
Rabies in Our Neighbourhood: Preparedness for an Emerging Infectious Disease


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. The Nature of Rabies: A Summary
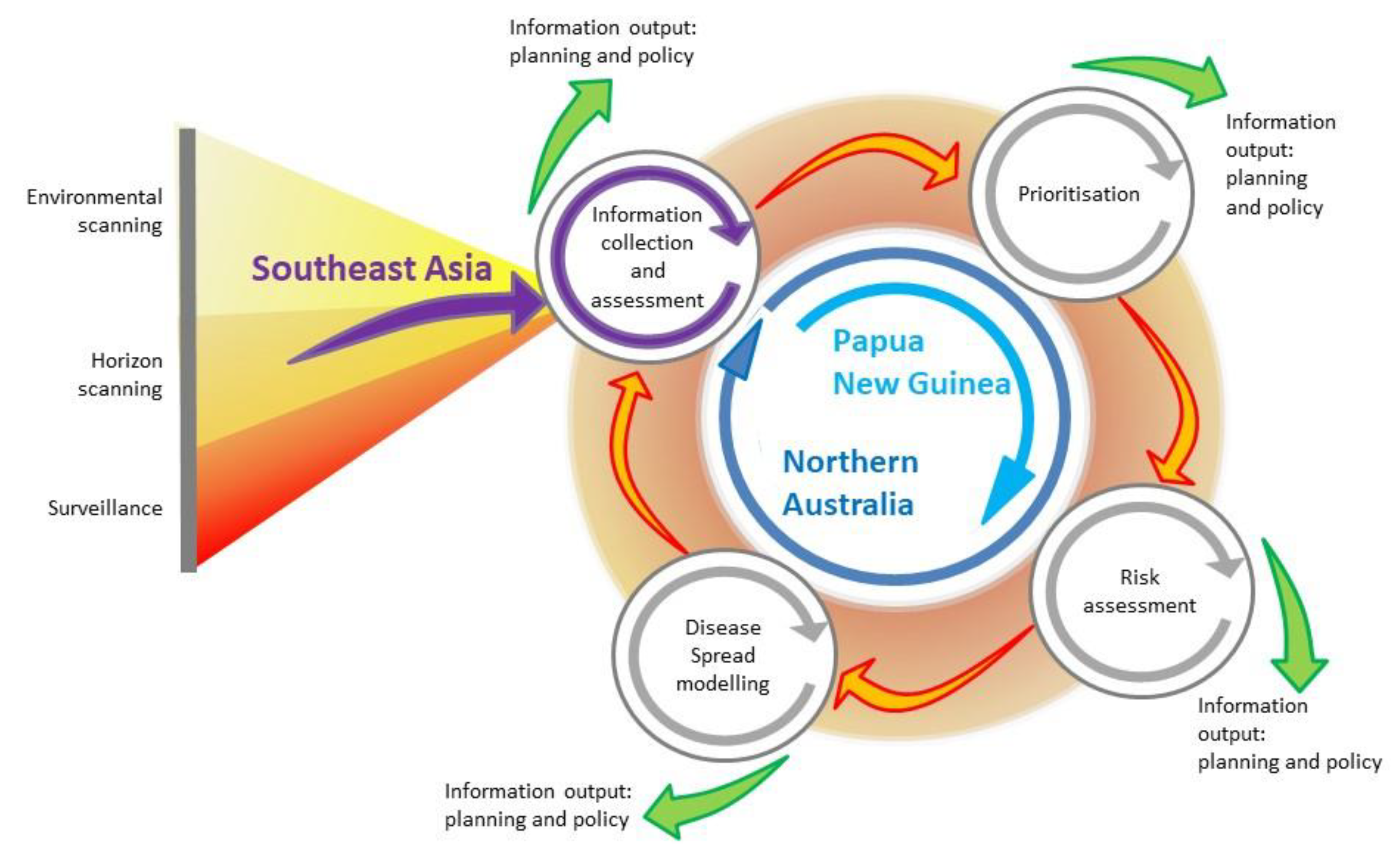
3. The Long-Term Risk of RABV to Australia: Foresighting, Environmental and Horizon Scanning, and Surveillance
4. Prioritisation and Risk Assessment: The Current Risk of RABV to Australia
5. Disease Modelling: Mitigating Expected Impacts of RABV in Australia
6. Discussion
- An increase in demand for hunting dogs. In this area, dogs are used to hunt pigs, and it appears that most of the meat is consumed within villages. However, demographic changes in the region (for example, increased disposable income, or changes in the cultural mix of the population) in which there is an increased demand for pig meat and therefore an increased price paid, could be a driver of a larger hunting dog population and therefore increased rabies spread.
