
Treatment of Human Babesiosis: Then and Now


Abstract
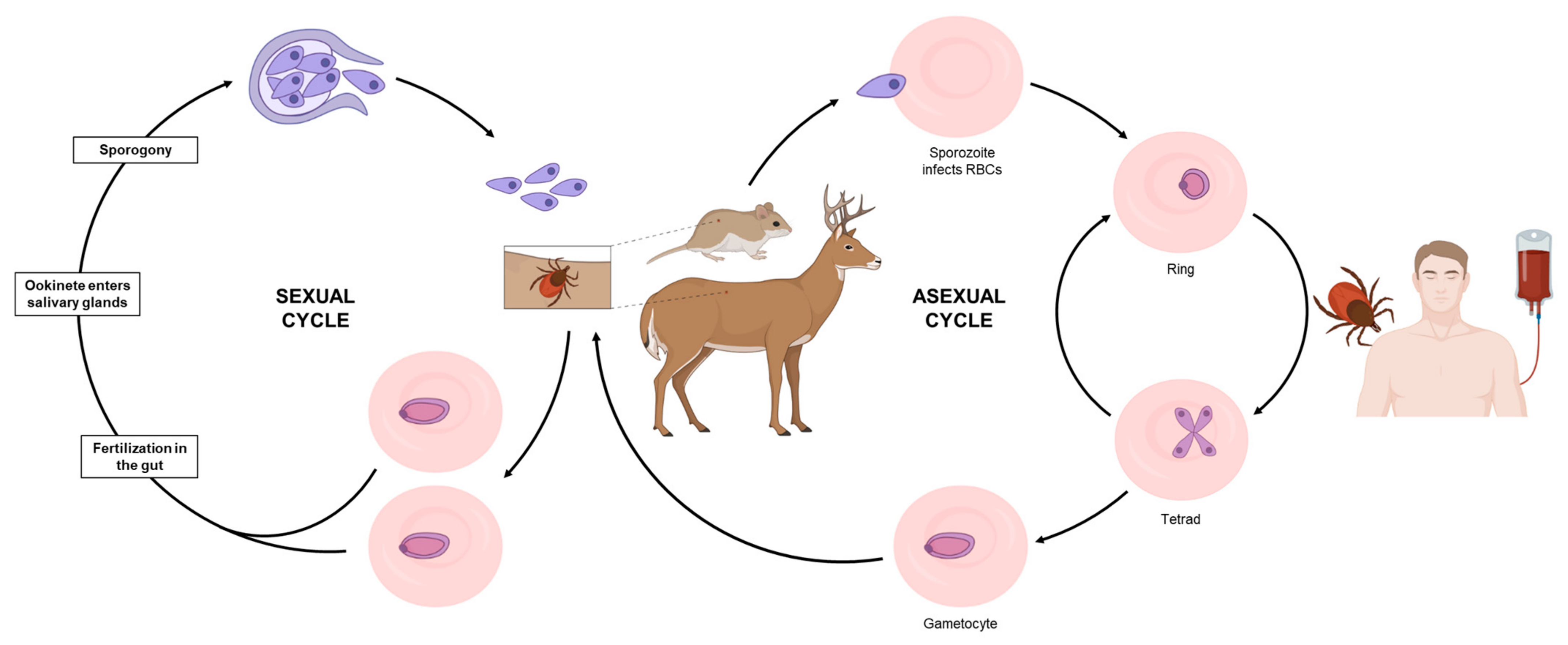
1. Introduction
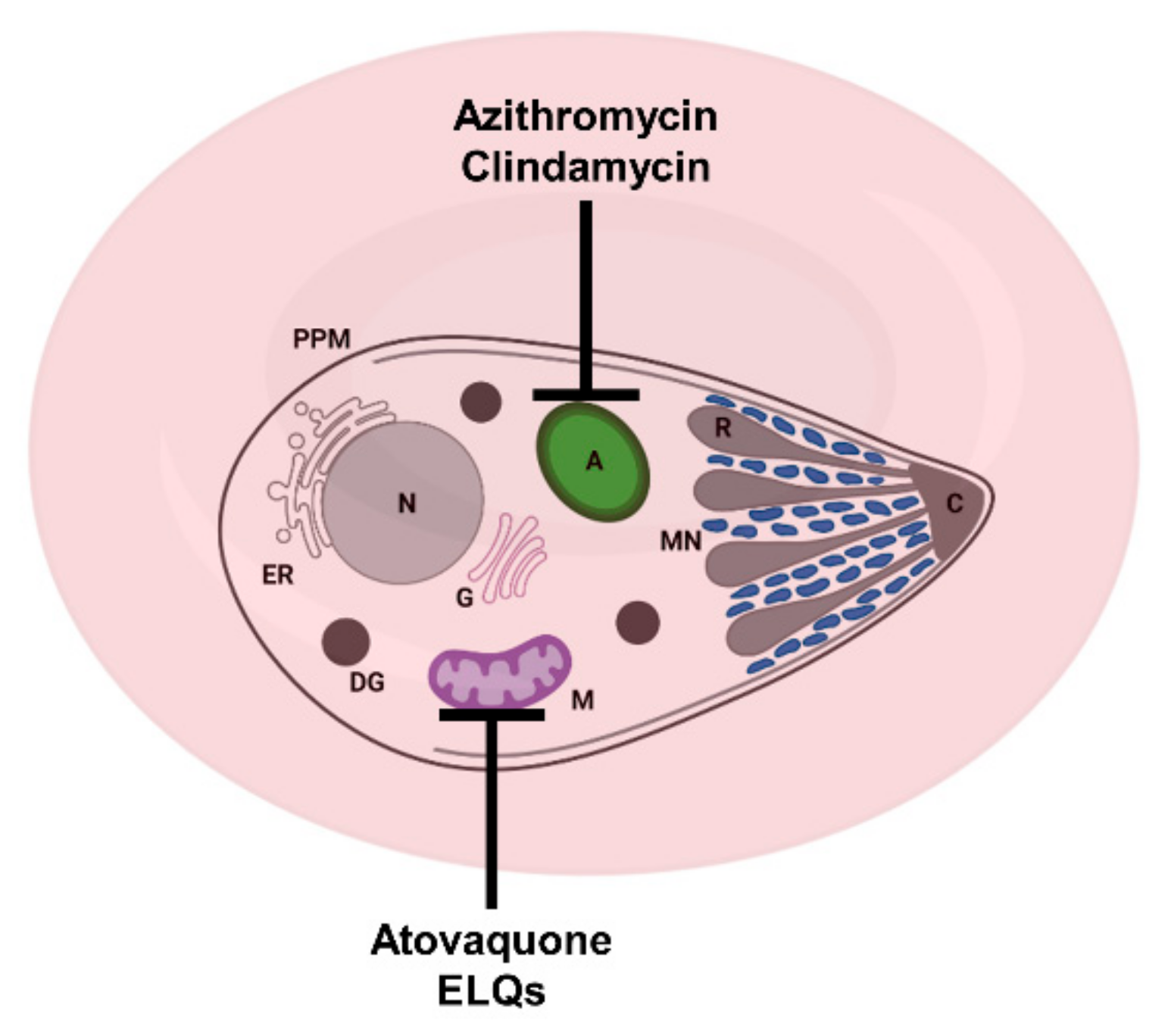
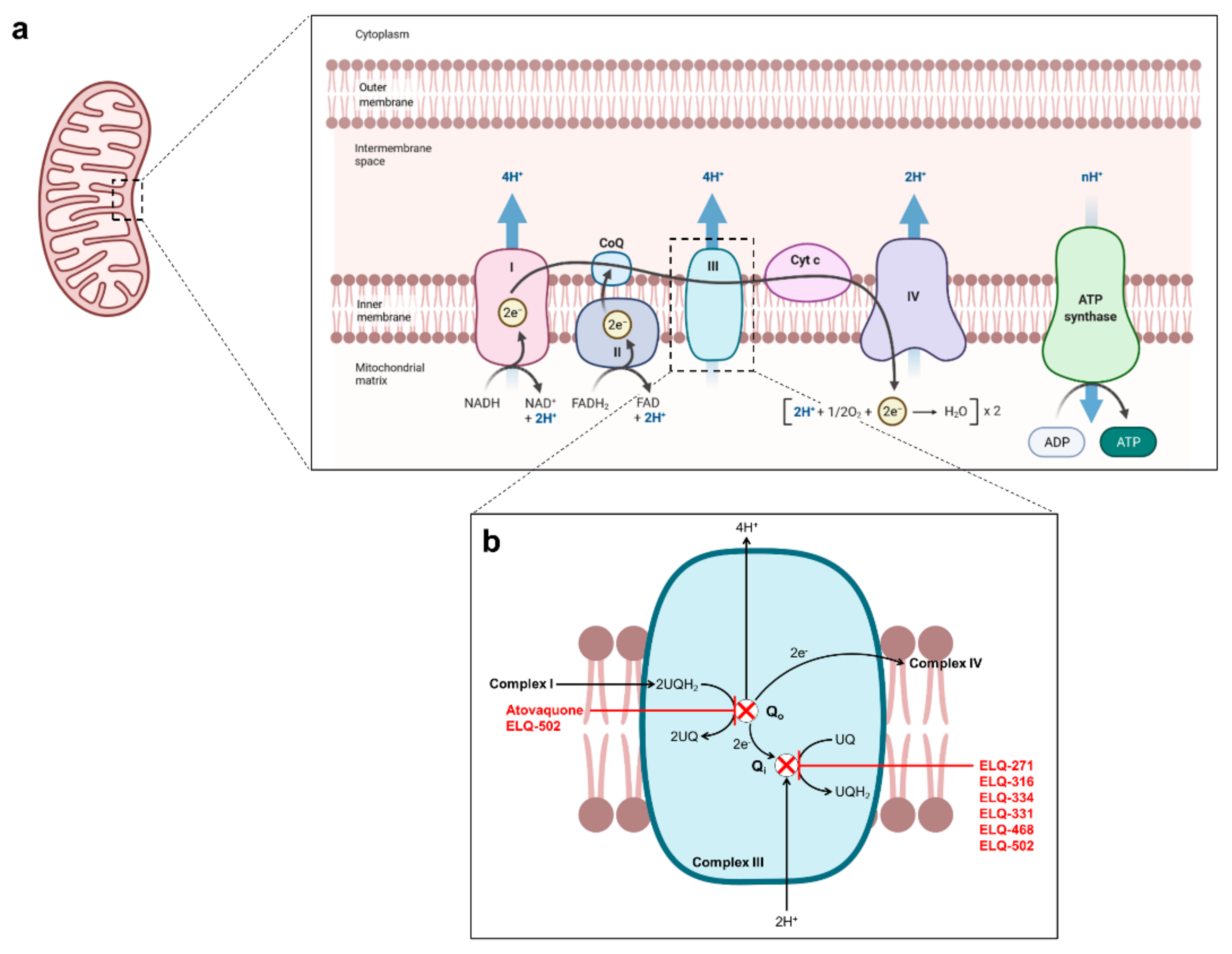
2. Current Treatments against Human Babesiosis

{kind=link}
{kind=link}
{kind=link}
| Drug | Treatment Regimen | Model | Effect | Ref. |
|---|---|---|---|---|
| Atovaquone | 20 mg/kg (p.o.), 5 d | B. microti Balb/c mice | ~5.7 × reduction in peak parasitemia. | [61] |
| 25 mg/kg (p.o.), 4 d | B. microti Balb/c mice | 77% suppression of parasitemia at DPI 9. | [54] | |
| 50 mg/kg (p.o.), 4 d | B. microti Balb/c mice | 87% suppression of parasitemia at DPI 9. | [54] | |
| 100 mg/kg (p.o.), 4 d | B. microti Balb/c mice | 93% suppression of parasitemia at DPI 9. | [54] | |
| 10 mg/kg (p.o.), 7 d | B. microti SCID mice | Parasitemia clearance followed by recrudescence by D5-9 post-treatment. | [11] | |
| 10 mg/kg (p.o.), 10 d | B. microti SCID mice | Parasitemia clearance followed by recrudescence by D14 post-treatment. | [57] | |
| 10 mg/kg (p.o.), 10 d | B. duncani C3H/HeJ mice | Parasitemia clearance followed by recrudescence by D10 post-treatment. 80% survival. | [57] | |
| Azithromycin | 25 mg/kg (p.o.), 4 d | B. microti Balb/c mice | 75% suppression of parasitemia at DPI 9. | [54] |
| 50 mg/kg (p.o.), 4 d | B. microti Balb/c mice | 96% suppression of parasitemia at DPI 9. | [54] | |
| 100 mg/kg (p.o.), 4 d | B. microti Balb/c mice | 95% suppression of parasitemia at DPI 9. | [54] | |
| 10 mg/kg (p.o.), 7 d | B. microti SCID mice | No effect. | [11] | |
| 50 mg/kg (p.o.), 7 d | B. microti SCID mice | No effect. | [11] | |
| Clindamycin | 300 mg/kg (p.o.), 5d | B. microti Mongolian jirds | 9.4% suppression of parasitemia at DPI 9. | [51] |
| 150 mg/kg (i.m.), 8d | B. microti Golden hamsters | ~2× reduction in peak parasitemia. | [52] | |
| 150 mg/kg (p.o.), 8d | B. microti Golden hamsters | ~2× reduction in peak parasitemia. | [52] | |
| 500 mg/kg (i.p.), 5d | B. microti Balb/c mice | ~3.2× reduction in peak parasitemia. | [53] | |
| 25 mg/kg (p.o.), 4 d | B. microti Balb/c mice | No effect. | [54] | |
| 50 mg/kg (p.o.), 4 d | B. microti Balb/c mice | No effect. | [54] | |
| 100 mg/kg (p.o.), 4 d | B. microti Balb/c mice | No effect. | [54] | |
| 10 mg/kg (p.o.), 7 d | B. microti SCID mice | No effect. | [11] | |
| 50 mg/kg (p.o.), 7 d | B. microti SCID mice | No effect. | [11] | |
| Quinine | 125 mg/kg (s.c.), 8d | B. microti Golden hamsters | No effect. | [52] |
| 250 mg/kg (p.o.), 8d | B. microti Golden hamsters | No effect. | [52] | |
| 25 mg/kg (p.o.), 4 d | B. microti Balb/c mice | No effect. | [54] | |
| 50 mg/kg (p.o.), 4 d | B. microti Balb/c mice | No effect. | [54] | |
| 100 mg/kg (p.o.), 4 d | B. microti Balb/c mice | No effect. | [54] | |
| 10 mg/kg (p.o.), 7 d | B. microti SCID mice | No effect. | [11] | |
| 50 mg/kg (p.o.), 7 d | B. microti SCID mice | No effect. | [11] | |
| 100 mg/kg (p.o.), 7 d | B. microti SCID mice | No effect. | [11] |
3. In Vitro and In Vivo Models for the Evaluation of Novel Anti-Babesia Therapies
4. Novel Therapies under Investigation for the Treatment of Human Babesiosis
5. Conclusions and Considerations for Future Drug Development
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krause, P.J. Human babesiosis. Int. J. Parasit. 2019, 49, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging Tick-Borne Diseases. Clin. Microbiol. Rev. 2020, 33, 34. [Google Scholar] [CrossRef]
- Levin, A.E.; Krause, P.J. Transfusion-transmitted babesiosis: Is it time to screen the blood supply? Curr. Opin. Hematol. 2016, 23, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Lobo, C.A.; Singh, M.; Rodriguez, M. Human babesiosis: Recent advances and future challenges. Curr. Opin. Hematol. 2020, 27, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Moritz, E.D.; Winton, C.S.; Tonnetti, L.; Townsend, R.L.; Berardi, V.P.; Hewins, M.E.; Weeks, K.E.; Dodd, R.Y.; Stramer, S.L. Screening for Babesia microti in the U. S. Blood Supply. N. Engl. J. Med. 2016, 375, 2236–2245. [Google Scholar] [CrossRef] [PubMed]
- Tonnetti, L.; Townsend, R.L.; Dodd, R.Y.; Stramer, S.L. Characteristics of transfusion-transmitted Babesia microti, American Red Cross 2010–2017. Transfusion 2019, 59, 2908–2912. [Google Scholar] [CrossRef]
- Mantadakis, E. Pneumocystis jirovecii Pneumonia in Children with Hematological Malignancies: Diagnosis and Approaches to Management. J. Fungi 2020, 6, 331. [Google Scholar] [CrossRef] [PubMed]
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of Toxoplasmosis: Historical Perspective, Animal Models, and Current Clinical Practice. Clin. Microbiol. Rev. 2018, 31, e00057-17. [Google Scholar] [CrossRef]
