
Molecular Diagnosis of Leishmaniasis: Quantification of Parasite Load by a Real-Time PCR Assay with High Sensitivity

 , ,
, , 
Abstract
1. Introduction
2. Results
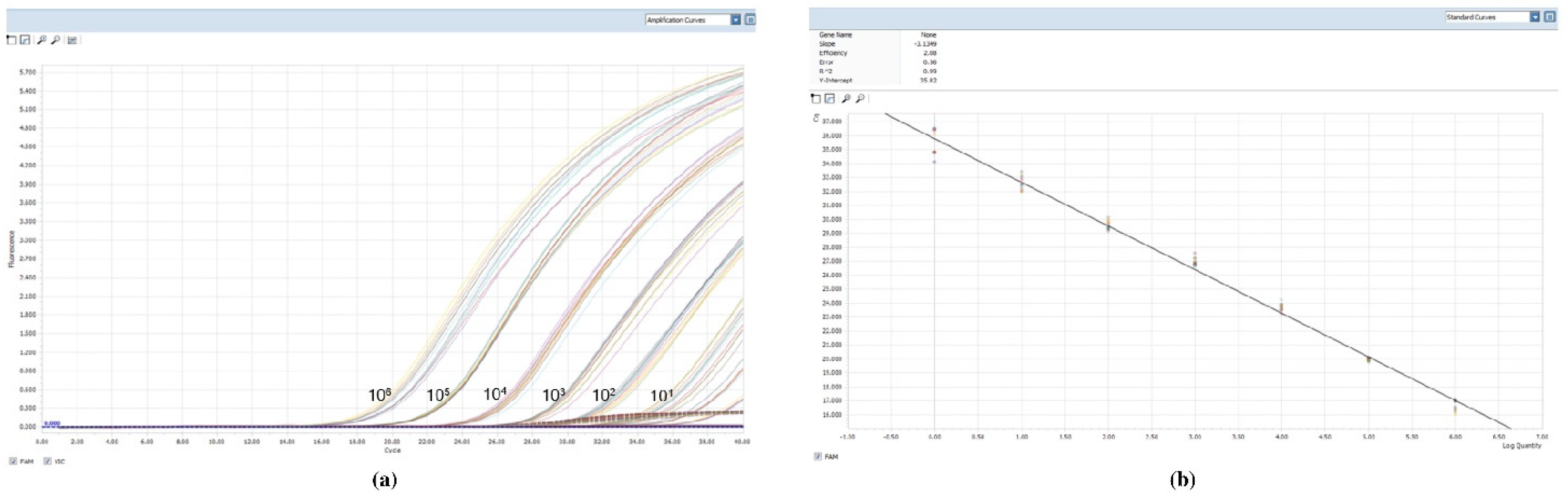
2.1. Sensitivity and Specificity of the Real-Time PCR Assay
2.2. Cloning and Correspondence Leishmania/mL-kDNA Copy Number
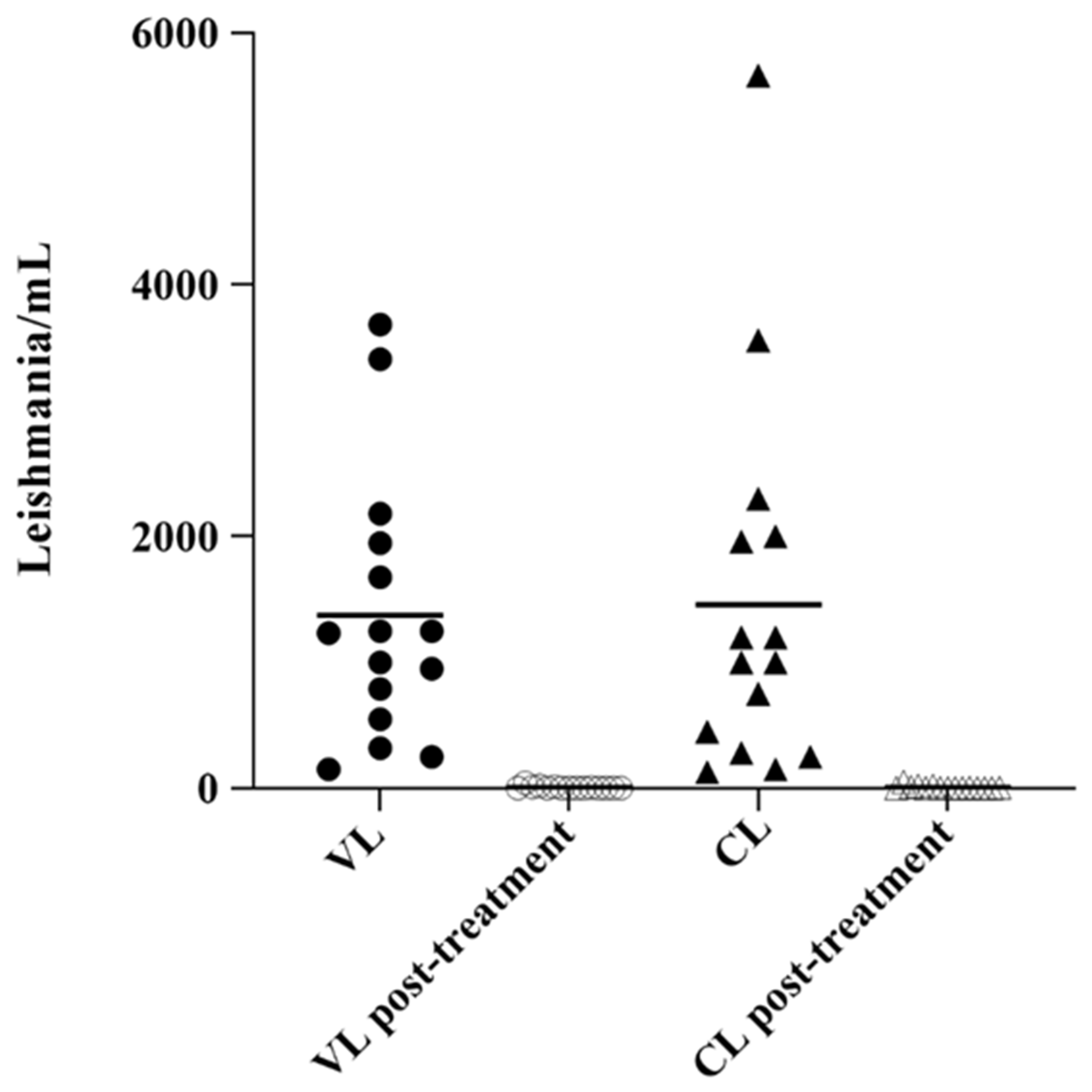
2.3. Analysis of Parasite Load in Clinical Samples
3. Discussion
4. Materials and Methods
4.1. Parasites
4.2. Clinical Samples
4.3. Conventional Diagnosis
4.4. DNA Extraction of Promastigote Cultures and Clinical Samples
4.5. Real-Time PCR
4.6. Cloning and Plasmid Generation
4.7. Clinical Sensitivity, Specificity & Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dedet, J.-P.; Pratlong, F. Leishmania, Trypanosoma and monoxenous trypanosomatids as emerging opportunistic agents. J. Eukaryot. Microbiol. 2000, 47, 37–39. [Google Scholar] [CrossRef] [PubMed]
- Baneth, G.; Aroch, I. Canine leishmaniasis: A diagnostic and clinical challenge. Vet. J. 2008, 175, 14–15. [Google Scholar] [CrossRef] [PubMed]
- Priolo, V.; Martínez-Orellana, P.; Pennisi, M.G.; Masucci, M.; Prandi, D.; Ippolito, D.; Bruno, F.; Castelli, G.; Solano-Gallego, L. Leishmania infantum-specific IFN-γ production in stimulated blood from cats living in areas where canine leishmaniosis is endemic. Parasit Vectors 2019, 12, 133. [Google Scholar] [CrossRef]
- Spada, E.; Perego, R.; Vitale, F.; Bruno, F.; Castelli, G.; Tarantola, G.; Baggiani, L.; Magistrelli, S.; Proverbio, D. Feline Leishmania spp. Infection in a Non-Endemic Area of Northern Italy. Animals 2020, 10, 817. [Google Scholar] [CrossRef]
- Baneth, G.; Thamsborg, S.M.; Otranto, D.; Guillot, J.; Blaga, R.; Deplazes, P.; Solano-Gallego, L. Major Parasitic Zoonoses Associated with Dogs and Cats in Europe. J. Comp. Pathol. 2016, 155, S54–S74. [Google Scholar] [CrossRef] [PubMed]
- Spada, E.; Canzi, I.; Baggiani, L.; Perego, R.; Vitale, F.; Migliazzo, A.; Proverbio, D. Prevalence of Leishmania infantum and co-infections in stray cats in northern Italy. Comp. Immunol. Microbiol. Infect. Dis. 2016, 45, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.A. Leishmaniasis, an Emerging Disease Found in Companion Animals in the United States. Top. Companion Anim. Med. 2009, 24, 182–188. [Google Scholar] [CrossRef]
- David, C.V.; Craft, N. Cutaneous and mucocutaneous leishmaniasis. Dermatol. Ther. 2009, 22, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Verso, M.G.; Vitale, F.; Castelli, G.; Bruno, F.; Migliazzo, A.; Bongiorno, M.R.; Santi, F.; Pistone, G.; Amodio, E.; Picciotto, D. Suspected cutaneous leishmaniasis in a sample of Westerns Sicily residents: What correlation with occupation? Med. Lav. Work Environ. Health 2017, 108, 123–129. [Google Scholar] [CrossRef]
- Sundar, S.; Rai, M. Laboratory Diagnosis of Visceral Leishmaniasis. Clin. Diagn. Lab. Immunol. 2002, 9, 951–958. [Google Scholar] [CrossRef]
- Adhya, S.; Hassan, M.Q.; Mukherjee, S.; Manna, P.P.; Basu, A.; Sen, S.; Bandyopadhyay, S. Visceral leishmaniasis in India: Promises and pitfalls of a PCR-based blood test. Trans. R. Soc. Trop. Med. Hyg. 2002, 96, S179–S183. [Google Scholar] [CrossRef]
