
Impact of Cytomegalovirus Infection and Genetic Background on the Frequencies of Peripheral Blood Suppressor Cells in Human Twins

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Basic Study Cohort Characteristics
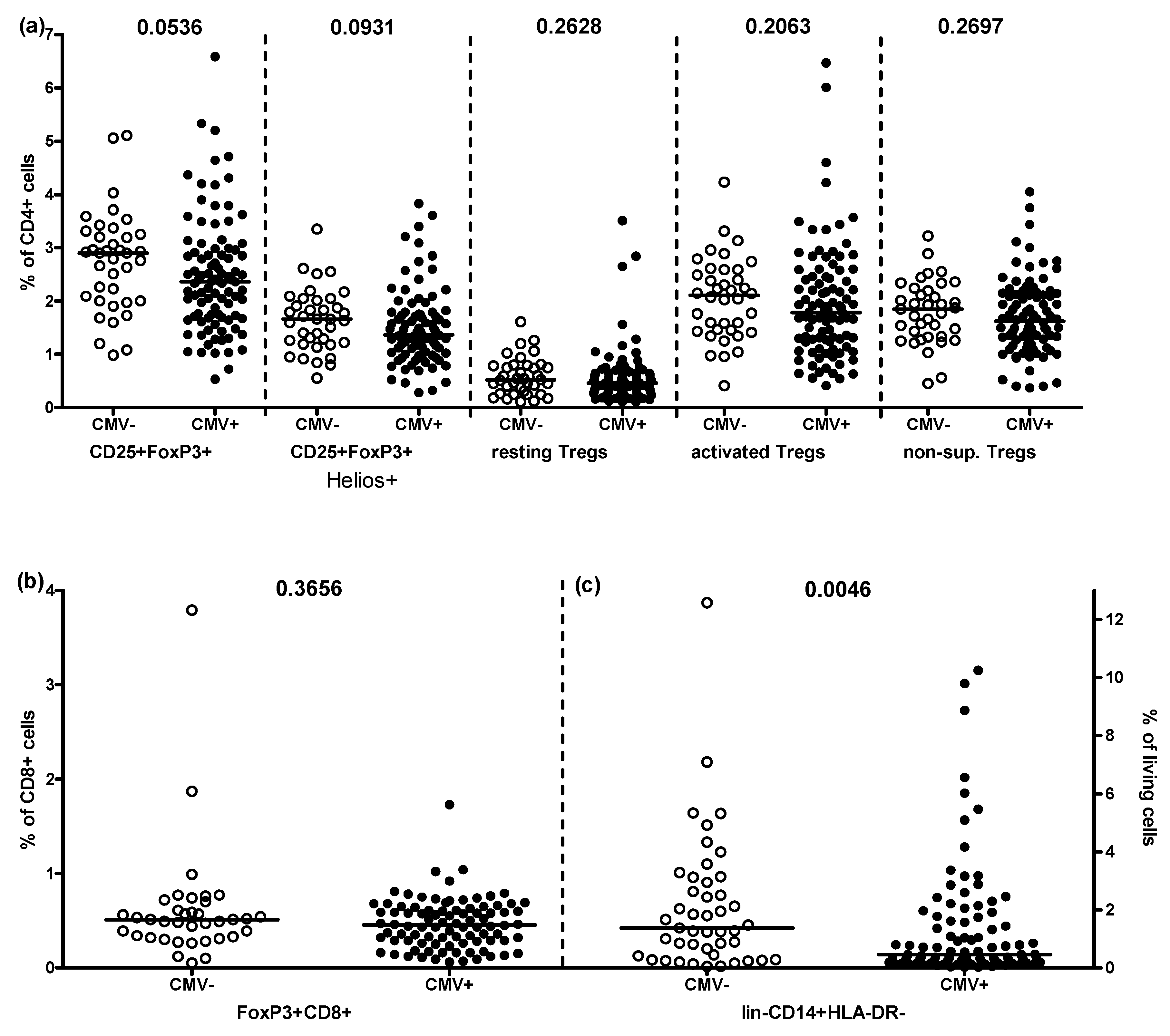
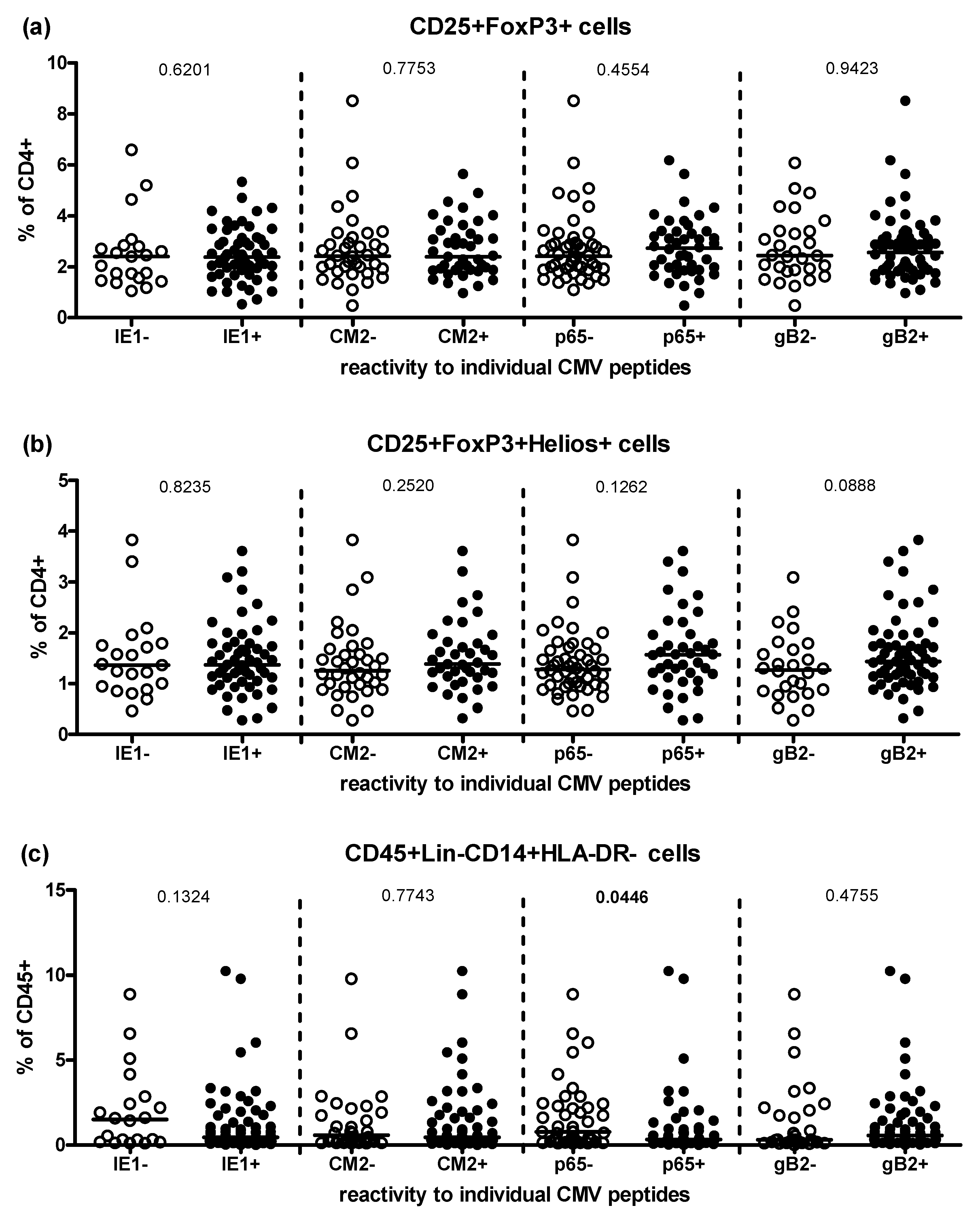
2.2. Effect of CMV-Seropositivity on Peripheral Treg and MDSC Frequencies
2.3. Correlations of Treg and MDSC Frequencies within Monozygotic and Dizygotic Twins
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Flow Cytometry
4.3. CMV-Serology
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adriaensen, W.; Derhovanessian, E.; Vaes, B.; Van Pottelbergh, G.; Degryse, J.M.; Pawelec, G.; Hamprecht, K.; Theeten, H.; Mathei, C. CD4:8 ratio >5 is associated with a dominant naive T-cell phenotype and impaired physical functioning in CMV-seropositive very elderly people: Results from the BELFRAIL study. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2015, 70, 143–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariza-Heredia, E.J.; Nesher, L.; Chemaly, R.F. Cytomegalovirus diseases after hematopoietic stem cell transplantation: A mini-review. Cancer Lett. 2014, 342, 1–8. [Google Scholar] [CrossRef]
- Barrett, L.; Fowke, K.R.; Grant, M.D. Cytomegalovirus, aging, and HIV: A perfect storm. AIDS Rev. 2012, 14, 159–167. [Google Scholar]
- Cantisan, S.; Torre-Cisneros, J.; Lara, R.; Zarraga, S.; Montejo, M.; Solana, R. Impact of cytomegalovirus on early immunosenescence of CD8+ T lymphocytes after solid organ transplantation. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2013, 68, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylwester, A.W.; Mitchell, B.L.; Edgar, J.B.; Taormina, C.; Pelte, C.; Ruchti, F.; Sleath, P.R.; Grabstein, K.H.; Hosken, N.A.; Kern, F.; et al. Broadly targeted human cytomegalovirus-specific CD4+ and CD8+ T cells dominate the memory compartments of exposed subjects. J. Exp. Med. 2005, 202, 673–685. [Google Scholar] [CrossRef] [Green Version]
- Derhovanessian, E.; Maier, A.B.; Beck, R.; Jahn, G.; Hahnel, K.; Slagboom, P.E.; de Craen, A.J.; Westendorp, R.G.; Pawelec, G. Hallmark features of immunosenescence are absent in familial longevity. J. Immunol. 2010, 185, 4618–4624. [Google Scholar] [CrossRef]
