
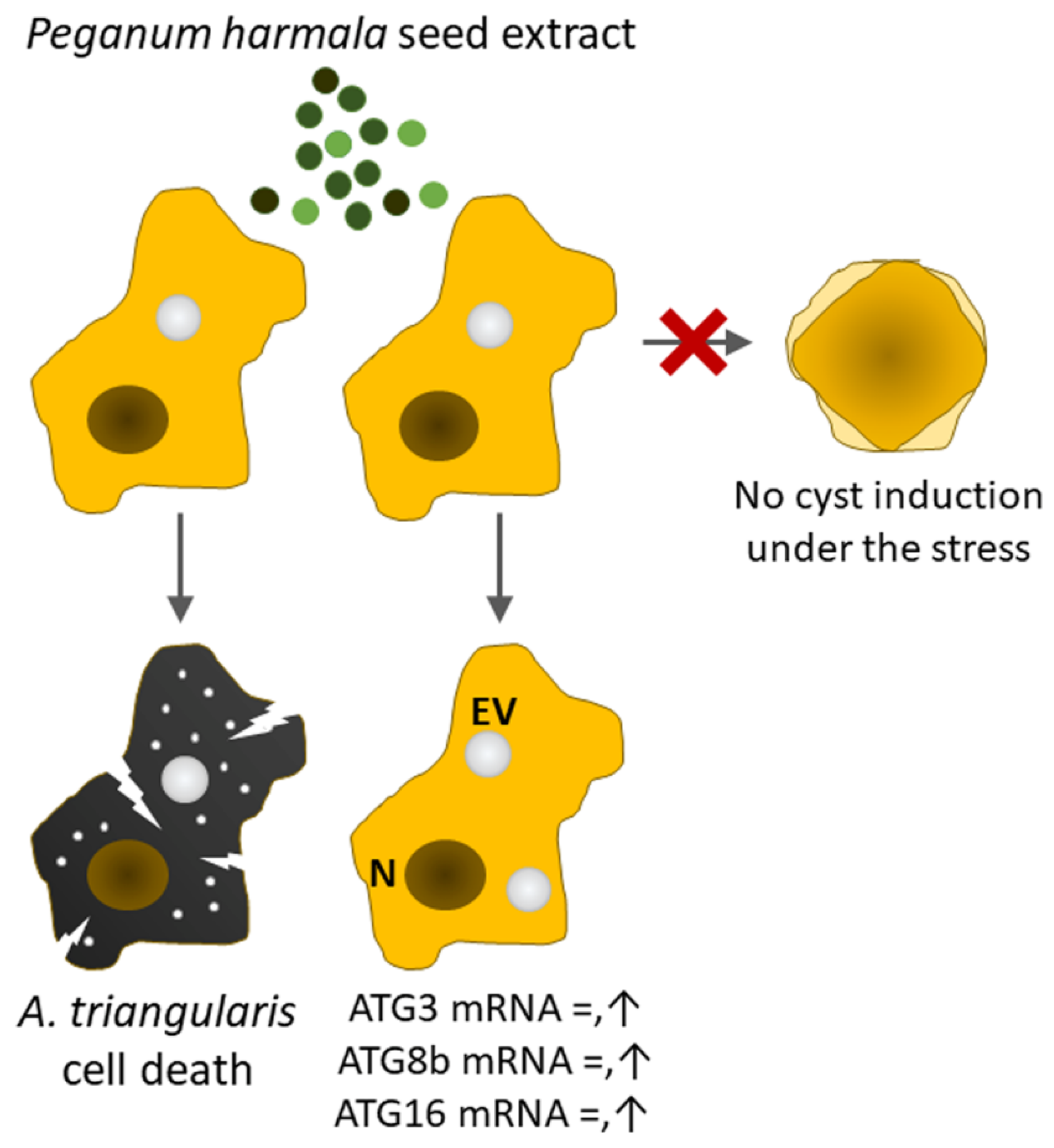
Peganum harmala Extract Has Antiamoebic Activity to Acanthamoeba triangularis Trophozoites and Changes Expression of Autophagy-Related Genes


 ,
,  ,
,  , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
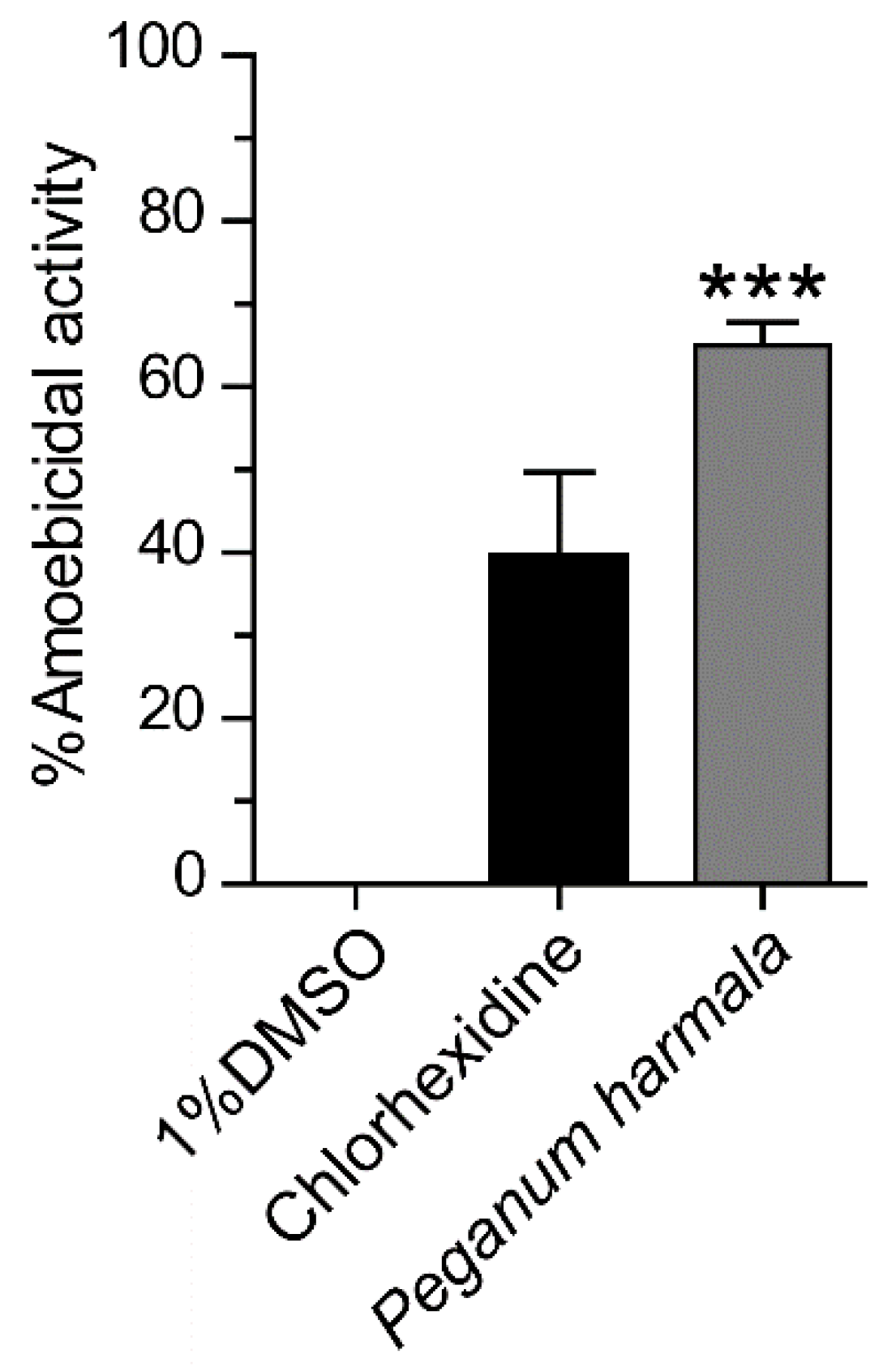
2.1. Anti-Acanthamoeba Activity of Peganum harmala Seed Extract
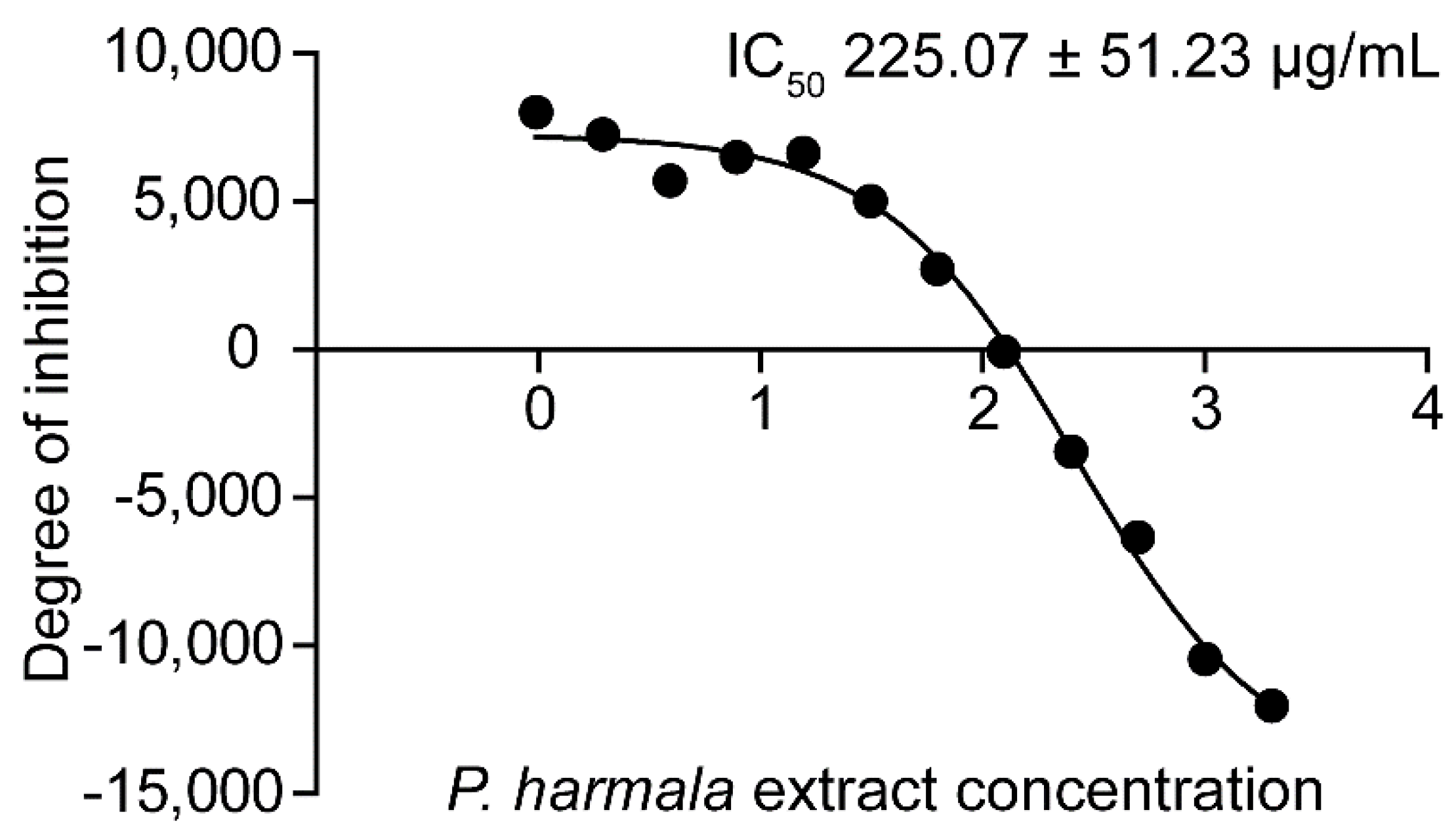
2.2. Determination of IC50 of Peganum harmala Extract and Its Morphological Effects
2.3. Drug Combination
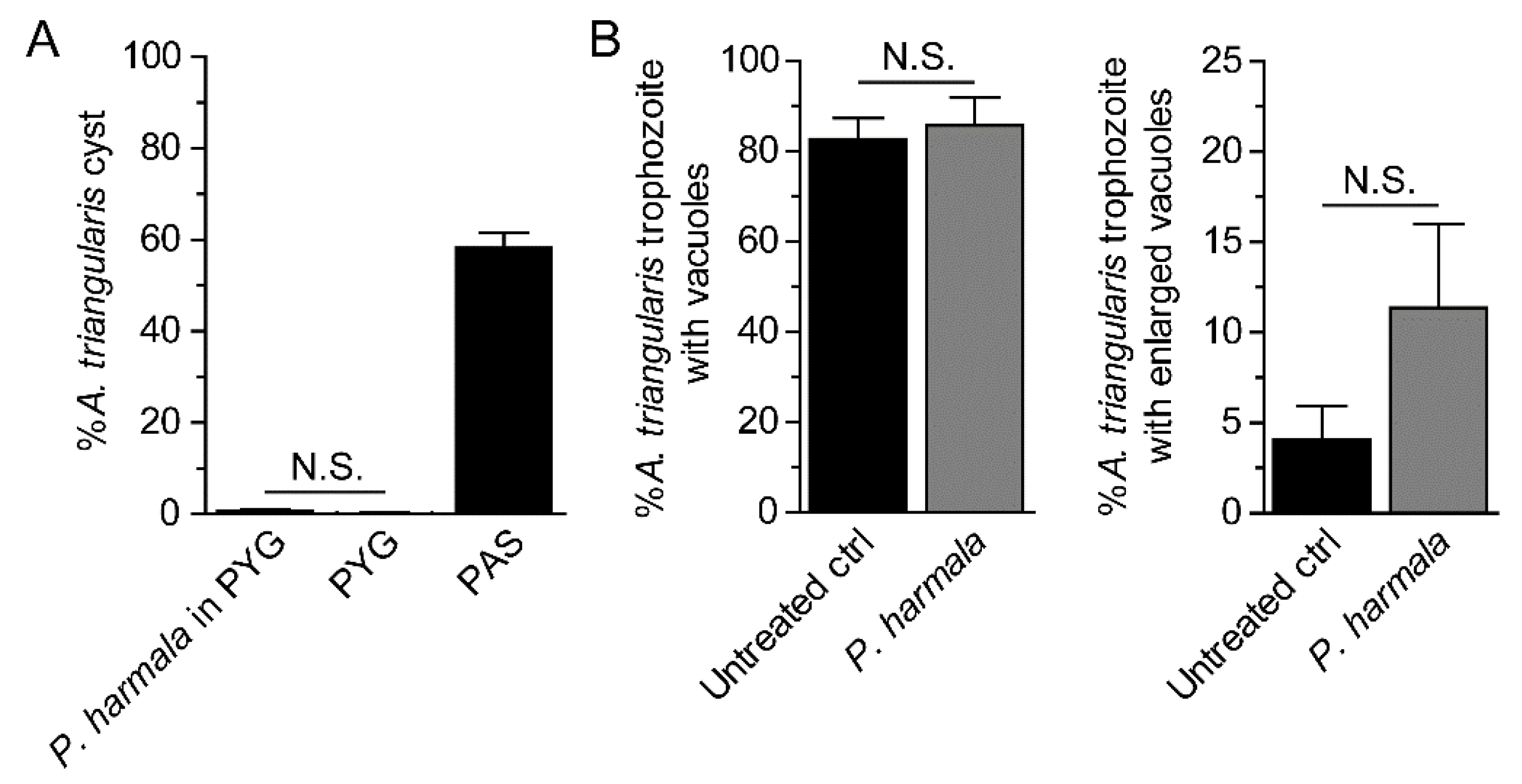
2.4. Analysis of Cyst Formation and Vacuolization
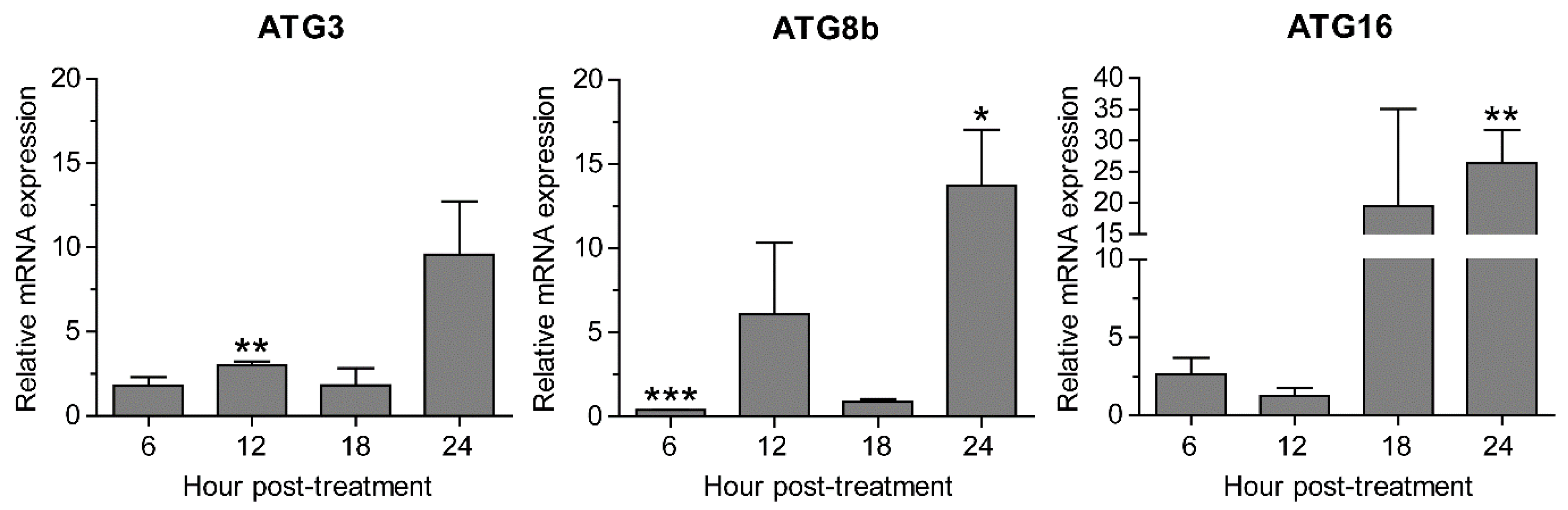
2.5. Transcriptional Expression of Autophagy-Related Genes after Peganum harmala Extract Treatment
3. Discussion
4. Materials and Methods
4.1. Plant Collection and Preparation of Extract
4.2. Acanthamoeba Cultivation
4.3. Determination of Anti-Acanthamoeba Activity
4.4. Determination of Inhibitory Concentration 50 (IC50)
4.5. Analysis of Cyst Formation and Vacuolization
4.6. Scanning Electron Microscopy
4.7. Determination of Minimal Inhibitory Concentration (MIC) and Drug Combination
4.8. Preparation of Total RNA and cDNA Synthesis
4.9. Validation of PCR Primers
4.10. Analysis of Gene Expression by Quantitative PCR
4.11. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, N.A. Acanthamoeba: Biology and increasing importance in human health. FEMS Microbiol. Rev. 2006, 30, 564–595. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo-Morales, J.; Khan, N.A.; Walochnik, J. An update on Acanthamoeba keratitis: Diagnosis, pathogenesis and treatment. Parasite 2015, 22, 10. [Google Scholar] [CrossRef] [Green Version]
- de Lacerda, A.G.; Lira, M. Acanthamoeba keratitis: A review of biology, pathophysiology and epidemiology. Ophthalmic Physiol. Opt. 2021, 41, 116–135. [Google Scholar] [CrossRef]
- Neelam, S.; Niederkorn, J.Y. Focus: Infectious diseases: Pathobiology and immunobiology of Acanthamoeba keratitis: Insights from animal models. Yale J. Biol. Med. 2017, 90, 261. [Google Scholar]
- Rayamajhee, B.; Subedi, D.; Peguda, H.K.; Willcox, M.D.; Henriquez, F.L.; Carnt, N. A systematic review of intracellular microorganisms within Acanthamoeba to understand potential impact for infection. Pathogens 2021, 10, 225. [Google Scholar] [CrossRef]
- Morrison, A.O.; Morris, R.; Shannon, A.; Lauer, S.R.; Guarner, J.; Kraft, C.S. Disseminated Acanthamoeba infection presenting with cutaneous lesions in an immunocompromised patient: A case report, review of histomorphologic findings, and potential diagnostic pitfalls. Am. J. Clin. Pathol. 2016, 145, 266–270. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Syms, M.J.; Holtel, M.R.; Nauschuetz, K.K. Acanthamoeba sinusitis with subsequent dissemination in an AIDS patient. Ear Nose Throat J. 2000, 79, 168–174. [Google Scholar] [CrossRef]