- Nixon, G.L.; Moss, D.M.; Shone, A.E.; Lalloo, D.G.; Fisher, N.; O’Neill, P.M.; Ward, S.A.; Biagini, G.A. Antimalarial pharmacology and therapeutics of atovaquone. J. Antimicrob. Chemother. 2013, 68, 977–985. [Google Scholar] [CrossRef]
- Jacobsen, L.; Husen, P.; Solov’yov, I.A. Inhibition Mechanism of Antimalarial Drugs Targeting the Cytochrome bc1 Complex. J. Chem. Inf. Modeling 2021, 61, 1334–1345. [Google Scholar] [CrossRef]
- Lawres, L.A.; Garg, A.; Kumar, V.; Bruzual, I.; Forquer, I.P.; Renard, I.; Virji, A.Z.; Boulard, P.; Rodriguez, E.X.; Allen, A.J.; et al. Radical cure of experimental babesiosis in immunodeficient mice using a combination of an endochin-like quinolone and atovaquone. J. Exp. Med. 2016, 213, 1307–1318. [Google Scholar] [CrossRef]
- Montazeri, M.; Mehrzadi, S.; Sharif, M.; Sarvi, S.; Shahdin, S.; Daryani, A. Activities of anti-Toxoplasma drugs and compounds against tissue cysts in the last three decades (1987 to 2017), a systematic review. Parasitol. Res. 2018, 117, 3045–3057. [Google Scholar] [CrossRef]
- Vaidya, A.B.; Mather, M.W. Atovaquone resistance in malaria parasites. Drug Resist. Updates 2000, 3, 283–287. [Google Scholar] [CrossRef]
- Daniel, R. Azithromycin, erythromycin and cloxacillin in the treatment of infections of skin and associated soft tissues. European Azithromycin Study Group. J. Int. Med. Res. 1991, 19, 433–445. [Google Scholar] [CrossRef]
- Dinwiddie, R. Anti-inflammatory therapy in cystic fibrosis. J. Cyst. Fibros. 2005, 4 (Suppl. S2), 45–48. [Google Scholar] [CrossRef] [PubMed]
- Ladhani, S.; Garbash, M. Staphylococcal skin infections in children: Rational drug therapy recommendations. Paediatr. Drugs 2005, 7, 77–102. [Google Scholar] [CrossRef] [PubMed]
- Carratala, J.; Garcia-Vidal, C. An update on Legionella. Curr. Opin. Infect. Dis. 2010, 23, 152–157. [Google Scholar] [CrossRef]
- Van Eijk, A.M.; Terlouw, D.J. Azithromycin for treating uncomplicated malaria. Cochrane Database Syst. Rev. 2011, 2011, Cd006688. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A. Understanding the biology of the Plasmodium falciparum apicoplast; an excellent target for antimalarial drug development. Life Sci. 2016, 158, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, A.B.S.; Sun, Q.G.; Nkrumah, L.J.; Dunne, M.W.; Sacchettini, J.C.; Fidock, D.A. In vitro efficacy, resistance selection, and structural modeling studies implicate the malarial parasite apicoplast as the target of azithromycin. J. Biol. Chem. 2007, 282, 2494–2504. [Google Scholar] [CrossRef] [PubMed]
- Beckers, C.J.M.; Roos, D.S.; Donald, R.G.K.; Luft, B.J.; Schwab, J.C.; Cao, Y.; Joiner, K.A. Inhibition of cytoplasmic and organellar protein synthesis in Toxoplasma gondii. Implications for the target of macrolide antibiotics. J. Clin. Investig. 1995, 95, 367–376. [Google Scholar] [CrossRef]
- Dahl, E.L.; Rosenthal, P.J. Multiple antibiotics exert delayed effects against the Plasmodium falciparum anicoplast. Antimicrob. Agents Chemother. 2007, 51, 3485–3490. [Google Scholar] [CrossRef]
- Murphy, P.B.; Bistas, K.G.; Le, J.K. Clindamycin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Griffith, K.S.; Lewis, L.S.; Mali, S.; Parise, M.E. Treatment of malaria in the United States: A systematic review. Jama 2007, 297, 2264–2277. [Google Scholar] [CrossRef]
- Lell, B.; Kremsner, P.G. Clindamycin as an antimalarial drug: Review of clinical trials. Antimicrob. Agents Chemother. 2002, 46, 2315–2320. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.P.; Hunfeld, K.P.; Krause, P.J. Management strategies for human babesiosis. Expert Rev. Anti-Infect. Ther. 2020, 18, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Pfefferkorn, E.R.; Borotz, S.E. Comparison of mutants of Toxoplasma gondii selected for resistance to azithromycin, spiramycin, or clindamycin. Antimicrob. Agents Chemother. 1994, 38, 31–37. [Google Scholar] [CrossRef]
- Talapko, J.; Škrlec, I.; Alebić, T.; Jukić, M.; Včev, A. Malaria: The Past and the Present. Microorganisms 2019, 7, 179. [Google Scholar] [CrossRef] [PubMed]
- Tse, E.G.; Korsik, M.; Todd, M.H. The past, present and future of anti-malarial medicines. Malar. J. 2019, 18, 93. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, D.J., Jr.; Gluzman, I.Y.; Russell, D.G.; Goldberg, D.E. On the molecular mechanism of chloroquine’s antimalarial action. Proc. Natl. Acad. Sci. USA 1996, 93, 11865–11870. [Google Scholar] [CrossRef]
- Tang, Y.Q.; Ye, Q.; Huang, H.; Zheng, W.Y. An Overview of Available Antimalarials: Discovery, Mode of Action and Drug Resistance. Curr. Mol. Med. 2020, 20, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Woodland, J.G.; Hunter, R.; Smith, P.J.; Egan, T.J. Shining new light on ancient drugs: Preparation and subcellular localisation of novel fluorescent analogues of Cinchona alkaloids in intraerythrocytic Plasmodium falciparum. Org. Biomol. Chem. 2017, 15, 589–597. [Google Scholar] [CrossRef]