- Goldeck, D.; Larsen, L.A.; Christiansen, L.; Christensen, K.; Hamprecht, K.; Pawelec, G.; Derhovanessian, E. Genetic Influence on the Peripheral Blood CD4+ T-cell Differentiation Status in CMV Infection. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2016, 71, 1537–1543. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev. 2007, 128, 92–105. [Google Scholar] [CrossRef]
- Pawelec, G.; Bronikowski, A.; Cunnane, S.C.; Ferrucci, L.; Franceschi, C.; Fulop, T.; Gaudreau, P.; Gladyshev, V.N.; Gonos, E.S.; Gorbunova, V.; et al. The conundrum of human immune system “senescence”. Mech. Ageing Dev. 2020, 192, 111357. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafe, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Pawelec, G.; Picard, E.; Bueno, V.; Verschoor, C.P.; Ostrand-Rosenberg, S. MDSCs, ageing and inflammageing. Cell. Immunol. 2021, 362, 104297. [Google Scholar] [CrossRef]
- Bektas, A.; Schurman, S.H.; Sen, R.; Ferrucci, L. Human T cell immunosenescence and inflammation in aging. J. Leukoc. Biol. 2017, 102, 977–988. [Google Scholar] [CrossRef] [Green Version]
- Morrisette-Thomas, V.; Cohen, A.A.; Fulop, T.; Riesco, E.; Legault, V.; Li, Q.; Milot, E.; Dusseault-Belanger, F.; Ferrucci, L. Inflamm-aging does not simply reflect increases in pro-inflammatory markers. Mech. Ageing Dev. 2014, 139, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Pawelec, G. Aging as an inflammatory disease and possible reversal strategies. J. Allergy Clin. Immunol. 2020, 145, 1355–1356. [Google Scholar] [CrossRef] [Green Version]
- Moss, P. ‘From immunosenescence to immune modulation’: A re-appraisal of the role of cytomegalovirus as major regulator of human immune function. Med. Microbiol. Immunol. 2019, 208, 271–280. [Google Scholar] [CrossRef]
- Savva, G.M.; Pachnio, A.; Kaul, B.; Morgan, K.; Huppert, F.A.; Brayne, C.; Moss, P.A.; The Medical Research Council Cognitive Function and Ageing Study. Cytomegalovirus infection is associated with increased mortality in the older population. Aging Cell 2013, 12, 381–387. [Google Scholar] [CrossRef]
