2011, 169, 108. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Covaleda, L.; Heatley, J.J.; Baroch, J.A.; Tizard, I.; Payne, S.L. Widespread avian bornavirus infection in mute swans in the Northeast United States. Vet. Med. 2012, 3, 49–52. [Google Scholar]
- Delnatte, P.; Ojkic, D.; DeLay, J.; Campbell, D.; Crawshaw, G.; Smith, D.A. Pathology and diagnosis of avian bornavirus infection in wild Canada geese (Branta canadensis), trumpeter swans (Cygnus buccinator) and mute swans (Cygnus olor) in Canada: A retrospective study. Avian. Pathol. 2013, 42, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Payne, S.; Covaleda, L.; Jianhua, G.; Swafford, S.; Baroch, J.; Ferro, P.J.; Lupiani, B.; Heatley, J.; Tizard, I. Detection and characterization of a distinct bornavirus lineage from healthy Canada geese (Branta canadensis). J. Virol. 2011, 85, 12053–12066. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Guo, J.; Tizard, I.; Baroch, J.; Shivaprasad, H.L.; Payne, S.L. Avian Bornaviruses in North American Gulls. J. Wildl. Dis. 2015, 51, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Guo, J.; Tizard, I.; Jennings, S.; Shivaprasad, H.L.; Payne, S.; Ellis, J.C.; Van Wettere, A.J.; O’Brien, K.M. Aquatic Bird Bornavirus-Associated Disease in Free-Living Canada Geese (Branta canadensis) in the Northeastern USA. J. Wildl. Dis. 2017, 53, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Shivaprasad, H.L.; Rech, R.R.; Heatley, J.J.; Tizard, I.; Payne, S. Characterization of a new genotype of avian bornavirus from wild ducks. Virol. J. 2014, 11, 197. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, A.F.; Nielsen, J.B.; Hjulsager, C.K.; Chriél, M.; Smith, D.A.; Bertelsen, M.F. Aquatic Bird Bornavirus 1 in Wild Geese, Denmark. Emerg. Infect. Dis. 2015, 21, 2201–2203. [Google Scholar] [CrossRef] [PubMed]
- Rubbenstroth, D.; Schmidt, V.; Rinder, M.; Legler, M.; Twietmeyer, S.; Schwemmer, P.; Corman, V.M. Phylogenetic Analysis Supports Horizontal Transmission as a Driving Force of the Spread of Avian Bornaviruses. PLoS ONE 2016, 11, e0160936. [Google Scholar] [CrossRef] [PubMed]
- Rubbenstroth, D.; Schlottau, K.; Schwemmle, M.; Rissland, J.; Beer, M. Human bornavirus research: Back on track! PLoS Pathog. 2019, 15, e1007873. [Google Scholar] [CrossRef] [PubMed]
- Rinder, M.; Ackermann, A.; Kempf, H.; Kaspers, B.; Korbel, R.; Staeheli, P. Broad tissue and cell tropism of avian bornavirus in parrots with proventricular dilatation disease. J. Virol. 2009, 83, 5401–5407. [Google Scholar] [CrossRef] [PubMed]
- Delnatte, P.; Mak, M.; Ojkic, D.; Raghav, R.; DeLay, J.; Smith, D.A. Detection of Avian bornavirus in multiple tissues of infected psittacine birds using real-time reverse transcription polymerase chain reaction. J. Vet. Diagn. Invest. 2014, 26, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Delnatte, P.; Nagy, É.; Ojkic, D.; Leishman, D.; Crawshaw, G.; Elias, K.; Smith, D.A. Avian bornavirus in free-ranging waterfowl: Prevalence of antibodies and cloacal shedding of viral RNA. J. Wildl. Dis. 2014, 50, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Sigrist, B.; Geers, J.; Albini, S.; Rubbenstroth, D.; Wolfrum, N. A New Multiplex Real-Time RT-PCR for Simultaneous Detection and Differentiation of Avian Bornaviruses. Viruses 2021, 13, 1358. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Number of Tested Birds | Number of ABBV-Positive Birds |
|---|---|---|
| mute swan (Cygnus olor) | 220 | 36 |
| whooper swan (Cygnus cygnus) | 2 | 1 |
| black swan (Cygnus atratus) | 9 | 0 |
| swan (Cygnus sp.) 1 | 170 | 15 |
| mallard (Anas platyrhynchos) | 48 | 0 |
| long-tailed duck (Clangula hyemalis) | 11 | 0 |
| garganey (Spatula querquedula) | 3 | 0 |
| wild duck 2 | 31 | 0 |
| other ducks 3 | 3 | 0 |
| bean goose (Anser fabalis) | 7 | 0 |
| tundra bean goose (Anser serrirostris) | 3 | 0 |
| greylag goose (Anser anser) | 3 | 0 |
| white-fronted goose (Anser albifrons) | 1 | 0 |
| wild goose 2 | 3 | 0 |
| Total | 514 | 52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Świętoń, E.; Dziadek, K.; Śmietanka, K. Avian Bornaviruses in Wild Aquatic Birds of the Anseriformes Order in Poland. Pathogens 2022, 11, 98. https://doi.org/10.3390/pathogens11010098
Świętoń E, Dziadek K, Śmietanka K. Avian Bornaviruses in Wild Aquatic Birds of the Anseriformes Order in Poland. Pathogens. 2022; 11(1):98. https://doi.org/10.3390/pathogens11010098
Chicago/Turabian StyleŚwiętoń, Edyta, Kamila Dziadek, and Krzysztof Śmietanka. 2022. "Avian Bornaviruses in Wild Aquatic Birds of the Anseriformes Order in Poland" Pathogens 11, no. 1: 98. https://doi.org/10.3390/pathogens11010098
APA StyleŚwiętoń, E., Dziadek, K., & Śmietanka, K. (2022). Avian Bornaviruses in Wild Aquatic Birds of the Anseriformes Order in Poland. Pathogens, 11(1), 98. https://doi.org/10.3390/pathogens11010098