- Adhya, S.; Chatterjee, M.; Hassan, M.Q.; Mukherjee, S.; Sen, S. Detection of Leishmania in the blood of early kala-azar patients with the aid of the polymerase chain reaction. Trans. R. Soc. Trop. Med. Hyg. 1995, 89, 622–624. [Google Scholar] [CrossRef]
- Lightner, L.K.; Chulay, J.D.; Bryceson, A.D. Comparison of microscopy and culture in the detection of Leishmania donovani from splenic aspirates. Am. J. Trop. Med. Hyg. 1983, 32, 296–299. [Google Scholar] [CrossRef] [PubMed]
- Massamba, N.N.; Mutinga, M.J. Recombinant kinetoplast DNA (kDNA) probe for identifying Leishmania tropica. Acta Trop. 1992, 52, 1–15. [Google Scholar] [CrossRef]
- Sudarshan, M.; Sundar, S. Parasite load estimation by qPCR differentiates between asymptomatic and symptomatic infection in Indian Visceral Leishmaniasis. Diagn. Microbiol. Infect. Dis. 2014, 80, 40–42. [Google Scholar] [CrossRef] [PubMed]
- Sundar, S.; Jha, T.K.; Thakur, C.P.; Engel, J.; Sindermann, H.; Fischer, C.; Junge, K.; Bryceson, A.; Berman, J. Oral Miltefosine for Indian Visceral Leishmaniasis. N. Engl. J. Med. 2002, 347, 1739–1746. [Google Scholar] [CrossRef] [PubMed]
- Sundar, S.; Singh, O.P. Molecular Diagnosis of Visceral Leishmaniasis. Mol. Diagn. Ther. 2018, 22, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.; Dayama, A.; Mehrotra, S.; Sundar, S. Diagnosis of visceral leishmaniasis. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Joshi, J.; Kaur, S. Leishmaniasis diagnosis: An update on the use of parasitological, immunological and molecular methods. J. Parasit. Dis. Off. Organ Indian Soc. Parasitol. 2020, 44, 253–272. [Google Scholar] [CrossRef]
- Reimão, J.Q.; Coser, E.M.; Lee, M.R.; Coelho, A.C. Laboratory Diagnosis of Cutaneous and Visceral Leishmaniasis: Current and Future Methods. Microorganisms 2020, 8, 1632. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Ceccarelli, M.; Diotallevi, A.; Menotta, M.; Magnani, M. Real-time PCR applications for diagnosis of leishmaniasis. Parasites Vectors 2018, 11, 273. [Google Scholar] [CrossRef]
- Paiva-Cavalcanti, M.; Silva de Morais, R.C.; Pessoa-e-Silva, R.; Trajano-Silva, L.A.; Gonçalves-de-Albuquerque, S.; Cavalcanti Tavares, D.; Brelaz-de-Castro, M.C.A.; de Freitas e Silva, R.; Pereira, V.R.A. Leishmaniases diagnosis: An update on the use of immunological and molecular tools. Cell Biosci. 2015, 5, 31. [Google Scholar] [CrossRef]
- Ahrberg, C.D.; Neužil, P. Doubling Throughput of a Real-Time PCR. Sci. Rep. 2015, 5, 12595. [Google Scholar] [CrossRef] [PubMed]
- Navarro, E.; Serrano-Heras, G.; Castaño, M.J.; Solera, J. Real-time PCR detection chemistry. Clin. Chim. Acta Int. J. Clin. Chem. 2015, 439, 231–250. [Google Scholar] [CrossRef]
- Quinnell, R.J.; Courtenay, O.; Davidson, S.; Garcez, L.; Lambson, B.; Ramos, P.; Shaw, J.J.; Shaw, M.A.; Dye, C. Detection of Leishmania infantum by PCR, serology and cellular immune response in a cohort study of Brazilian dogs. Parasitology 2001, 122, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Guarga, J.L.; Moreno, J.; Lucientes, J.; Gracia, M.J.; Peribáñez, M.A.; Alvar, J.; Castillo, J.A. Canine leishmaniasis transmission: Higher infectivity amongst naturally infected dogs to sand flies is associated with lower proportions of T helper cells. Res. Vet. Sci. 2000, 69, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Courtenay, O.; Quinnell, R.J.; Garcez, L.; Shaw, J.J.; Dye, C. Infectiousness in a cohort of brazilian dogs: Why culling fails to control visceral leishmaniasis in areas of high transmission. J. Infect. Dis. 2002, 186, 1314–1320. [Google Scholar] [CrossRef]