- Chen, S.; Pawelec, G.; Trompet, S.; Goldeck, D.; Mortensen, L.H.; Slagboom, P.E.; Christensen, K.; Gussekloo, J.; Kearney, P.; Buckley, B.M.; et al. Associations of Cytomegalovirus Infection with All-Cause and Cardiovascular Mortality in Multiple Observational Cohort Studies of Older Adults. J. Infect. Dis. 2021, 223, 238–246. [Google Scholar] [CrossRef]
- Brodin, P.; Jojic, V.; Gao, T.; Bhattacharya, S.; Angel, C.J.; Furman, D.; Shen-Orr, S.; Dekker, C.L.; Swan, G.E.; Butte, A.J.; et al. Variation in the human immune system is largely driven by non-heritable influences. Cell 2015, 160, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.; Maecker, H.T.; Brodin, P.; Nygaard, U.C.; Lyu, S.C.; Davis, M.M.; Nadeau, K.C.; Andorf, S. Aging and CMV discordance are associated with increased immune diversity between monozygotic twins. Immun. Ageing 2021, 18, 5. [Google Scholar] [CrossRef] [PubMed]
- Mangino, M.; Roederer, M.; Beddall, M.H.; Nestle, F.O.; Spector, T.D. Innate and adaptive immune traits are differentially affected by genetic and environmental factors. Nat. Commun. 2017, 8, 13850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, D.A.; Larsen, L.A.; Nygaard, M.; Mengel-From, J.; McGue, M.; Dalgard, C.; Hvidberg, L.; Hjelmborg, J.; Skytthe, A.; Holm, N.V.; et al. The Danish Twin Registry: An Updated Overview. Twin Res. Hum. Genet. 2019, 22, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, L.; Frederiksen, H.; Schousboe, K.; Skytthe, A.; von Wurmb-Schwark, N.; Christensen, K.; Kyvik, K. Age- and sex-differences in the validity of questionnaire-based zygosity in twins. Twin Res. 2003, 6, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Goldeck, D.; Adriaensen, W.; Oettinger, L.; Vaes, B.; van Pottelbergh, G.; Degryse, J.M.; Hamprecht, K.; Mathei, C.; Pawelec, G. Cellular immune phenotypes and worsening scores of frailty-associated parameters over an 18-month period in the very old. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2021. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Immune Subset | MZ Nonparametric 1 | MZ Linear R. 2 | DZ Nonparametroic 1 | DZ Linear R. 2 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sr | p Value | Outlier | r2 | p Value | Outlier | r2 | p Value | |||