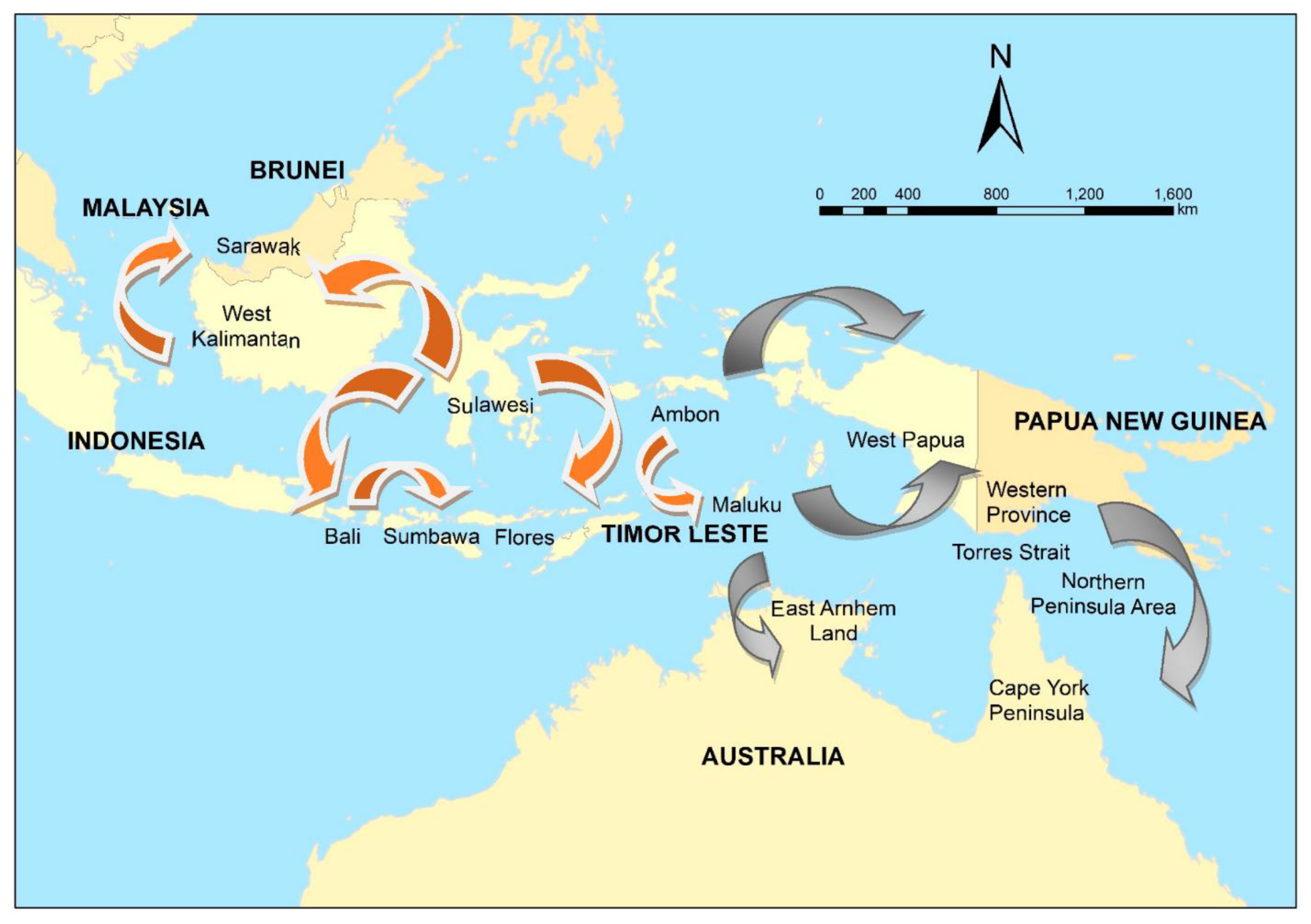
- An increase in demand for dogs to guard crops. Dogs are used within the region to guard crops from wildlife, such as wild pigs and monkeys. As above, an increase in demand for crops due to immigration or socioeconomic changes could drive an increase demand for guard dogs. In 2019, a rabies incursion in Sumbawa, Indonesia was reported. Initial investigations suggested that this incursion in the previously rabies-free island was due to the movement of dogs from either Bali or Sulawesi, Indonesia, for the purposes of guarding corn crops from wild monkeys.