- Pennisi, M.G.; De Majo, M.; Masucci, M.; Britti, D.; Vitale, F.; Del Maso, R. Efficacy of the treatment of dogs with leishmaniosis with a combination of metronidazole and spiramycin. Vet. Rec. 2005, 156, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, M.G.; Reale, S.; Lo Giudice, S.; Masucci, M.; Caracappa, S.; Vitale, M.; Vitale, F. Real-time PCR in dogs treated for leishmaniasis with allopurinol. Vet. Res. Commun. 2005, 29 (Suppl. 2), 301–303. [Google Scholar] [CrossRef]
- Heid, C.A.; Stevens, S.; Livak, K.J.; Williams, P.M. Real time quantitative PCR. Genome Res. 1996, 6, 986–994. [Google Scholar] [CrossRef]
- Nicolas, L.; Prina, E.; Lang, T.; Milon, G. Real-Time PCR for Detection and Quantitation of Leishmania in Mouse Tissues. J. Clin. Microbiol. 2002, 40, 1666–1669. [Google Scholar] [CrossRef] [PubMed]
- Wittwer, C.T.; Ririe, K.M.; Andrew, R.V.; David, D.A.; Gundry, R.A.; Balis, U.J. The LightCycler: A microvolume multisample fluorimeter with rapid temperature control. BioTechniques 1997, 22, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Bretagne, S.; Durand, R.; Olivi, M.; Garin, J.F.; Sulahian, A.; Rivollet, D.; Vidaud, M.; Deniau, M. Real-time PCR as a new tool for quantifying Leishmania infantum in liver in infected mice. Clin. Diagn. Lab. Immunol. 2001, 8, 828–831. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schulz, A.; Mellenthin, K.; Schönian, G.; Fleischer, B.; Drosten, C. Detection, differentiation, and quantitation of pathogenic leishmania organisms by a fluorescence resonance energy transfer-based real-time PCR assay. J. Clin. Microbiol. 2003, 41, 1529–1535. [Google Scholar] [CrossRef] [PubMed]
- Peacock, C.S.; Seeger, K.; Harris, D.; Murphy, L.; Ruiz, J.C.; Quail, M.A.; Peters, N.; Adlem, E.; Tivey, A.; Aslett, M.; et al. Comparative genomic analysis of three Leishmania species that cause diverse human disease. Nat. Genet. 2007, 39, 839–847. [Google Scholar] [CrossRef]
- Smith, D.F.; Peacock, C.S.; Cruzc, A. Comparative genomics: From genotype to disease phenotype in the leishmaniases. Int. J. Parasitol. 2007, 37, 1173–1186. [Google Scholar] [CrossRef] [PubMed]
- Weirather, J.L.; Jeronimo, S.M.B.; Gautam, S.; Sundar, S.; Kang, M.; Kurtz, M.A.; Haque, R.; Schriefer, A.; Talhari, S.; Carvalho, E.M.; et al. Serial Quantitative PCR Assay for Detection, Species Discrimination, and Quantification of Leishmania spp. in Human Samples. J. Clin. Microbiol. 2011, 49, 3892–3904. [Google Scholar] [CrossRef] [PubMed]
- Urrea, D.A.; Triana-Chavez, O.; Alzate, J.F. Mitochondrial genomics of human pathogenic parasite Leishmania (Viannia) panamensis. PeerJ 2019, 7, e7235. [Google Scholar] [CrossRef]
- Ceccarelli, M.; Buffi, G.; Diotallevi, A.; Andreoni, F.; Bencardino, D.; Vitale, F.; Castelli, G.; Bruno, F.; Magnani, M.; Galluzzi, L. Evaluation of a KDNA-Based QPCR Assay for the Detection and Quantification of Old World Leishmania Species. Microorganisms 2020, 8, 2006. [Google Scholar] [CrossRef] [PubMed]