| Tregs, 19 MZ and 17 DZ pair | ||||||||||
| CD25+ | 0.42 | 0.077 | 0 | 0.21 | 0.059 | 0.59 | 0.012 | 0 | 0.43 | 0.005 |
| FoxP3+ | 0.53 | 0.029 | 0 | 0.17 | 0.098 | 0.69 | 0.002 | 0 | 0.73 | <0.001 |
| CD25+FoxP3+ | 0.60 | 0.009 | 0 | 0.29 | 0.022 | 0.64 | 0.006 | 0 | 0.69 | <0.001 |
| Non suppressive Tregs | 0.46 | 0.062 | 0 | 0.24 | 0.045 | 0.635 | 0.006 | 0 | 0.38 | 0.008 |
| CD25+FoxP3+Helios+ | 0.40 | 0.11 | 0 | 0.18 | 0.092 | 0.62 | 0.008 | 0 | 0.58 | <0.001 |
| resting Tregs | 0.71 | 0.001 | 0 | 0.93 | <0.001 | −0.19 | 0.46 | 0 | 0.13 | 0.154 |
| activated Tregs | 0.55 | 0.014 | 0 | 0.60 | <0.001 | 0.58 | 0.014 | 0 | 0.57 | 0.001 |
| activated CD25+ Tregs | 0.51 | 0.004 | 0 | 0.21 | 0.063 | 0.64 | 0.006 | 0 | 0.57 | 0.001 |
| FoxP3-Helios+ | 0.78 | <0.001 | 0 | 0.59 | <0.001 | 0.64 | 0.005 | 0 | 0.20 | 0.075 |
| FoxP3+Helios+ | 0.28 | 0.271 | 0 | 0.20 | 0.076 | 0.66 | 0.004 | 0 | 0.71 | <0.001 |
| FoxP3+Helios- | 0.60 | 0.011 | 0 | 0.32 | 0.018 | 0.62 | 0.008 | 0 | 0.42 | 0.005 |
| CD45RA+CD45RO- | 0.82 | <0.001 | 0 | 0.71 | <0.001 | 0.28 | 0.277 | 0 | 0.11 | 0.199 |
| CD45RA+CD45RO-FoxP3+Helios+ | 0.31 | 0.233 | 0 | 0.75 | <0.001 | 0.05 | 0.0861 | 0 | <0.01 | 0.998 |
| CD45RA-CD45RO+FoxP3+Helios+ | 0.42 | 0.094 | 0 | 0.26 | 0.037 | 0.59 | 0.134 | 0 | 0.61 | <0.001 |
| CD8+FoxP3+ | 0.34 | 0.3448 | 1 | 0.11 | 0.193 | 0.69 | 0.002 | 0 | 0.39 | 0.007 |
| CD8+Helios+ | 0.23 | 0.374 | 0 | 0.09 | 0.252 | 0.14 | 0.59 | 0 | <0.01 | 0.915 |
| MDSC, 20 MZ and 13 DZ pairs | ||||||||||
| CD14+ | 0.51 | 0.021 | 0 | 0.33 | 0.008 | −0.23 | 0.448 | 0 | 0.01 | 0.720 |
| Lin- | 0.39 | 0.092 | 0 | 0.63 | <0.001 | 0.09 | 0.78 | 0 | 0.22 | 0.106 |
| Lin-CD14- | 0.53 | 0.016 | 0 | 0.65 | <0.001 | 0.31 | 0.306 | 0 | 0.31 | 0.047 |
| Lin-CD14-CD15+ | 0.34 | 0.1414 | 0 | 0.14 | 0.096 | 0.82 | 0.001 | 0 | 0.45 | 0.012 |
| Lin-CD14-CD15+CD11b+ | 0.14 | 0.707 | 0 | <0.01 | 0.876 | 0.49 | 0.216 | 0 | 0.33 | 0.140 |
| Lin-CD16-CD14+ | 0.45 | 0.047 | 0 | 0.38 | 0.004 | 0.10 | 0.734 | 0 | <0.01 | 0.846 |