- Matson, D.O.; Rouah, E.; Lee, R.T.; Armstrong, D.; Parke, J.T.; Baker, C.J. Acanthameba meningoencephalitis masquerading as neurocysticercosis. Pediatr. Infect. Dis. J. 1988, 7, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A.; Anwar, A.; Siddiqui, R. Acanthamoeba keratitis: Current status and urgent research priorities. Curr. Med. Chem. 2019, 26, 5711–5726. [Google Scholar] [CrossRef]
- Rayamajhee, B.; Willcox, M.D.; Henriquez, F.L.; Petsoglou, C.; Carnt, N. Acanthamoeba keratitis: An increasingly common infectious disease of the cornea. Lancet Microbe 2021. [Google Scholar] [CrossRef]
- Juarez, M.M.; Tártara, L.I.; Cid, A.G.; Real, J.P.; Bermúdez, J.M.; Rajal, V.B.; Palma, S.D. Acanthamoeba in the eye, can the parasite hide even more? Latest developments on the disease. Cont. Lens Anterior Eye 2018, 41, 245–251. [Google Scholar] [CrossRef]
- Hasni, I.; Andréani, J.; Colson, P.; La Scola, B. Description of virulent factors and horizontal gene transfers of keratitis-associated amoeba Acanthamoeba triangularis by genome analysis. Pathogens 2020, 9, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuan, Y.-H.; Chung, B.-S.; Hong, Y.-C.; Kong, H.-H.; Hahn, T.-W.; Chung, D.-I. Keratitis by Acanthamoeba triangularis: Report of cases and characterization of isolates. Korean J. Parasitol. 2008, 46, 157. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.; Khan, N.A.; Siddiqui, R. Combating Acanthamoeba spp. cysts: What are the options? Parasite Vector 2018, 11, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abjani, F.; Khan, N.A.; Yousuf, F.A.; Siddiqui, R. Targeting cyst wall is an effective strategy in improving the efficacy of marketed contact lens disinfecting solutions against Acanthamoeba castellanii cysts. Contact Lens Anterior Eye 2016, 39, 239–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur, T.; Hadaś, E.; Iwanicka, I. The duration of the cyst stage and the viability and virulence of Acanthamoeba isolates. Trop. Med. Parasitol. (GTZ) 1995, 46, 106–108. [Google Scholar]