- Woodland, J.G.; Hunter, R.; Smith, P.J.; Egan, T.J. Chemical Proteomics and Super-resolution Imaging Reveal That Chloroquine Interacts with Plasmodium falciparum Multidrug Resistance-Associated Protein and Lipids. ACS Chem. Biol. 2018, 13, 2939–2948. [Google Scholar] [CrossRef] [PubMed]
- Punihaole, D.; Workman, R.J.; Upadhyay, S.; Van Bruggen, C.; Schmitz, A.J.; Reineke, T.M.; Frontiera, R.R. New Insights into Quinine-DNA Binding Using Raman Spectroscopy and Molecular Dynamics Simulations. J. Phys. Chem. B 2018, 122, 9840–9851. [Google Scholar] [CrossRef] [PubMed]
- Golan, D.E.; Armstrong, E.J.; Armstrong, A.W.; Tashjian, A.H. Principles of Pharmacology: The Pathophysiologic Basis of Drug Therapy, 3rd ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012; pp. 1–1956. [Google Scholar]
- Percário, S.; Moreira, D.R.; Gomes, B.A.; Ferreira, M.E.; Gonçalves, A.C.; Laurindo, P.S.; Vilhena, T.C.; Dolabela, M.F.; Green, M.D. Oxidative stress in malaria. Int. J. Mol. Sci. 2012, 13, 16346–16372. [Google Scholar] [CrossRef]
- Dziekan, J.M.; Yu, H.; Chen, D.; Dai, L.; Wirjanata, G.; Larsson, A.; Prabhu, N.; Sobota, R.M.; Bozdech, Z.; Nordlund, P. Identifying purine nucleoside phosphorylase as the target of quinine using cellular thermal shift assay. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef]
- Krause, P.J.; Auwaerter, P.G.; Bannuru, R.R.; Branda, J.A.; Falck-Ytter, Y.T.; Lantos, P.M.; Lavergne, V.; Meissner, H.C.; Osani, M.C.; Rips, J.G.; et al. Clinical Practice Guidelines by the Infectious Diseases Society of America (IDSA): 2020 Guideline on Diagnosis and Management of Babesiosis. Clin. Infect. Dis. 2021, 72, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Kletsova, E.A.; Spitzer, E.D.; Fries, B.C.; Marcos, L.A. Babesiosis in Long Island: Review of 62 cases focusing on treatment with azithromycin and atovaquone. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 7. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Gewurz, B.E.; Hill, D.; Marty, F.M.; Vannier, E.; Foppa, I.M.; Furman, R.R.; Neuhaus, E.; Skowron, G.; Gupta, S.; et al. Persistent and relapsing babesiosis in Immunocompromised patients. Clin. Infect. Dis. 2008, 46, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Raffalli, J.; Wormser, G.P. Persistence of babesiosis for >2 years in a patient on rituximab for rheumatoid arthritis. Diagn. Microbiol. Infect. Dis. 2016, 85, 231–232. [Google Scholar] [CrossRef]
- Simon, M.S.; Westblade, L.F.; Dziedziech, A.; Visone, J.E.; Furman, R.R.; Jenkins, S.G.; Schuetz, A.N.; Kirkman, L.A. Clinical and Molecular Evidence of Atovaquone and Azithromycin Resistance in Relapsed Babesia microti Infection Associated with Rituximab and Chronic Lymphocytic Leukemia. Clin. Infect. Dis. 2017, 65, 1222–1225. [Google Scholar] [CrossRef]
- Lemieux, J.E.; Tran, A.D.; Freimark, L.; Schaffner, S.F.; Goethert, H.; Andersen, K.G.; Bazner, S.; Li, A.; McGrath, G.; Sloan, L.; et al. A global map of genetic diversity in Babesia microti reveals strong population structure and identifies variants associated with clinical relapse. Nat. Microbiol. 2016, 1, 7. [Google Scholar] [CrossRef]
- Birth, D.; Kao, W.C.; Hunte, C. Structural analysis of atovaquone-inhibited cytochrome bc(1) complex reveals the molecular basis of antimalarial drug action. Nat. Commun. 2014, 5, 11. [Google Scholar] [CrossRef]
- Doktor, S.Z.; Shortridge, V.D.; Beyer, J.M.; Flamm, R.K. Epidemiology of macrolide and/or lincosamide resistant Streptococcus pneumoniae clinical isolates with ribosomal mutations. Diagn. Microbiol. Infect. Dis. 2004, 49, 47–52. [Google Scholar] [CrossRef]
- Li, Y.J.; Stanley, S.; Villalba, J.A.; Nelson, S.; Gelfand, J. Case Report: Overwhelming Babesia Parasitemia Successfully Treated Promptly with RBC Apheresis and Triple Therapy with Clindamycin, Azithromycin, and Atovaquone. Open Forum Infect. Dis. 2020, 7, 3. [Google Scholar] [CrossRef]
- Vyas, J.M.; Telford, S.R.; Robbins, G.K. Treatment of refractory Babesia microti infection with atovaquone-proguanil in an HIV-infected patient: Case report. Clin. Infect. Dis. 2007, 45, 1588–1590. [Google Scholar] [CrossRef]
- Man, S.Q.; Qiao, K.; Cui, J.; Feng, M.; Fu, Y.F.; Cheng, X.J. A case of human infection with a novel Babesia species in China. Infect. Dis. Poverty 2016, 5, 6. [Google Scholar] [CrossRef]
- Gao, J.C.; Gong, Z.D.; Montesano, D.; Glazer, E.; Liegner, K. “Repurposing” Disulfiram in the Treatment of Lyme Disease and Babesiosis: Retrospective Review of First 3 Years’ Experience in One Medical Practice. Antibiotics 2020, 9, 868. [Google Scholar] [CrossRef]
- Radcliffe, C.; Krause, P.J.; Grant, M. Repeat exchange transfusion for treatment of severe babesiosis. Transfus. Apher. Sci. 2019, 58, 638–640. [Google Scholar] [CrossRef] [PubMed]