| Lin-CD16-CD14+HLA-DR+ | 0.39 | 0.088 | 0 | 0.25 | 0.025 | −0.07 | 0.817 | 0 | <0.01 | 0.961 |
| Lin-CD16+CD14+ | 0.49 | 0.029 | 0 | 0.14 | 0.077 | 0.178 | 0.583 | 0 | 0.01 | 0.799 |
| Lin-CD16+CD14+HLA-DR+ | 0.49 | 0.030 | 0 | 0.19 | 0.056 | 0.07 | 0.831 | 0 | <0.01 | 0.952 |
| Lin-CD16+CD14low | 0.79 | <0.001 | 2 | 0.51 | 0.001 | 0.51 | 0.074 | 0 | 0.24 | 0.088 |
| Lin-CD16+CD14lowHLA-DR+ | 0.78 | <0.001 | 0 | 0.65 | <0.001 | 0.l15 | 0.629 | 0 | 0.01 | 0.751 |
| Lin-CD16-CD14- | 0.26 | 0.260 | 0 | 0.09 | 0.194 | 0.80 | 0.001 | 0 | 0.50 | 0.007 |
| Lin-CD14+HLA-DR- | 0.82 | <0.001 | 3 | 0.72 | <0.001 | 0.77 | 0.002 | 1 | 0.58 | 0.002 |
| Lin-CD14+HLA-DR+ | 0.43 | 0.056 | 0 | 0.20 | 0.047 | −0.13 | 0.668 | 0 | 0.01 | 0.762 |
| Lin-CD14-HLA-DR- | 0.70 | 0.001 | 0 | 0.45 | 0.001 | 0.10 | 0.748 | 0 | <0.01 | 0.897 |
| Lin-CD14-HLA-DR+ | 0.78 | <0.001 | 0 | 0.71 | <0.001 | 0.25 | 0.42 | 0 | 0.27 | 0.070 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goldeck, D.; Larsen, L.A.; Christensen, K.; Hamprecht, K.; Öttinger, L.; Hähnel, K.; Pawelec, G. Impact of Cytomegalovirus Infection and Genetic Background on the Frequencies of Peripheral Blood Suppressor Cells in Human Twins. Pathogens 2021, 10, 963. https://doi.org/10.3390/pathogens10080963
Goldeck D, Larsen LA, Christensen K, Hamprecht K, Öttinger L, Hähnel K, Pawelec G. Impact of Cytomegalovirus Infection and Genetic Background on the Frequencies of Peripheral Blood Suppressor Cells in Human Twins. Pathogens. 2021; 10(8):963. https://doi.org/10.3390/pathogens10080963
Chicago/Turabian StyleGoldeck, David, Lisbeth Aagaard Larsen, Kaare Christensen, Klaus Hamprecht, Lilly Öttinger, Karin Hähnel, and Graham Pawelec. 2021. "Impact of Cytomegalovirus Infection and Genetic Background on the Frequencies of Peripheral Blood Suppressor Cells in Human Twins" Pathogens 10, no. 8: 963. https://doi.org/10.3390/pathogens10080963
APA StyleGoldeck, D., Larsen, L. A., Christensen, K., Hamprecht, K., Öttinger, L., Hähnel, K., & Pawelec, G. (2021). Impact of Cytomegalovirus Infection and Genetic Background on the Frequencies of Peripheral Blood Suppressor Cells in Human Twins. Pathogens, 10(8), 963. https://doi.org/10.3390/pathogens10080963