- Huang, F.-C.; Shih, M.-H.; Chang, K.-F.; Huang, J.-M.; Shin, J.-W.; Lin, W.-C. Characterizing clinical isolates of Acanthamoeba castellanii with high resistance to polyhexamethylene biguanide in Taiwan. J. Microbiol. Immunol. Infect. 2017, 50, 570–577. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res. 2014, 24, 24–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yorimitsu, T.; Klionsky, D.J. Autophagy: Molecular machinery for self-eating. Cell Death Differ. 2005, 12, 1542–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Park, S.; Biering, S.B.; Selleck, E.; Liu, C.Y.; Zhang, X.; Fujita, N.; Saitoh, T.; Akira, S.; Yoshimori, T. The parasitophorous vacuole membrane of Toxoplasma gondii is targeted for disruption by ubiquitin-like conjugation systems of autophagy. Immunity 2014, 40, 924–935. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-H.; Shenoy, A.R.; Kumar, P.; Das, R.; Tiwari, S.; MacMicking, J.D. A family of IFN-γ–inducible 65-kD GTPases protects against bacterial infection. Science 2011, 332, 717–721. [Google Scholar] [CrossRef]
- Menzies, F.M.; Fleming, A.; Rubinsztein, D.C. Compromised autophagy and neurodegenerative diseases. Nat. Rev. Neurosci. 2015, 16, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Guhde, G.; Suter, A.; Eskelinen, E.-L.; Hartmann, D.; Lüllmann-Rauch, R.; Janssen, P.M.; Blanz, J.; Von Figura, K.; Saftig, P. Accumulation of autophagic vacuoles and cardiomyopathy in LAMP-2-deficient mice. Nature 2000, 406, 902–906. [Google Scholar] [CrossRef] [PubMed]
- Khambu, B.; Yan, S.; Huda, N.; Liu, G.; Yin, X.-M. Autophagy in non-alcoholic fatty liver disease and alcoholic liver disease. Liver Res. 2018, 2, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Moon, E.-K.; Hong, Y.; Chung, D.-I.; Kong, H.-H. Autophagy protein 12 plays an essential role in Acanthamoeba encystation. Exp. Parasitol. 2015, 159, 46–52. [Google Scholar] [CrossRef]
- Moon, E.-K.; Chung, D.-I.; Hong, Y.-C.; Kong, H.-H. Autophagy protein 8 mediating autophagosome in encysting Acanthamoeba. Mol. Biochem. Parasitol. 2009, 168, 43–48. [Google Scholar] [CrossRef]
- Picazarri, K.; Nakada-Tsukui, K.; Nozaki, T. Autophagy during proliferation and encystation in the protozoan parasite Entamoeba invadens. Infect. Immun. 2008, 76, 278–288. [Google Scholar] [CrossRef] [Green Version]
- Song, S.-M.; Han, B.-I.; Moon, E.-K.; Lee, Y.-R.; Yu, H.S.; Jha, B.K.; Danne, D.-B.S.; Kong, H.-H.; Chung, D.-I.; Hong, Y. Autophagy protein 16-mediated autophagy is required for the encystation of Acanthamoeba castellanii. Mol. Biochem. Parasitol. 2012, 183, 158–165. [Google Scholar] [CrossRef]
- Moon, E.-K.; Chung, D.-I.; Hong, Y.; Kong, H.-H. Atg3-mediated lipidation of Atg8 is involved in encystation of Acanthamoeba. Korean J. Parasitol. 2011, 49, 103. [Google Scholar] [CrossRef]