- Ethnic, cultural and religious trends. In the district studied, it appeared that in rural areas rabies is more likely in villages that are predominantly Christian, versus those that are predominantly Muslim. It is also likely associated with indigenous Dayak populations. The association between ethnic, cultural and religious affiliations and dog ownership and movement have been previously described [93].
- Gifting of puppies has been linked to the spread of rabies to previously free areas, such as Tanibar, Maluku Province, Indonesia. It appears that gifting puppies is a traditional practice in the region. However, increased disposable income and an increase in pet shops or other pet traders might increase the risk of rabies spread.
- Dog meat consumption. Most dogs appear to eventually be consumed when their other uses are exhausted. This is a local activity and likely not a potential driver of regional rabies spread. However, the most likely reason for long-distance movement is supply of dogs for consumption (especially during the harvest festivals between January and June) if dogs for consumption are scarce in the local area. Gawai (harvest festivals) are held in every village between January and July. The time varies between villages due to the harvest time. There is also a district Gawai. Dogs are consumed during these festivals. This system of festivals appears to be relatively constant (and perhaps even decreasing in popularity). Changes that promote traditional festivals—such as an increase in tourism—could create a scenario of rabies spread via dog movements for consumption.
- Regional road improvements. Major roads throughout the region have been sealed within the past 5 years. Better roads enable increased trade and travel. If dogs are taken on such trips, then the risk of rabies spread can be expected to increase in a future scenario.
- Oil palm plantations. The increasing number of oil palm plantations in the region has provided opportunities for work and trade. The oil palm plantations are numerous on the Malaysian side of the border and have increased unofficial, local border travel. Indonesians work on Malaysian owned plantations (forest on the Indonesian side, oil palm on the Malaysian side) and also trade (in Malaysian currency). Dogs are used on oil palm plantations for protection, although details on the roles played by dogs on oil palm plantations are not readily available.
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gardner, J.W.; Stewart, W.H. Use of Vital and Health Records in Epidemiologic Research. A Report of the United States National Committee on Vital and Health Statistics; U.S. Department of Health, Education, and Welfare: Washington, DC, USA, 1968. Available online: http://www.cdc.gov/nchs/data/series/sr_04/sr04_007.pdf (accessed on 24 February 2021).
- Wells, G.A.; Scott, A.C.; Johnson, C.T.; Gunning, R.F.; Hancock, R.D.; Jeffrey, M.; Dawson, M.; Bradley, R. A novel progressive spongiform encephalopathy in cattle. Vet. Rec. 1987, 121, 419–420. [Google Scholar] [CrossRef] [PubMed]
- Will, R.; Ironside, J.; Zeidler, M.; Estibeiro, K.; Cousens, S.; Smith, P.; Alperovitch, A.; Poser, S.; Pocchiari, M.; Hofman, A. A new variant of Creutzfeldt-Jakob disease in the UK. Lancet 1996, 347, 921–925. [Google Scholar] [CrossRef]