- Vitale, F.; Reale, S.; Vitale, M.; Petrotta, E.; Torina, A.; Caracappa, S. TaqMan-Based Detection of Leishmania infantum DNA Using Canine Samples. Ann. N. Y. Acad. Sci. 2004, 1026, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Manna, L.; Reale, S.; Vitale, F.; Picillo, E.; Pavone, L.M.; Gravino, A.E. Real-time PCR assay in Leishmania-infected dogs treated with meglumine antimoniate and allopurinol. Vet. J. 2008, 177, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Pita-Pereira, D.; Lins, R.; Oliveira, M.P.; Lima, R.B.; Pereira, B.A.S.; Moreira, O.C.; Brazil, R.P.; Britto, C. SYBR Green-based Real-Time PCR targeting kinetoplast DNA can be used to discriminate between the main etiologic agents of Brazilian cutaneous and visceral leishmaniases. Parasit Vectors 2012, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Bertozzini, E.; Penna, A.; Perini, F.; Pigalarga, A.; Graneli, E.; Magnani, M. Detection and quantification of Prymnesium parvum (Haptophyceae) by real-time PCR. Lett. Appl. Microbiol. 2008, 46, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Mackay, I.M.; Arden, K.E.; Nitsche, A. Real-time PCR in virology. Nucleic Acids Res. 2002, 30, 1292–1305. [Google Scholar] [CrossRef] [PubMed]
- Lukeš, J.; Wheeler, R.; Jirsová, D.; David, V.; Archibald, J.M. Massive mitochondrial DNA content in diplonemid and kinetoplastid protists. IUBMB Life 2018, 70, 1267–1274. [Google Scholar] [CrossRef]
- Akhoundi, M.; Downing, T.; Votýpka, J.; Kuhls, K.; Lukeš, J.; Cannet, A.; Ravel, C.; Marty, P.; Delaunay, P.; Kasbari, M.; et al. Leishmania infections: Molecular targets and diagnosis. Mol. Aspects Med. 2017, 57, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, M.; Galluzzi, L.; Diotallevi, A.; Andreoni, F.; Fowler, H.; Petersen, C.; Vitale, F.; Magnani, M. The use of kDNA minicircle subclass relative abundance to differentiate between Leishmania (L.) infantum and Leishmania (L.) amazonensis. Parasit Vectors 2017, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, M.; Galluzzi, L.; Migliazzo, A.; Magnani, M. Detection and Characterization of Leishmania (Leishmania) and Leishmania (Viannia) by SYBR Green-Based Real-Time PCR and High Resolution Melt Analysis Targeting Kinetoplast Minicircle DNA. PLoS ONE 2014, 9, e88845. [Google Scholar] [CrossRef] [PubMed]
- Morganti, G.; Veronesi, F.; Stefanetti, V.; Di Muccio, T.; Fiorentino, E.; Diaferia, M.; Santoro, A.; Passamonti, F.; Gramiccia, M. Emerging feline vector-borne pathogens in Italy. Parasit Vectors 2019, 12, 193. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, S.; Hejazi, S.H.; Hashemzadeh, M.; Eslami, G.; Darani, H.Y. Molecular diagnosis of Old World leishmaniasis: Real-time PCR based on tryparedoxin peroxidase gene for the detection and identification of Leishmania spp. J. Vector Borne Dis. 2012, 49, 15–18. [Google Scholar] [PubMed]
- El-Beshbishy, H.A.; Al-Ali, K.; El-Badry, A.A. Molecular characterization of cutaneous leishmaniasis in Al-Madinah Al-Munawarah Province, western Saudi Arabia. Int. J. Infect. Dis. IJID 2013, 17, 334–E338. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kevric, I.; Cappel, M.A.; Keeling, J.H. New World and Old World Leishmania Infections: A Practical Review. Dermatol. Clin. 2015, 33, 579–593. [Google Scholar] [CrossRef]