- Moon, E.-K.; Hong, Y.; Chung, D.-I.; Kong, H.-H. Identification of atg8 isoform in encysting Acanthamoeba. Korean J. Parasitol. 2013, 51, 497. [Google Scholar] [CrossRef]
- Ibrahim, Y.W.; Boase, D.L.; Cree, I.A. How could contact lens wearers be at risk of Acanthamoeba infection? A review. J. Optom. 2009, 2, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, S.E.; Ivarsen, A.; Hjortdal, J. Increasing incidence of Acanthamoeba keratitis in a large tertiary ophthalmology department from year 1994 to 2018. Acta Ophthalmol. 2020, 98, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Iovieno, A.; Oechsler, R.A.; Ledee, D.R.; Miller, D.; Alfonso, E.C. Drug-resistant severe Acanthamoeba keratitis caused by rare T5 Acanthamoeba genotype. Eye Contact Lens 2010, 36, 183–184. [Google Scholar] [CrossRef]
- Szentmáry, N.; Daas, L.; Shi, L.; Laurik, K.L.; Lepper, S.; Milioti, G.; Seitz, B. Acanthamoeba keratitis–Clinical signs, differential diagnosis and treatment. J. Curr. Ophthalmol. 2019, 31, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Niyyati, M.; Dodangeh, S.; Lorenzo-Morales, J. A review of the current research trends in the application of medicinal plants as a source for novel therapeutic agents against Acanthamoeba infections. Iran. J. Pharm. Res. 2016, 15, 893. [Google Scholar]
- Sanguan, S.; Wannasan, A.; Junkum, A.; Jitpakdi, A.; Riyong, D.; Champakaew, D.; Pitasawat, B. Screening for in vitro amoebicidal activity of plant essential oils against Acanthamoeba sp. Chiang Mai Med. J. 2018, 57, 89–98. [Google Scholar]
- Mehta, K.C.; Dargad, R.R.; Borade, D.M.; Swami, O.C. Burden of antibiotic resistance in common infectious diseases: Role of antibiotic combination therapy. J. Clin. Diagn. Res. JCDR 2014, 8, ME05. [Google Scholar] [CrossRef] [PubMed]
- Tyers, M.; Wright, G.D. Drug combinations: A strategy to extend the life of antibiotics in the 21st century. Nat. Rev. Microbiol. 2019, 17, 141–155. [Google Scholar] [CrossRef]
- Mitsuwan, W.; Bunsuwansakul, C.; Leonard, T.E.; Laohaprapanon, S.; Hounkong, K.; Bunluepuech, K.; Chalermpol, K.; Mahboob, T.; Sumudi Raju, C.; Dhobi, M. Curcuma longa ethanol extract and Curcumin inhibit the growth of Acanthamoeba triangularis trophozoites and cysts isolated from water reservoirs at Walailak University, Thailand. Pathog. Glob. Health 2020, 114, 194–204. [Google Scholar] [CrossRef]
- Kolören, O.; Kolören, Z.; Şekeroğlu, Z.A.; Colayvaz, M.; Karanis, P. Amoebicidal and amoebistatic effects of Artemisia argyi methanolic extracts on Acanthamoeba castellanii trophozoites and cysts. Acta Parasitol. 2019, 64, 63–70. [Google Scholar] [CrossRef]