- Sharp, P.M.; Bailes, E.; Chaudhuri, R.R.; Rodenburg, C.M.; Santiago, M.O.; Hahn, B.H. The origins of acquired immune deficiency syndrome viruses: Where and when? Philos. Trans. R. Soc. B Biol. Sci. 2001, 356, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.P.; Aucken, H.M.; Cavendish, S.; Ganner, M.; Wale, M.C.J.; Warner, M.; Livermore, D.M.; Cookson, B.D.; Participants, T.U.E. Dominance of EMRSA-15 and -16 among MRSA causing nosocomial bacteraemia in the UK: Analysis of isolates from the European Antimicrobial Resistance Surveillance System (EARSS). J. Antimicrob. Chemother. 2001, 48, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nat. Cell Biol. 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Raj, V.S.; Osterhaus, A.D.; Fouchier, R.A.; Haagmans, B.L. MERS: Emergence of a novel human coronavirus. Curr. Opin. Virol. 2014, 5, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Baize, S.; Pannetier, D.; Oestereich, L.; Rieger, T.; Koivogui, L.; Magassouba, N.; Soropogui, B.; Sow, M.S.; Kéïta, S.; De Clerck, H.; et al. Emergence of Zaire Ebola Virus Disease in Guinea. N. Engl. J. Med. 2014, 371, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Diamond, M.S. The emergence of Zika virus and its new clinical syndromes. Nat. Cell Biol. 2018, 560, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Cleaveland, S.; Hampson, K. Rabies elimination research: Juxtaposing optimism, pragmatism and realism. Proc. R. Soc. B Boil. Sci. 2017, 284, 20171880. [Google Scholar] [CrossRef]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important Reservoir Hosts of Emerging Viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Magouras, I.; Brookes, V.J.; Jori, F.; Martin, A.; Pfeiffer, D.U.; Dürr, S. Emerging Zoonotic Diseases: Should We Rethink the Animal–Human Interface? Front. Vet. Sci. 2020, 7, 582743. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.G.; Sawleshwarkar, S.; Hossain, S.; Mor, S.M. Whence the next pandemic? The intersecting global geography of the animal-human interface, poor health systems and air transit centrality reveals conduits for high-impact spillover. One Health 2020, 11, 100177. [Google Scholar] [CrossRef] [PubMed]
- Brookes, V.J.; Hernández-Jover, M.; Black, P.F.; Ward, M.P. Preparedness for emerging infectious diseases: Pathways from anticipation to action. Epidemiol. Infect. 2015, 143, 2043–2058. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, C.E.; Freuling, C.M.; Mani, R.S.; Palacios, C.; Sabeta, C.T.; Ward, M. A history of rabies—The foundation for global canine rabies elimination. In Rabies, Scientific Basis of the Disease and Its Management, 4th ed.; Fooks, A.R., Jackson, A.C., Eds.; Academic Press: London, UK, 2020; Chapter 1; pp. 1–44. ISBN 978-0-12-818705-0. [Google Scholar]
- Hampson, K.; Coudeville, L.; Lembo, T.; Sambo, M.; Kieffer, A.; Attlan, M.; Barrat, J.; Blanton, J.D.; Briggs, D.J.; Cleaveland, S.; et al. Estimating the Global Burden of Endemic Canine Rabies. PLOS Neglect. Trop. Dis. 2015, 9, e0003709. [Google Scholar] [CrossRef]
- World Health Organization. WHO Expert Consultation on Rabies: Second Report; World Health Organization: Geneva, Switzerland, 2013; Available online: http://www.who.int/iris/handle/10665/85346 (accessed on 24 February 2021).
- Hampson, K.; Dushoff, J.; Cleaveland, S.; Haydon, D.T.; Kaare, M.; Packer, C.; Dobson, A. Transmission Dynamics and Prospects for the Elimination of Canine Rabies. PLoS Biol. 2009, 7, e53. [Google Scholar] [CrossRef] [PubMed]
- Fekadu, M.; Chandler, F.W.; Harrison, A.K. Pathogenesis of rabies in dogs inoculated with an Ethiopian rabies virus strain. Immunofluorescence, histologic and ultrastructural studies of the central nervous system. Arch. Virol. 1982, 71, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Tojinbara, K.; Sugiura, K.; Yamada, A.; Kakitani, I.; Kwan, N. Estimating the probability distribution of the incubation period for rabies using data from the 1948–1954 rabies epidemic in Tokyo. Prev. Vet. Med. 2016, 123, 102–105. [Google Scholar] [CrossRef]
- World Organization for Animal Health (OIE). Rabies (infection with rabies virus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; Paris, France, 2017; Available online: http://www.oie.int/en/international-standard-setting/terrestrial-manual/access-online/ (accessed on 24 February 2021).