- Ruebush, T.K.; Contacos, P.G.; Steck, E.A. Chemotherapy of Babesia microti infections in Mongolian Jirds. Antimicrob. Agents Chemother. 1980, 18, 289–291. [Google Scholar] [CrossRef] [PubMed]
- Rowin, K.S.; Tanowitz, H.B.; Wittner, M. Therapy of Experimental Babesiosis. Ann. Intern. Med. 1982, 97, 556–558. [Google Scholar] [CrossRef]
- AbouLaila, M.; Munkhjargal, T.; Sivakumar, T.; Ueno, A.; Nakano, Y.; Yokoyama, M.; Yoshinari, T.; Nagano, D.; Katayama, K.; El-Bahy, N.; et al. Apicoplast-Targeting Antibacterials Inhibit the Growth of Babesia Parasites. Antimicrob. Agents Chemother. 2012, 56, 3196–3206. [Google Scholar] [CrossRef]
- Yao, J.M.; Zhang, H.B.; Liu, C.S.; Tao, Y.; Yin, M. Inhibitory effects of 19 antiprotozoal drugs and antibiotics on Babesia microti infection in BALB/c mice. J. Infect. Dev. Ctries. 2015, 9, 1004–1010. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Marley, S.E.; Eberhard, M.L.; Steurer, F.J.; Ellis, W.L.; McGreevy, P.B.; Ruebush, T.K. Evaluation of selected antiprotozoal drugs in the Babesia microti-hamster model. Antimicrob. Agents Chemother. 1997, 41, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Wittner, M.; Lederman, J.; Tanowitz, H.B.; Rosenbaum, G.S.; Weiss, L.M. Atovaquone in the treatment of Babesia microti infections in hamsters. Am. J. Trop. Med. Hyg. 1996, 55, 219–222. [Google Scholar] [CrossRef]
- Chiu, J.E.; Renard, I.; Pal, A.C.; Singh, P.; Vydyam, P.; Thekkiniath, J.; Kumar, M.; Gihaz, S.; Pou, S.; Winter, R.W.; et al. Effective Therapy Targeting Cytochrome bc(1) Prevents Babesia Erythrocytic Development and Protects from Lethal Infection. Antimicrob. Agents Chemother. 2021, 65, AAC-00662. [Google Scholar] [CrossRef] [PubMed]
- Hughes, W.T.; Oz, H.S. Successful Prevention and Treatment of Babesiosis with Atovaquone. J. Infect. Dis. 1995, 172, 1042–1046. [Google Scholar] [CrossRef] [PubMed]
- Pudney, M.; Gray, J.S. Therapeutic efficacy of atovaquone against the bovine intraerythrocytic parasite, Babesia divergens. J. Parasitol. 1997, 83, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J. Babesiosis. Med. Clin. North. Am. 2002, 86, 361–373. [Google Scholar] [CrossRef]
- Beshbishy, A.M.; Batiha, G.E.S.; Alkazmi, L.; Nadwa, E.; Rashwan, E.; Abdeen, A.; Yokoyama, N.; Igarashi, I. Therapeutic Effects of Atranorin towards the Proliferation of Babesia and Theileria Parasites. Pathogens 2020, 9, 127. [Google Scholar] [CrossRef]
- Chen, D.; Copeman, D.B.; Hutchinson, G.W.; Burnell, J. Inhibition of growth of cultured Babesia microti by serum and macrophages in the presence or absence of T cells. Parasitol. Int. 2000, 48, 223–231. [Google Scholar] [CrossRef]
- Matsubara, J.; Koura, M.; Kamiyama, T. Infection of Immunodeficient Mice with a Mouse-Adapted Substrain of the Gray Strain of Babesia microti. J. Parasitol. 1993, 79, 783–786. [Google Scholar] [CrossRef]
- Ruebush, M.J.; Hanson, W.L. Susceptibility of 5 Strains of Mice to Babesia microti of Human Origin. J. Parasitol. 1979, 65, 430–433. [Google Scholar] [CrossRef]
- Skariah, S.; Arnaboldi, P.; Dattwyler, R.J.; Sultan, A.A.; Gaylets, C.; Walwyn, O.; Mulhall, H.; Wu, X.; Dargham, S.R.; Mordue, D.G. Elimination of Babesia microti Is Dependent on Intraerythrocytic Killing and CD4(+) T Cells. J. Immunol. 2017, 199, 633–642. [Google Scholar] [CrossRef]
- Hu, R.J.; Yeh, M.T.; Hyland, K.E.; Mather, T.N. Experimental Babesia microti infection in golden hamsters: Immunoglobulin G response and recovery from severe hemolytic anemia. J. Parasitol. 1996, 82, 728–732. [Google Scholar] [CrossRef]
- Oz, H.S.; Hughes, W.T. Acute fulminating babesiosis in hamsters infected with Babesia microti. Int. J. Parasit. 1996, 26, 667–670. [Google Scholar] [CrossRef]
- Chauvin, A.; Valentin, A.; Malandrin, L.; L’Hostis, M. Sheep as a new experimental host for Babesia divergens. Vet. Res. 2002, 33, 429–433. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gorenflot, A.; Brasseur, P.; Precigout, E.; Lhostis, M.; Marchand, A.; Schrevel, J. Cytological and immunological responses to Babesia-divergens in different hosts- Ox, gerbil, man. Parasitol. Res. 1991, 77, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Musa, N.B.; Phillips, R.S. The adaptation of 3 isolates of Babesia-divergensto continuous culture in rat erythrocytes. Parasitology 1991, 103, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.S.; Moreira, C.K.; Elsworth, B.; Allred, D.R.; Duraisingh, M.T. Extensive Shared Chemosensitivity between Malaria and Babesiosis Blood-Stage Parasites. Antimicrob. Agents Chemother. 2016, 60, 5059–5063. [Google Scholar] [CrossRef] [PubMed]
- Vayrynen, R.; Tuomi, J. Continuous in vitro cultivation of Babesia divergens. Acta Vet. Scand. 1982, 23, 471–472. [Google Scholar] [CrossRef]