- Anwar, A.; Ting, E.L.S.; Anwar, A.; ul Ain, N.; Faizi, S.; Shah, M.R.; Khan, N.A.; Siddiqui, R. Antiamoebic activity of plant-based natural products and their conjugated silver nanoparticles against Acanthamoeba castellanii (ATCC 50492). AMB Express 2020, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Moloudizargari, M.; Mikaili, P.; Aghajanshakeri, S.; Asghari, M.H.; Shayegh, J. Pharmacological and therapeutic effects of Peganum harmala and its main alkaloids. Pharmacogn. Rev. 2013, 7, 199. [Google Scholar] [CrossRef] [Green Version]
- Seyed Hassan Tehrani, S.; Hashemi Sheikh Shabani, S.; Tahmasebi Enferadi, S.; Rabiei, Z. Growth inhibitory impact of Peganum harmala L. on two breast cancer cell lines. Iran. J. Biotechnol. 2014, 12, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Khlifi, D.; Sghaier, R.M.; Amouri, S.; Laouini, D.; Hamdi, M.; Bouajila, J. Composition and anti-oxidant, anti-cancer and anti-inflammatory activities of Artemisia herba-alba, Ruta chalpensis L. and Peganum harmala L. Food Chem. Toxicol. 2013, 55, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Davoodi, H.; Ghaemi, E.; Mazandarani, M.; Shakeri, F.; Javid, S.N.; Klishadi, M. Anti-mycobacterial and anti-inflammatory activity of Peganum harmala. J. Chem. Pharm. Res. 2015, 7, 1611–1616. [Google Scholar]
- Moradi, M.-T.; Karimi, A.; Rafieian-Kopaei, M.; Fotouhi, F. In vitro antiviral effects of Peganum harmala seed extract and its total alkaloids against influenza virus. Microb. Pathog. 2017, 110, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Nenaah, G. Antibacterial and antifungal activities of (beta)-carboline alkaloids of Peganum harmala (L.) seeds and their combination effects. Fitoterapia 2010, 81, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Rahimi-Moghaddam, P.; Ebrahimi, S.A.; Ourmazdi, H.; Selseleh, M.; Karjalian, M.; Haj-Hassani, G.; Alimohammadian, M.H.; Mahmoudian, M.; Shafiei, M. In vitro and in vivo activities of Peganum harmala extract against Leishmania major. J. Res. Med. Sci. 2011, 16, 1032. [Google Scholar] [PubMed]
- Tanweer, A.J.; Saddique, U.; Bailey, C.; Khan, R. Antiparasitic effect of wild rue (Peganum harmala L.) against experimentally induced coccidiosis in broiler chicks. Parasitol. Res. 2014, 113, 2951–2960. [Google Scholar] [CrossRef]
- Shohaib, H.M.; Nawaz, S.; Matin, A. Methanolic extract of Peganum harmala exhibit potent activity against Acanthamoeba castellanii cysts and its encystment in vitro. Pak. J. Pharm. Sci 2016, 29, 1993–1996. [Google Scholar]