- Zinsstag, J.; Durr, S.; Penny, M.A.; Mindekem, R.; Roth, F.; Gonzalez, S.M.; Naissengar, S.; Hattendorf, J. Transmission dynamics and economics of rabies control in dogs and humans in an African city. Proc. Natl. Acad. Sci. USA 2009, 106, 14996–15001. [Google Scholar] [CrossRef] [PubMed]
- Lembo, T.; Attlan, M.; Bourhy, H.; Cleaveland, S.; Costa, P.; De Balogh, K.; Dodet, B.; Fooks, A.R.; Hiby, E.; Leanes, F.; et al. Renewed Global Partnerships and Redesigned Roadmaps for Rabies Prevention and Control. Vet. Med. Int. 2011, 2011, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Vallat, B. The OIE’s commitment to fight rabies worldwide. Rabies: A priority for humans and animals. OIE Bull. 2011, 3, 1–2. [Google Scholar]
- Dodet, B. Report of the sixth AREB meeting, Manila, The Philippines, 10–12 November 2009: Conference report. Vaccine 2010, 28, 3265–3268. [Google Scholar] [CrossRef] [PubMed]
- Harischandra, P.A.L. Dog rabies control program and achievement in Sri Lanka. In Proceedings of the Third Rabies in Asia conference, Colombo, Sri Lanka, 28−30 November 2011. [Google Scholar]
- Bögel, K.; Meslin, F.X. Economics of human and canine rabies elimination: Guidelines for programme orientation. Bull. World Health Organ. 1990, 68, 281–291. [Google Scholar] [PubMed]
- Knobel, D.L.; Cleaveland, S.; Coleman, P.G.; Fèvre, E.M.; Meltzer, M.I.; Miranda, M.E.; Shaw, A.; Zinsstag, J.; Meslin, F.X. Re-evaluating the burden of rabies in Africa and Asia. Bull. World Health Organ. 2005, 83, 360–368. [Google Scholar] [PubMed]
- Wells, C.W. The control of rabies in Malaya through compulsory mass vaccination of dogs. Bull. World Heal. Organ. 1954, 10, 731–742. [Google Scholar]
- Shahirudin, S. Rabies control in animals in Malaysia. In Rabies Control in Asia, Proceedings of the Fourth International Symposium, Hanoi, Viet Nam, 5–9 March 2001; Dodet, B., Meslin, F.X., Eds.; Fondation Mérieux: Lyon, France; p. 2001.
- Cleaveland, S.; Kaare, M.; Knobel, D.; Laurenson, M.K. Canine vaccination—Providing broader benefits for disease control. Vet. Microbiol. 2006, 117, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Omoe, H.; Omoe, K.; Okabe, N. Regulatory Systems for Prevention and Control of Rabies, Japan. Emerg. Infect. Dis. 2008, 14, 1368–1374. [Google Scholar] [CrossRef]
- Weng, H.-Y.; Wu, P.-I.; Yang, P.-C.; Tsai, Y.-L.; Chang, C.-C. A quantitative risk assessment model to evaluate effective border control measures for rabies prevention. Vet. Res. 2009, 41, 11. [Google Scholar] [CrossRef]
- Gongal, G.; Wright, A.E. Human Rabies in the WHO Southeast Asia Region: Forward Steps for Elimination. Adv. Prev. Med. 2011, 2011, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Crowther, W.E.L.H. A case of so-called hydrophobia: A matter of diagnosis. Med. J. Aust. 1946, 1, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Robertson, W.A.N. Milestones in the Pastoral Age of Australia. Aust. Assoc. Adv. Sci. 1932, 31, 295–325. [Google Scholar]
- Pullar, E.M.; McIntosh, K.S. The relation of Australia to the world rabies problem. Aust. Vet. J. 1954, 30, 326–336. [Google Scholar] [CrossRef]
- Bek, M.D.; Levy, M.H.; Rubin, G.L.; Smith, W.T.; Sullivan, E. Rabies case in New South Wales, 1990: Public health aspects. Med. J. Aust. 1992, 156, 596–600. [Google Scholar] [CrossRef]
- Grattansmith, P.J.; Oregan, W.J.; Ellis, P.S.J.; Oflaherty, S.J.; McIntyre, P.B.; Barnes, C.J. Rabies—A 2nd Australian case, with a long incubation period. Med. J. Aust. 1992, 156, 651–654. [Google Scholar] [CrossRef]
- Australian Government, Department of Agriculture, Water and the Environment. Available online: https://www.agriculture.gov.au/biosecurity/australia/naqs/significant-events (accessed on 24 February 2021).