- Disch, J.; Pedras, M.J.; Orsini, M.; Pirmez, C.; de Oliveira, M.C.; Castro, M.; Rabello, A. Leishmania (Viannia) subgenus kDNA amplification for the diagnosis of mucosal leishmaniasis. Diagn. Microbiol. Infect. Dis. 2005, 51, 185–190. [Google Scholar] [CrossRef]
- Belinchón-Lorenzo, S.; Iniesta, V.; Parejo, J.C.; Fernández-Cotrina, J.; Muñoz-Madrid, R.; Soto, M.; Alonso, C.; Gómez Nieto, L.C. Detection of Leishmania infantum kinetoplast minicircle DNA by Real Time PCR in hair of dogs with leishmaniosis. Vet. Parasitol. 2013, 192, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L. Kinetoplast DNA in Trypanosomid Flagellates. Int. Rev. Cytol. 1986, 99, 119–179. [Google Scholar] [CrossRef] [PubMed]
- Castelli, G.; Bruno, F.; Caputo, V.; Fiorella, S.; Sammarco, I.; Lupo, T.; Migliazzo, A.; Vitale, F.; Reale, S. Genetic tools discriminate strains of Leishmania infantum isolated from humans and dogs in Sicily, Italy. PLoS Negl. Trop. Dis. 2020, 14, e0008465. [Google Scholar] [CrossRef] [PubMed]
- Mesa, L.E.; Manrique, R.; Muskus, C.; Robledo, S.M. Test accuracy of polymerase chain reaction methods against conventional diagnostic techniques for Cutaneous Leishmaniasis (CL) in patients with clinical or epidemiological suspicion of CL: Systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2020, 14, e0007981. [Google Scholar] [CrossRef] [PubMed]
- Bensoussan, E.; Nasereddin, A.; Jonas, F.; Schnur, L.F.; Jaffe, C.L. Comparison of PCR Assays for Diagnosis of Cutaneous Leishmaniasis. J. Clin. Microbiol. 2006, 44, 1435–1439. [Google Scholar] [CrossRef] [PubMed]
- Aviles, H.; Belli, A.; Armijos, R.; Monroy, F.P.; Harris, E. PCR detection and identification of Leishmania parasites in clinical specimens in Ecuador: A comparison with classical diagnostic methods. J. Parasitol. 1999, 85, 181–187. [Google Scholar] [CrossRef]
- Medeiros, A.C.R.; Rodrigues, S.S.; Roselino AM, F. Comparison of the specificity of PCR and the histopathological detection of leishmania for the diagnosis of American cutaneous leishmaniasis. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Med. Biol. 2002, 35, 421–424. [Google Scholar] [CrossRef]
- Rodríguez, N.; Guzman, B.; Rodas, A.; Takiff, H.; Bloom, B.R.; Convit, J. Diagnosis of cutaneous leishmaniasis and species discrimination of parasites by PCR and hybridization. J. Clin. Microbiol. 1994, 32, 2246–2252. [Google Scholar] [CrossRef]
- Safaei, A.; Motazedian, M.H.; Vasei, M. Polymerase chain reaction for diagnosis of cutaneous leishmaniasis in histologically positive, suspicious and negative skin biopsies. Dermatology 2002, 205, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Andresen, K.; Gaafar, A.; El-Hassan, A.M.; Ismail, A.; Dafalla, M.; Theander, T.G.; Kharazmi, A. Evaluation of the polymerase chain reaction in the diagnosis of cutaneous leishmaniasis due to Leishmania major: A comparison with direct microscopy of smears and sections from lesions. Trans. R. Soc. Trop. Med. Hyg. 1996, 90, 133–135. [Google Scholar] [CrossRef]
- Laskay, T.; Mikó, T.L.; Negesse, Y.; Solbach, W.; Röllinghoff, M.; Frommel, D. Detection of cutaneous Leishmania infection in paraffin-embedded skin biopsies using the polymerase chain reaction. Trans. R. Soc. Trop. Med. Hyg. 1995, 89, 273–275. [Google Scholar] [CrossRef]