- Heredero-Bermejo, I.; Martín-Pérez, T.; Copa-Patiño, J.L.; Gómez, R.; de la Mata, F.J.; Soliveri, J.; Pérez-Serrano, J. Ultrastructural study of Acanthamoeba polyphaga trophozoites and cysts treated in vitro with cationic carbosilane dendrimers. Pharmaceutics 2020, 12, 565. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X.; Gueydan, C.; Han, J. Plasma membrane changes during programmed cell deaths. Cell Res. 2018, 28, 9–21. [Google Scholar] [CrossRef]
- Lazuana, T.; Astuty, H.; Sari, I.P. Effect of cellulase enzyme Treatment on cyst wall degradation of Acanthamoeba sp. J. Parasitol. Res. 2019, 2019, 8915314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munafó, D.B.; Colombo, M.I. A novel assay to study autophagy: Regulation of autophagosome vacuole size by amino acid deprivation. J. Cell Sci. 2001, 114, 3619–3629. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Mashima, H.; Miura, K.; Maeda, D.; Goto, A.; Goto, T.; Sun-Wada, G.-H.; Wada, Y.; Ohnishi, H. Disruption of small GTPase Rab7 exacerbates the severity of acute pancreatitis in experimental mouse models. Sci. Rep. 2017, 7, 2817. [Google Scholar] [CrossRef]
- Boonhok, R.; Sangkanu, S.; Norouzi, R.; Siyadatpanah, A.; Mirzaei, F.; Mitsuwan, W.; Charong, N.; Wisessombat, S.; de Lourdes Pereira, M.; Rahmatullah, M. Amoebicidal activity of Cassia angustifolia extract and its effect on Acanthamoeba triangularis autophagy-related gene expression at the transcriptional level. Parasitology 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bouyer, S.; Rodier, M.-H.; Guillot, A.; Héchard, Y. Acanthamoeba castellanii: Proteins involved in actin dynamics, glycolysis, and proteolysis are regulated during encystation. Exp. Parasitol. 2009, 123, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Moon, E.-K.; Hong, Y.; Chung, D.-I.; Kong, H.-H. Cysteine protease involving in autophagosomal degradation of mitochondria during encystation of Acanthamoeba. Mol. Biochem. Parasitol. 2012, 185, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Gatica, D.; Lahiri, V.; Klionsky, D.J. Cargo recognition and degradation by selective autophagy. Nat. Cell Biol. 2018, 20, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Garajová, M.; Mrva, M.; Vaškovicová, N.; Martinka, M.; Melicherová, J.; Valigurová, A. Cellulose fibrils formation and organisation of cytoskeleton during encystment are essential for Acanthamoeba cyst wall architecture. Sci. Rep. 2019, 9, 4466. [Google Scholar] [CrossRef]