- Zintl, A.; Westbrook, C.; Mulcahy, G.; Skerrett, H.E.; Gray, J.S. Invasion, and short- and long-term survival of Babesia divergens (Phylum Apicomplexa) cultures in non-bovine sera and erythrocytes. Parasitology 2002, 124, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Entrican, J.H.; Williams, H.; Cook, I.A.; Lancaster, W.M.; Clark, J.C.; Joyner, L.P.; Lewis, D. Babesiosis in man- Report of a case from scotland with observations on the infecting strain. J. Infect. 1979, 1, 227–234. [Google Scholar] [CrossRef]
- Gray, J.S. Chemotherapy of Babesia divergens in the gerbil, Meriones unguiculatus. Res. Vet. Sci. 1983, 35, 318–324. [Google Scholar] [CrossRef]
- Brasseur, P.; Lecoublet, S.; Kapel, N.; Favennec, L.; Ballet, J.J. In vitro evaluation of drug susceptibilities of Babesia divergens isolates. Antimicrob. Agents Chemother. 1998, 42, 818–820. [Google Scholar] [CrossRef]
- Thomford, J.W.; Conrad, P.A.; Telford, S.R.; Mathiesen, D.; Bowman, B.H.; Spielman, A.; Eberhard, M.L.; Herwaldt, B.L.; Quick, R.E.; Persing, D.H. Cultivation and Phylogenetic Characterization of a Newly Recognized Human Pathogenic Protozoan. J. Infect. Dis. 1994, 169, 1050–1056. [Google Scholar] [CrossRef]
- McCormack, K.A.; Alhaboubi, A.; Pollard, D.A.; Fuller, L.; Holman, P.J. In vitro cultivation of Babesia duncani (Apicomplexa: Babesiidae), a zoonotic hemoprotozoan, using infected blood from Syrian hamsters (Mesocricetus auratus). Parasitol. Res. 2019, 118, 2409–2417. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Brasov, I.; Thekkiniath, J.; Kilian, N.; Lawres, L.; Gao, R.Y.; DeBus, K.; He, L.; Yu, X.; Zhu, G.; et al. Establishment of a continuous in vitro culture of Babesia duncani in human erythrocytes reveals unusually high tolerance to recommended therapies. J. Biol. Chem. 2018, 293, 19974–19981. [Google Scholar] [CrossRef] [PubMed]
- Dao, A.H.; Eberhard, M.L. Pathology of acute fatal babesiosis in hamsters experimentally infected with the WA-1 strain of Babesia. Lab. Investig. 1996, 74, 853–859. [Google Scholar]
- Wozniak, E.J.; Lowenstine, L.J.; Hemmer, R.; Robinson, T.; Conrad, P.A. Comparative pathogenesis of human WA1 and Babesia microti isolates in a Syrian hamster model. Lab. Anim. Sci. 1996, 46, 507–515. [Google Scholar] [PubMed]
- Aguilar-Delfin, I.; Homer, M.J.; Wettstein, P.J.; Persing, D.H. Innate resistance to Babesia infection is influenced by genetic background and gender. Infect. Immun. 2001, 69, 7955–7958. [Google Scholar] [CrossRef] [PubMed]
- Hemmer, R.M.; Wozniak, E.J.; Lowenstine, L.J.; Plopper, C.G.; Wong, V.; Conrad, P.A. Endothelial cell changes are associated with pulmonary edema and respiratory distress in mice infected with the WA1 human Babesia parasite. J. Parasitol. 1999, 85, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Spangenberg, T.; Burrows, J.N.; Kowalczyk, P.; McDonald, S.; Wells, T.N.C.; Willis, P. The Open Access Malaria Box: A Drug Discovery Catalyst for Neglected Diseases. PLoS ONE 2013, 8, 8. [Google Scholar] [CrossRef]
- Nugraha, A.B.; Tuvshintulga, B.; Guswanto, A.; Tayebwa, D.S.; Rizk, M.A.; Gantuya, S.; Batiha, G.E.; Beshbishy, A.M.; Sivakumar, T.; Yokoyama, N.; et al. Screening the Medicines for Malaria Venture Pathogen Box against piroplasm parasites. Int. J. Parasitol.-Drugs Drug Resist. 2019, 10, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.A.; El-Sayed, S.A.; El-Khodery, S.; Yokoyama, N.; Igarashi, I. Discovering the in vitro potent inhibitors against Babesia and Theileria parasites by repurposing the Malaria Box: A review. Vet. Parasitol. 2019, 274, 10. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.A.; El-Sayed, S.A.; AbouLaila, M.; Tuvshintulga, B.; Yokoyama, N.; Igarashi, I. Large-scale drug screening against Babesia divergens parasite using a fluorescence-based high-throughput screening assay. Vet. Parasitol. 2016, 227, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.A.; El-Sayed, S.A.; AbouLaila, M.; Yokoyama, N.; Igarashi, I. Evaluation of the inhibitory effect of N-acetyl-L-cysteine on Babesia and Theileria parasites. Exp. Parasitol. 2017, 179, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.; Beshbishy, A.M.; Tayebwa, D.S.; Adeyemi, O.S.; Shaheen, H.; Yokoyama, N.; Igarashi, I. The effects of trans-chalcone and chalcone 4 hydrate on the growth of Babesia and Theileria. PLoS Negl. Trop. Dis. 2019, 13, e0007030. [Google Scholar] [CrossRef]
- Batiha, G.E.; Beshbishy, A.M.; Alkazmi, L.M.; Nadwa, E.H.; Rashwan, E.K.; Yokoyama, N.; Igarashi, I. In vitro and in vivo growth inhibitory activities of cryptolepine hydrate against several Babesia species and Theileria equi. PLoS Negl. Trop. Dis. 2020, 14, 15. [Google Scholar] [CrossRef]
- Beshbishy, A.M.; Batiha, G.E.; Yokoyama, N.; Igarashi, I. Ellagic acid microspheres restrict the growth of Babesia and Theileria in vitro and Babesia microti in vivo. Parasit Vectors 2019, 12, 269. [Google Scholar] [CrossRef]