- Ashford, D.A.; Bozza, M.; Freire, M.; Miranda, J.C.; Italo Sherlock, I.; Eulalio, C.; Lopes, U.; Fernandes, O.; Degrave, W.; Barker, R.H., Jr.; et al. Comparison of the polymerase chain reaction and serology for the detection of canine visceral leishmaniasis. Am. J. Trop. Med. Hyg. 1995, 53, 251–255. [Google Scholar] [CrossRef]
- Adel, A.; Berkvens, D.; Abatih, E.; Soukehal, A.; Bianchini, J.; Saegerman, C. Evaluation of Immunofluorescence Antibody Test Used for the Diagnosis of Canine Leishmaniasis in the Mediterranean Basin: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0161051. [Google Scholar] [CrossRef]
- Mancianti, F.; Meciani, N. Specific serodiagnosis of canine leishmaniasis by indirect immunofluorescence, indirect hemagglutination, and counterimmunoelectrophoresis. Am. J. Vet. Res. 1988, 49, 1409–1411. [Google Scholar]
- Leishmaniosis. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.11_LEISHMANIOSIS.pdf (accessed on 26 March 2020).
- Van Griensven, J.; Carrillo, E.; López-Vélez, R.; Lynen, L.; Moreno, J. Leishmaniasis in immunosuppressed individuals. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2014, 20, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Urbani, L.; Tirolo, A.; Daniela Salvatore, D.; Tumbarello, M.; Segatore, S.; Battilani, M.; Balboni, A.; Dondi, F. Serological, molecular and clinicopathological findings associated with Leishmania infantum infection in cats in Northern Italy. J. Feline Med. Surg. 2020, 22, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, M.-G.; Cardoso, L.; Baneth, G.; Bourdeau, P.; Koutinas, A.; Miró, G.; Oliva, G.; Solano-Gallego, L. LeishVet update and recommendations on feline leishmaniosis. Parasit Vectors 2015, 8, 302. [Google Scholar] [CrossRef] [PubMed]
- Daprà, F.; Scalone, A.; Mignone, W.; Ferroglio, E.; Mannelli, A.; Biglino, A.; Zanatta, R.; Gradoni, L.; Rosati, S. Validation of a Recombinant Based Antibody ELISA for Diagnosis of Human and Canine Leishmaniasis. J. Immunoassay Immunochem. 2008, 29, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Boarino, A.; Bollo, E.; Prunotto, L.; Canale, L.; Uslenghi, F.; Poletti, P. Application of a recombinant protein for the serological diagnosis of canine leishmaniasis. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Reale, S.; Maxia, L.; Vitale, F.; Glorioso, N.S.; Caracappa, S.; Vesco, G. Detection of Leishmania infantum in Dogs by PCR with Lymph Node Aspirates and Blood. J. Clin. Microbiol. 1999, 37, 2931–2935. [Google Scholar]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A method for the absolute quantification of cDNA using real-time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef]
- Martínez, V.; Quilez, J.; Sanchez, A.; Roura, X.; Francino, O.; Altet, L. Canine leishmaniasis: The key points for qPCR result interpretation. Parasites Vectors 2011, 4, 57. [Google Scholar] [CrossRef]
- Castelli, G.; Galante, A.; Lo Verde, V.; Migliazzo, A.; Reale, S.; Lupo, T.; Piazza, M.; Vitale, F.; Bruno, F. Evaluation of two modified culture media for Leishmania infantum cultivation versus different culture media. J. Parasitol. 2014, 100, 228–230. [Google Scholar] [CrossRef] [PubMed]