- Lakhundi, S.; Siddiqui, R.; Khan, N.A. Cellulose degradation: A therapeutic strategy in the improved treatment of Acanthamoeba infections. Parasit Vectors 2015, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, E.-K.; Kong, H.-H. Short-cut pathway to synthesize cellulose of encysting Acanthamoeba. Korean J. Parasitol. 2012, 50, 361. [Google Scholar] [CrossRef]
- Moon, E.-K.; Hong, Y.; Chung, D.-I.; Goo, Y.-K.; Kong, H.-H. Down-regulation of cellulose synthase inhibits the formation of endocysts in Acanthamoeba. Korean J. Parasitol. 2014, 52, 131. [Google Scholar] [CrossRef]
- Moon, E.-K.; Chung, D.-I.; Hong, Y.-C.; Kong, H.-H. Differentially expressed genes of Acanthamoeba castellanii during encystation. Korean J. Parasitol. 2007, 45, 283. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.-T.; Kong, H.-H.; Ha, Y.-R.; Hong, Y.-C.; Jeong, H.J.; Yu, H.S.; Chung, D.-I. Comparison of specific activity and cytopathic effects of purified 33 kDa serine proteinase from Acanthamoeba strains with different degree of virulence. Korean J. Parasitol. 2006, 44, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano-Luna, J.d.J.; Cervantes-Sandoval, I.; Calderón, J.; Navarro-García, F.; Tsutsumi, V.; Shibayama, M. Protease activities of Acanthamoeba polyphaga and Acanthamoeba castellanii. Can. J. Microbiol. 2006, 52, 16–23. [Google Scholar] [CrossRef]
- Moon, E.-K.; Chung, D.-I.; Hong, Y.-C.; Kong, H.-H. Characterization of a serine proteinase mediating encystation of Acanthamoeba. Eukaryot. Cell 2008, 7, 1513–1517. [Google Scholar] [CrossRef] [Green Version]
- Vongsak, B.; Sithisarn, P.; Mangmool, S.; Thongpraditchote, S.; Wongkrajang, Y.; Gritsanapan, W. Maximizing total phenolics, total flavonoids contents and antioxidant activity of Moringa oleifera leaf extract by the appropriate extraction method. Ind. Crop. Prod. 2013, 44, 566–571. [Google Scholar] [CrossRef]
- Taravaud, A.; Loiseau, P.M.; Pomel, S. In vitro evaluation of antimicrobial agents on Acanthamoeba sp. and evidence of a natural resilience to amphotericin B. Int. J. Parasitol Drugs Drug Resist. 2017, 7, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Aqeel, Y.; Siddiqui, R.; Iftikhar, H.; Khan, N.A. The effect of different environmental conditions on the encystation of Acanthamoeba castellanii belonging to the T4 genotype. Exp. Parasitol. 2013, 135, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, F.A.; Boonhok, R.; Cabrera, M.; Mbenda, H.G.N.; Wang, M.; Min, H.; Liang, X.; Qin, J.; Zhu, X.; Miao, J. Role of Plasmodium falciparum Kelch 13 protein mutations in P. falciparum populations from northeastern Myanmar in mediating artemisinin resistance. mBio 2020, 11, e01134-19. [Google Scholar] [CrossRef] [Green Version]