- Batiha, G.E.; Beshbishy, A.M.; Adeyemi, O.S.; Nadwa, E.; Rashwan, E.; Yokoyama, N.; Igarashi, I. Safety and efficacy of hydroxyurea and eflornithine against most blood parasites Babesia and Theileria. PLoS ONE 2020, 15, 15. [Google Scholar] [CrossRef]
- Salama, A.A.; AbouLaila, M.; Moussa, A.A.; Nayel, M.A.; Ei-Sify, A.; Terkawi, M.A.; Hassan, H.Y.; Yokoyama, N.; Igarashi, I. Evaluation of in vitro and in vivo inhibitory effects of fusidic acid on Babesia and Theileria parasites. Vet. Parasitol. 2013, 191, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.Y.; Luo, X.Y.; Wang, S.; He, L.; Zhao, J.L. Xanthohumol and Gossypol Are Promising Inhibitors against Babesia microti by In Vitro Culture via High-Throughput Screening of 133 Natural Products. Vaccines 2020, 8, 613. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.A.; El-Sayed, S.A.; Terkawi, M.A.; Youssef, M.A.; El Said, E.E.; Elsayed, G.; El-Khodery, S.; El-Ashker, M.; Elsify, A.; Omar, M.; et al. Optimization of a Fluorescence-Based Assay for Large-Scale Drug Screening against Babesia and Theileria Parasites. PLoS ONE 2015, 10, 15. [Google Scholar] [CrossRef]
- Munkhjargal, T.; AbouLaila, M.; Terkawi, M.A.; Sivakumar, T.; Ichikawa, M.; Davaasuren, B.; Nyamjargal, T.; Yokoyama, N.; Igarashi, I. Inhibitory Effects of Pepstatin A and Mefloquine on the Growth of Babesia Parasites. Am. J. Trop. Med. Hyg. 2012, 87, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.A.; AbouLaila, M.; El-Sayed, S.A.S.; Guswanto, A.; Yokoyama, N.; Igarashi, I. Inhibitory effects of fluoroquinolone antibiotics on Babesia divergens and Babesia microti, blood parasites of veterinary and zoonotic importance. Infect. Drug Resist. 2018, 11, 1605–1615. [Google Scholar] [CrossRef]
- Omar, M.A.; Salama, A.; Elsify, A.; Rizk, M.A.; Al-Aboody, M.S.; AbouLaila, M.; El-Sayed, S.A.; Igarashi, I. Evaluation of in vitro inhibitory effect of enoxacin on Babesia and Theileria parasites. Exp. Parasitol. 2016, 161, 62–67. [Google Scholar] [CrossRef]
- Batiha, G.E.S.; Tayebwa, D.S.; Beshbishy, A.M.; N’Da, D.D.; Yokoyama, N.; Igarashi, I. Inhibitory effects of novel ciprofloxacin derivatives on the growth of four Babesia species and Theileria equi. Parasitol. Res. 2020, 119, 3061–3073. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Beshbishy, A.M.; Tayebwa, D.S.; Shaheen, H.M.; Yokoyama, N.; Igarashi, I. Inhibitory effects of Syzygium aromaticum and Camellia sinensis methanolic extracts on the growth of Babesia and Theileria parasites. Ticks Tick-Borne Dis. 2019, 10, 949–958. [Google Scholar] [CrossRef]
- Batiha, G.E.; Beshbishy, A.M.; Guswanto, A.; Nugraha, A.; Munkhjargal, T.; M Abdel-Daim, M.; Mosqueda, J.; Igarashi, I. Phytochemical Characterization and Chemotherapeutic Potential of Cinnamomum verum Extracts on the Multiplication of Protozoan Parasites In Vitro and In Vivo. Molecules 2020, 25, 996. [Google Scholar] [CrossRef]
- Beshbishy, A.M.; Batiha, G.E.S.; Adeyemi, O.S.; Yokoyama, N.; Igarashi, I. Inhibitory effects of methanolic Olea europaea and acetonic Acacia laeta on growth of Babesia and Theileria. Asian Pac. J. Trop. Med. 2019, 12, 425–434. [Google Scholar] [CrossRef]
- Frampton, J.E. Tafenoquine: First Global Approval. Drugs 2018, 78, 1517–1523. [Google Scholar] [CrossRef]
- Carvalho, L.J.M.; Tuvshintulga, B.; Nugraha, A.B.; Sivakumar, T.; Yokoyama, N. Activities of artesunate-based combinations and tafenoquine against Babesia bovis in vitro and Babesia microti in vivo. Parasites Vectors 2020, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Ji, S.; Kondoh, D.; Galon, E.M.; Li, J.; Tomihari, M.; Yanagawa, M.; Tagawa, M.; Adachi, M.; Asada, M.; et al. Tafenoquine Is a Promising Drug Candidate for the Treatment of Babesiosis. Antimicrob. Agents Chemother. 2021, 65, e0020421. [Google Scholar] [CrossRef]
- Mordue, D.G.; Wormser, G.P. Could the Drug Tafenoquine Revolutionize Treatment of Babesia microti Infection? J. Infect. Dis. 2019, 220, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.S.; Hwang, J. Tafenoquine: A toxicity overview. Expert Opin. Drug Saf. 2021, 20, 349–362. [Google Scholar] [CrossRef]
- Rueangweerayut, R.; Bancone, G.; Harrell, E.J.; Beelen, A.P.; Kongpatanakul, S.; Möhrle, J.J.; Rousell, V.; Mohamed, K.; Qureshi, A.; Narayan, S.; et al. Hemolytic Potential of Tafenoquine in Female Volunteers Heterozygous for Glucose-6-Phosphate Dehydrogenase (G6PD) Deficiency (G6PD Mahidol Variant) versus G6PD-Normal Volunteers. Am. J. Trop. Med. Hyg 2017, 97, 702–711. [Google Scholar] [CrossRef]
- Chiu, J.E.; Renard, I.; George, S.; Pal, A.; Alday, H.; Narasimhan, S.; Riscoe, M.K.; Doggett, J.S.; Ben Mamoun, C. Cytochrome b Drug Resistance Mutation Decreases Babesia Fitness in the Tick Stages but not the Mammalian Erythrocytic Cycle. J. Infect. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Tuvshintulga, B.; Vannier, E.; Tayebwa, D.S.; Gantuya, S.; Sivakumar, T.; Guswanto, A.; Krause, P.J.; Yokoyama, N.; Igarashi, I. Clofazimine, a Promising Drug for the Treatment of Babesia microti Infection in Severely Immunocompromised Hosts. J. Infect. Dis. 2020, 222, 1027–1036. [Google Scholar] [CrossRef]