- Hossain, F.; Ghosh, P.; Khan, M.A.A.; Duthie, M.S.; Vallur, A.C.; Picone, A.; Howard, R.F.; Reed, S.G.; Mondal, D. Real-time PCR in detection and quantitation of Leishmania donovani for the diagnosis of Visceral Leishmaniasis patients and the monitoring of their response to treatment. PLoS ONE 2017, 12, e0185606. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Clinical Presentation | Conventional Diagnosis (IFAT, PC, MH) | Real-Time PCR (qPCR) kDNA | |||
|---|---|---|---|---|---|
| Clinical Samples | IFAT Positive/Total (%) | PC Positive/Total (%) | MH Positive/Total (%) | Positive/Total (%) | |
| VL | Peripheral blood/serum | 13/15 (86.6) | 4/15 (26.6) | 10/15 (66.6) | 13/15 (86.6) |
| Bone marrow | n.a. | 8/15 (53.3) | 13/15 (86.6) | 15/15 (100) | |
| CL | Tissue/skin biopsy | n.a. | 9/15 (60) | 10/15 (66.6) | 15/15 (100) |
| CanL | Peripheral blood/serum | 19/30 (63.3) | 5/30 (16.6) | 5/30 (16.6) | 19/30 (63.3) |
| Popliteal lymph node | n.a. | 16/30 (53.3) | 17/30 (56.6) | 30/30 (100) | |
| FeL | Peripheral blood/serum | 8/20 (40) | 0/20 (0) | 0/20 (0) | 1/20 (5) |
| Popliteal lymph node | n.a. | 1/20 (5) | 1/20 (5) | 20/20 (100) | |
| Conventional Methods | Molecular Method | ||||||
|---|---|---|---|---|---|---|---|
| Species | Clinical Status | Sample ID | IFAT Titer | Parasite Culture | Microscopy-Histology | qPCR Blood (Leishmania/mL) | qPCR Lymph Node (Leishmania/mL) |
| Canine | Symptomatic | 1 | 1:160 | Blood − Lymph node − | Blood − Lymph node − | 45 | 540 |
| 2 | 1:5120 | Blood + Lymph node + | Blood + Lymph node + | 590 | 15,680 | ||
| 3 | 1:640 | Blood − Lymph node + | Blood − Lymph node + | 30 | 970 | ||
| 4 | 1:1280 | Blood − Lymph node + | Blood − Lymph node + | 120 | 7838 | ||
| 5 | 1:320 | Blood − Lymph node + | Blood − Lymph node + | 75 | 660 | ||
| 6 | 1:2560 | Blood − Lymph node + | Blood − Lymph node + | 60 | 9275 | ||
| 7 | Negative | Blood − Lymph node − | Blood − Lymph node − | Negative | 50 | ||
| 8 | 1:80 | Blood − Lymph node − | Blood − Lymph node − | Negative | 100 | ||
| 9 | 1:5120 | Blood + lymph node + | Blood + Lymph node + | 645 | 24,890 | ||
| 10 | 1:160 | Blood − Lymph node + | Blood − Lymph node + | 10 | 390 | ||
| 11 | 1:160 | Blood − Lymph node + | Blood − Lymph node + | 20 | 460 | ||
| 12 | 1:640 | Blood − Lymph node + | Blood − Lymph node + | 285 | 5300 | ||
| 13 | 1:320 | Blood − Lymph node + | Blood − Lymph node + | 230 | 910 | ||
| 14 | 1:2560 | Blood − lymph node + | Blood − Lymph node + | 185 | 19,020 | ||
| 15 | 1:2560 | Blood + lymph node + | Blood+ Lymph node + | 580 | 17,535 | ||
| 16 | 1:320 | Blood − Lymph node − | Blood − Lymph node + | 150 | 4230 | ||
| 17 | 1:1280 | Blood − Lymph node + | Blood − Lymph node + | 320 | 12,545 | ||
| 18 | 1:160 | Blood − Lymph node − | Blood − Lymph node − | 35 | 850 | ||
| 19 | 1:5120 | Blood + lymph node + | Blood + Lymph node + | 495 | 1,000,000 | ||
| 20 | 1:1280 | Blood + Lymph node + | Blood + Lymph node + | 320 | 6710 | ||
| Asymptomatic | 21 | Negative | Blood − Lymph node − | Blood − Lymph node − | Negative | 10 | |
| 22 | 1:40 | Blood – Lymph node − | Blood − Lymph node − | Negative | 20 | ||
| 23 | 1:160 | Blood − Lymph node + | Blood − Lymph node − | 10 | 120 | ||
| 24 | 1:40 | Blood − Lymph node − | Blood − Lymph node − | Negative | 15 | ||