- Sangkanu, S.; Mitsuwan, W.; Mahabusarakam, W.; Jimoh, T.O.; Wilairatana, P.; Girol, A.P.; Verma, A.K.; de Lourdes Pereira, M.; Rahmatullah, M.; Wiart, C. Anti-Acanthamoeba synergistic effect of chlorhexidine and Garcinia mangostana extract or α-mangostin against Acanthamoeba triangularis trophozoite and cyst forms. Sci. Rep. 2021, 11, 8053. [Google Scholar] [CrossRef] [PubMed]
- Chanda, S.; Rakholiya, K. Combination therapy: Synergism between natural plant extracts and antibiotics against infectious diseases. Microbiol. Book Ser. 2011, 1, 520–529. [Google Scholar]
- Lo, E.; Nguyen, J.; Oo, W.; Hemming-Schroeder, E.; Zhou, G.; Yang, Z.; Cui, L.; Yan, G. Examining Plasmodium falciparum and P. vivax clearance subsequent to antimalarial drug treatment in the Myanmar-China border area based on quantitative real-time polymerase chain reaction. BMC Infect. Dis. 2016, 16, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boonhok, R.; Sangkanu, S.; Chuprom, J.; Srisuphanunt, M.; Norouzi, R.; Siyadatpanah, A.; Mirzaei, F.; Mitsuwan, W.; Wisessombat, S.; de Lourdes Pereira, M.; et al. Peganum harmala Extract Has Antiamoebic Activity to Acanthamoeba triangularis Trophozoites and Changes Expression of Autophagy-Related Genes. Pathogens 2021, 10, 842. https://doi.org/10.3390/pathogens10070842
Boonhok R, Sangkanu S, Chuprom J, Srisuphanunt M, Norouzi R, Siyadatpanah A, Mirzaei F, Mitsuwan W, Wisessombat S, de Lourdes Pereira M, et al. Peganum harmala Extract Has Antiamoebic Activity to Acanthamoeba triangularis Trophozoites and Changes Expression of Autophagy-Related Genes. Pathogens. 2021; 10(7):842. https://doi.org/10.3390/pathogens10070842
Chicago/Turabian StyleBoonhok, Rachasak, Suthinee Sangkanu, Julalak Chuprom, Mayuna Srisuphanunt, Roghayeh Norouzi, Abolghasem Siyadatpanah, Farzaneh Mirzaei, Watcharapong Mitsuwan, Sueptrakool Wisessombat, Maria de Lourdes Pereira, and et al. 2021. "Peganum harmala Extract Has Antiamoebic Activity to Acanthamoeba triangularis Trophozoites and Changes Expression of Autophagy-Related Genes" Pathogens 10, no. 7: 842. https://doi.org/10.3390/pathogens10070842
APA StyleBoonhok, R., Sangkanu, S., Chuprom, J., Srisuphanunt, M., Norouzi, R., Siyadatpanah, A., Mirzaei, F., Mitsuwan, W., Wisessombat, S., de Lourdes Pereira, M., Rahmatullah, M., Wilairatana, P., Wiart, C., Ling, L. C., Dolma, K. G., & Nissapatorn, V. (2021). Peganum harmala Extract Has Antiamoebic Activity to Acanthamoeba triangularis Trophozoites and Changes Expression of Autophagy-Related Genes. Pathogens, 10(7), 842. https://doi.org/10.3390/pathogens10070842