- Riccardi, N.; Giacomelli, A.; Canetti, D.; Comelli, A.; Intini, E.; Gaiera, G.; Diaw, M.M.; Udwadia, Z.; Besozzi, G.; Codecasa, L.; et al. Clofazimine: An old drug for never-ending diseases. Future Microbiol. 2020, 15, 557–566. [Google Scholar] [CrossRef]
- Jang, J.G.; Chung, J.H. Diagnosis and treatment of multidrug-resistant tuberculosis. Yeungnam Univ. J. Med. 2020, 37, 277–285. [Google Scholar] [CrossRef]
- Tuvshintulga, B.; AbouLaila, M.; Davaasuren, B.; Ishiyama, A.; Sivakumar, T.; Yokoyama, N.; Iwatsuki, M.; Otoguro, K.; Omura, S.; Igarashi, I. Clofazimine Inhibits the Growth of Babesia and Theileria Parasites In Vitro and In Vivo. Antimicrob. Agents Chemother. 2016, 60, 2739–2746. [Google Scholar] [CrossRef]
- Mitnick, C.D.; Shin, S.S.; Seung, K.J.; Rich, M.L.; Atwood, S.S.; Furin, J.J.; Fitzmaurice, G.M.; Viru, F.A.A.; Appleton, S.C.; Bayona, J.N.; et al. Comprehensive treatment of extensively drug-resistant tuberculosis. N. Engl. J. Med. 2008, 359, 563–574. [Google Scholar] [CrossRef]
- Frueh, L.; Li, Y.X.; Mather, M.W.; Li, Q.G.; Pou, S.; Nilsen, A.; Winter, R.W.; Forquer, I.P.; Pershing, A.M.; Xie, L.H.; et al. Alkoxycarbonate Ester Prodrugs of Preclinical Drug Candidate ELQ-300 for Prophylaxis and Treatment of Malaria. ACS Infect. Dis. 2017, 3, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Miley, G.P.; Pou, S.; Winter, R.; Nilsen, A.; Li, Y.X.; Kelly, J.X.; Stickles, A.M.; Mather, M.W.; Forquer, I.P.; Pershing, A.M.; et al. ELQ-300 Prodrugs for Enhanced Delivery and Single-Dose Cure of Malaria. Antimicrob. Agents Chemother. 2015, 59, 5555–5560. [Google Scholar] [CrossRef]
- Nilsen, A.; LaCrue, A.N.; White, K.L.; Forquer, I.P.; Cross, R.M.; Marfurt, J.; Mather, M.W.; Delves, M.J.; Shackleford, D.M.; Saenz, F.E.; et al. Quinolone-3-Diarylethers: A New Class of Antimalarial Drug. Sci. Transl. Med. 2013, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, A.; Miley, G.P.; Forquer, I.P.; Mather, M.W.; Katneni, K.; Li, Y.X.; Pou, S.; Pershing, A.M.; Stickles, A.M.; Ryan, E.; et al. Discovery, Synthesis, and Optimization of Antimalarial 4(1H)-Quinolone-3-Diarylethers. J. Med. Chem. 2014, 57, 3818–3834. [Google Scholar] [CrossRef]
- Smilkstein, M.J.; Pou, S.; Krollenbrock, A.; Bleyle, L.A.; Dodean, R.A.; Frueh, L.; Hinrichs, D.J.; Li, Y.X.; Martinson, T.; Munar, M.Y.; et al. ELQ-331 as a prototype for extremely durable chemoprotection against malaria. Malar. J. 2019, 18, 17. [Google Scholar] [CrossRef] [PubMed]
- Stickles, A.M.; de Almeida, M.J.; Morrisey, J.M.; Sheridan, K.A.; Forquer, I.P.; Nilsen, A.; Winter, R.W.; Burrows, J.N.; Fidock, D.A.; Vaidya, A.B.; et al. Subtle Changes in Endochin-Like Quinolone Structure Alter the Site of Inhibition within the Cytochrome bc(1) Complex of Plasmodium falciparum. Antimicrob. Agents Chemother. 2015, 59, 1977–1982. [Google Scholar] [CrossRef]
- Stickles, A.M.; Smilkstein, M.J.; Morrisey, J.M.; Li, Y.X.; Forquer, I.P.; Kelly, J.X.; Pou, S.; Winter, R.W.; Nilsen, A.; Vaidya, A.B.; et al. Atovaquone and ELQ-300 Combination Therapy as a Novel Dual-Site Cytochrome bc(1) Inhibition Strategy for Malaria. Antimicrob. Agents Chemother. 2016, 60, 4853–4859. [Google Scholar] [CrossRef] [PubMed]
- Stickles, A.M.; Ting, L.M.; Morrisey, J.M.; Li, Y.X.; Mather, M.W.; Meermeier, E.; Pershing, A.M.; Forquer, I.P.; Miley, G.P.; Pou, S.; et al. Inhibition of Cytochrome bc(1) as a Strategy for Single-Dose, Multi-Stage Antimalarial Therapy. Am. J. Trop. Med. Hyg. 2015, 92, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Van Schalkwyk, D.A.; Riscoe, M.K.; Pou, S.; Winter, R.W.; Nilsen, A.; Duffey, M.; Moon, R.W.; Sutherland, C.J. Novel Endochin-Like Quinolones Exhibit Potent In Vitro Activity against Plasmodium knowlesi but Do Not Synergize with Proguanil. Antimicrob. Agents Chemother. 2020, 64, 8. [Google Scholar] [CrossRef]
- Winter, R.; Kelly, J.X.; Smilkstein, M.J.; Hinrichs, D.; Koop, D.R.; Riscoe, M.K. Optimization of endochin-like quinolones for antimalarial activity. Exp. Parasitol. 2011, 127, 545–551. [Google Scholar] [CrossRef]
- Winter, R.W.; Kelly, J.X.; Smilkstein, M.J.; Dodean, R.; Hinrichs, D.; Riscoe, M.K. Antimalarial quinolones: Synthesis, potency, and mechanistic studies. Exp. Parasitol. 2008, 118, 487–497. [Google Scholar] [CrossRef]
- Doggett, J.S.; Nilsen, A.; Forquer, I.; Wegmann, K.W.; Jones-Brando, L.; Yolken, R.H.; Bordon, C.; Charman, S.A.; Katneni, K.; Schultz, T.; et al. Endochin-like quinolones are highly efficacious against acute and latent experimental toxoplasmosis. Proc. Natl. Acad. Sci. USA 2012, 109, 15936–15941. [Google Scholar] [CrossRef] [PubMed]