| 25 | 1:80 | Blood − Lymph node − | Blood − Lymph node − | Negative | 20 | ||
| 26 | Negative | Blood − Lymph node − | Blood − Lymph node − | Negative | 15 | ||
| 27 | 1:40 | Blood − Lymph node − | Blood − Lymph node − | Negative | 30 | ||
| 28 | 1:40 | Blood − Lymph node − | Blood − Lymph node − | Negative | 65 | ||
| 29 | 1:80 | Blood − Lymph node − | Blood − Lymph node − | Negative | 80 | ||
| 30 | 1:80 | Blood − Lymph node − | Blood − Lymph node − | Negative | 35 | ||
| Feline | Symptomatic | 31 | 1:80 | Blood − Lymph node − | Blood − Lymph node − | Negative | 5 |
| 32 | 1:80 | Blood − Lymph node − | Blood − Lymph node − | Negative | 10 | ||
| 33 | 1:160 | Blood − Lymph node − | Blood − Lymph node − | Negative | 20 | ||
| 34 | 1:160 | Blood − Lymph node − | Blood − Lymph node − | Negative | 5 | ||
| 35 | Negative | Blood − Lymph node − | Blood − Lymph node − | Negative | 10 | ||
| 36 | 1:640 | Blood − Lymph node + | Blood − Lymph node + | 250 | 980 | ||
| 37 | Negative | Blood − Lymph node − | Blood − Lymph node − | Negative | 20 | ||
| 38 | Negative | Blood − Lymph node − | Blood − Lymph node − | Negative | 25 | ||
| 39 | 1:80 | Blood − Lymph node − | Blood − Lymph node − | Negative | 30 | ||
| 40 | 1:160 | Blood − Lymph node − | Blood − Lymph node − | Negative | 55 | ||
| 41 | 1:320 | Blood − Lymph node − | Blood − Lymph node + | 10 | 150 | ||
| 42 | 1:80 | Blood − Lymph node − | Blood − Lymph node − | Negative | 15 | ||
| 43 | 1:80 | Blood − Lymph node − | Blood − Lymph node − | Negative | 10 | ||
| 44 | 1:160 | Blood − Lymph node − | Blood − Lymph node − | Negative | 35 | ||
| 45 | 1:80 | Blood − Lymph node − | Blood − Lymph node − | Negative | 10 | ||
| 46 | Negative | Blood − Lymph node − | Blood − Lymph node − | Negative | 5 | ||
| 47 | 1:160 | Blood − Lymph node − | Blood − Lymph node − | Negative | 50 | ||
| 48 | Negative | Blood − Lymph node − | Blood − Lymph node − | Negative | 5 | ||
| 49 | 1:320 | Blood − Lymph node − | Blood − Lymph node − | 5 | 350 | ||
| 50 | Negative | Blood − Lymph node − | Blood − Lymph node − | Negative | 60 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castelli, G.; Bruno, F.; Reale, S.; Catanzaro, S.; Valenza, V.; Vitale, F. Molecular Diagnosis of Leishmaniasis: Quantification of Parasite Load by a Real-Time PCR Assay with High Sensitivity. Pathogens 2021, 10, 865. https://doi.org/10.3390/pathogens10070865
Castelli G, Bruno F, Reale S, Catanzaro S, Valenza V, Vitale F. Molecular Diagnosis of Leishmaniasis: Quantification of Parasite Load by a Real-Time PCR Assay with High Sensitivity. Pathogens. 2021; 10(7):865. https://doi.org/10.3390/pathogens10070865
Chicago/Turabian StyleCastelli, Germano, Federica Bruno, Stefano Reale, Simone Catanzaro, Viviana Valenza, and Fabrizio Vitale. 2021. "Molecular Diagnosis of Leishmaniasis: Quantification of Parasite Load by a Real-Time PCR Assay with High Sensitivity" Pathogens 10, no. 7: 865. https://doi.org/10.3390/pathogens10070865
APA StyleCastelli, G., Bruno, F., Reale, S., Catanzaro, S., Valenza, V., & Vitale, F. (2021). Molecular Diagnosis of Leishmaniasis: Quantification of Parasite Load by a Real-Time PCR Assay with High Sensitivity. Pathogens, 10(7), 865. https://doi.org/10.3390/pathogens10070865