- Doggett, J.S.; Schultz, T.; Miller, A.J.; Bruzual, I.; Pou, S.; Winter, R.; Dodean, R.; Zakharov, L.N.; Nilsen, A.; Riscoe, M.K.; et al. Orally Bioavailable Endochin-Like Quinolone Carbonate Ester Prodrug Reduces Toxoplasma gondii Brain Cysts. Antimicrob. Agents Chemother. 2020, 64, 13. [Google Scholar] [CrossRef]
- Ortiz, D.; Forquer, I.; Boitz, J.; Soysa, R.; Elya, C.; Fulwiler, A.; Nilsen, A.; Polley, T.; Riscoe, M.K.; Ullman, B.; et al. Targeting the Cytochrome bc(1) Complex of Leishmania Parasites for Discovery of Novel Drugs. Antimicrob. Agents Chemother. 2016, 60, 4972–4982. [Google Scholar] [CrossRef] [PubMed]
- Alday, P.H.; Bruzual, I.; Nilsen, A.; Pou, S.; Winter, R.; Ben Mamoun, C.; Riscoe, M.K.; Doggett, J.S. Genetic Evidence for Cytochrome b Q(i) Site Inhibition by 4(1H)-Quinolone-3-Diarylethers and Antimycin in Toxoplasma gondii. Antimicrob. Agents Chemother. 2017, 61, 8. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Iorga, B.I.; Mounkoro, P.; Fisher, N.; Meunier, B. The antimalarial compound ELQ-400 is an unusual inhibitor of the bc(1) complex, targeting both Q(o) and Q(i) sites. FEBS Lett. 2018, 592, 1346–1356. [Google Scholar] [CrossRef] [PubMed]



| Babesia Species | In Vitro System | In Vivo Model |
|---|---|---|
| B. microti | Short-term ex vivo system [11,62] | Mice [63,64,65], hamsters [58,66,67], gerbils [51] |
| B. divergens | Continuous in vitro culture system in human RBCs [69] | Gerbils [74,75] |
| B. duncani | Continuous in vitro culture system in hamster [77,78] and human [79] RBCs | Mice [57,82,83], hamsters [80,81] |
| Drug | Treatment Regimen | Model | Effect | Ref. |
|---|---|---|---|---|
| ELQ-271 | 10 mg/kg (p.o.), 7 d | B. microti SCID mice | Parasitemia clearance followed by recrudescence by D12 post-treatment. | [11] |
| ELQ-316 | 10 mg/kg (p.o.), 7 d | B. microti SCID mice | Parasitemia clearance followed by recrudescence by D12 post-treatment. | [11] |
| ELQ-334 | 10 mg/kg (p.o.), 7 d | B. microti SCID mice | Parasitemia clearance followed by recrudescence by D16 post-treatment. | [11] |
| ELQ-334 + Atovaquone | 10 + 10 mg/kg (p.o.), 7 d | B. microti SCID mice | Parasitemia clearance throughout experiment. | [11] |
| ELQ-502 | 10 mg/kg (p.o.), 5 d | B. microti SCID mice | Parasitemia clearance followed by recrudescence by D17 post-treatment. | [109] |
| 10 mg/kg (p.o.), 10 d | B. microti SCID mice | Parasitemia clearance throughout study (DPI 91). | [57] | |
| 10 mg/kg (p.o.), 10 d | B. duncani C3H/HeJ mice | Parasitemia clearance throughout study (DPI 91). 100% survival. | [57] | |
| ELQ-502 + Atovaquone | 10 + 10 mg/kg (p.o.), 10 d | B. microti SCID mice | Parasitemia clearance throughout study (DPI 91). | [109] |
| 10 + 10 mg/kg (p.o.), 10 d | B. duncani C3H/HeJ mice | Parasitemia clearance throughout study (DPI 91). 100% survival | [109] | |
| Tafenoquine | 52 mg/kg (i.m.), 4 d (b.i.d.) | B. microti Golden hamsters | 100% suppression of parasitemia at D3 post-treatment. Reinfection of clean hamster negative. | [55] |
| 13 mg/kg (i.m.), 4 d (b.i.d.) | B. microti Golden hamsters | 99% suppression of parasitemia at D3 post-treatment. | [55] | |
| 3.25 mg/kg (i.m.), 4 d (b.i.d.) | B. microti Golden hamsters | 91% suppression of parasitemia at D3 post-treatment. | [55] | |
| 52 mg/kg (i.m.), 2 d (b.i.d.) | B. microti Golden hamsters | 99% suppression of parasitemia at D3 post-treatment. | [55] | |
| 20 mg/kg (p.o.), 1 d | B. microti SCID mice | Parasitemia clearance followed by recrudescence by D37 post-treatment. | [106] | |
| 25 mg/kg (p.o.), 1 d, + 12.5 mg/kg (p.o.), 1 d (4 d after 1st dose) | B. microti SCID mice | Parasitemia clear through D28 post-treatment. Reinfection of “clean” mice positive. | [106] | |
| 10 mg/kg (p.o.), 3 d | B. microti Balb/c mice | ~5.6× reduction in peak parasitemia. | [104] | |
| Clofazimine | 20 mg/kg (p.o.), 52 d | B. microti Balb/c mice | Parasitemia clear through DPI 90 (smear + PCR negative). | [110] |
| 20 mg/kg (p.o.), 7 d | B. microti Balb/c mice | Parasitemia clearance followed by recrudescence on DPI 26, unresponsive to a 2nd course of clofazimine 20 mg/kg (p.o.) (14 d). | [110] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renard, I.; Ben Mamoun, C. Treatment of Human Babesiosis: Then and Now. Pathogens 2021, 10, 1120. https://doi.org/10.3390/pathogens10091120
Renard I, Ben Mamoun C. Treatment of Human Babesiosis: Then and Now. Pathogens. 2021; 10(9):1120. https://doi.org/10.3390/pathogens10091120
Chicago/Turabian StyleRenard, Isaline, and Choukri Ben Mamoun. 2021. "Treatment of Human Babesiosis: Then and Now" Pathogens 10, no. 9: 1120. https://doi.org/10.3390/pathogens10091120
APA StyleRenard, I., & Ben Mamoun, C. (2021). Treatment of Human Babesiosis: Then and Now. Pathogens, 10(9), 1120. https://doi.org/10.3390/pathogens10091120