- Newsome, A.; Catling, P. Host range and its implications for wildlife rabies in Australia. In Wildlife Rabies Contingency Planning in Australia; O’Brien, P., Berry, G.E., Eds.; Australian Government Publishing Service: Canberra, Australia, 1992. [Google Scholar]
- Forman, A. The threat of rabies introduction and establishment in Australia. Aust. Vet. J. 1993, 70, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.G. World rabies picture—Implications for Australia. In Wildlife Rabies Contingency Planning in Australia; O’Brien, P., Berry, G.E., Eds.; Australian Government Publishing Service: Canberra, Australia, 1992. [Google Scholar]
- Animal Health Australia, 2019: AUSVETPLAN Manuals and Documents. Available online: https://animalhealthaustralia.com.au/our-publications/ausvetplan-manuals-and-documents/ (accessed on 24 February 2021).
- O’Brien, P.; Berry, G.E. Wildlife Rabies Contingency Planning in Australia: Bureau of Rural Resources Proceedings No. 11; Australian Government Publishing Service: Canberra, Australia, 1992.
- Slaughter, R.A. Futures for the Third Millenium: Enabling the Forward View; Prospect Media: Sydney, Australia, 1999. [Google Scholar]
- Frishammar, J. Characteristics in information processing approaches. Int. J. Inf. Manag. 2002, 22, 143–156. [Google Scholar] [CrossRef]
- Horton, A. A simple guide to successful foresight. Foresight 1999, 1, 5–9. [Google Scholar] [CrossRef]
- King, D.A.; Thomas, S.M. Taking science out of the box—Foresight recast. Science 2007, 316, 1701–1702. [Google Scholar] [CrossRef]
- Voros, J. A generic foresight process framework. Foresight 2003, 5, 10–21. [Google Scholar] [CrossRef]
- Willis, N.G.; Monroe, F.A.; Empringham, R.E.; Renwick, S.A.; Van der Linden, I.W.M.; Dunlop, J.R. Using foresight to prepare animal health today for tomorrow’s challenges. Can. Vet. J. 2011, 52, 614–618. [Google Scholar]
- Windiyaningsih, C.; Wilde, H.; Meslin, F.X.; Suroso, T.; Widarso, H.S. The rabies epidemic on Flores Island, Indonesia (1998–2003). J. Med. Assoc. Thail. 2004, 87, 1389–1393. [Google Scholar]
- Clifton, M. How not to fight a rabies epidemic: A history in Bali. Asian Biomed. 2010, 4, 663–670. [Google Scholar] [CrossRef]
- Putra, A.A.G.; Hampson, K.; Girardi, J.; Hiby, E.; Knobel, D.; Mardiana, W.; Townsend, S.; Scott-Orr, H. Response to a Rabies Epidemic, Bali, Indonesia, 2008–2011. Emerg. Infect. Dis. 2013, 19, 648–651. [Google Scholar] [CrossRef]
- Putra, A.A.; Mardiana, I.W.; Scott-Orr, H.; Hiby, E.; Hampson, K.; Haydon, D.T.; Girardi, J.; Knobel, D.; Townsend, S. Progress of rabies elimination program in Bali, Indonesia. In Proceedings of the Third Rabies in Asia conference, Colombo, Sri Lanka, 28−30 November 2011. [Google Scholar]
- Wera, E.; Mourits, M.C.M.; Hogeveen, H. Uptake of Rabies Control Measures by Dog Owners in Flores Island, Indonesia. PLOS Negl. Trop. Dis. 2015, 9, e0003589. [Google Scholar] [CrossRef] [PubMed]
- Townsend, S.E.; Sumantra, I.P.; Bagus, G.N.; Brum, E.; Cleaveland, S.; Crafter, S.; Dewi, A.P.; Dharma, D.M.; Dushoff, J.; Girardi, J.; et al. Designing Programs for Eliminating Canine Rabies from Islands: Bali, Indonesia as a Case Study. PLOS Negl. Trop. Dis. 2013, 7, e2372. [Google Scholar] [CrossRef] [PubMed]
- Putra, A.A.G. Rabies eradication program, Bali. One Medicine Forum. In Proceedings of the Australian Veterinary Association Annual Conference, Darwin, Australia, 18 May 2009. [Google Scholar]
- Cookson, B.; Sergeant, E.S.G.; Martin, P.A.J. Risk-based prioritisation of surveillance for exotic animal diseases in northern Australia. In Proceedings of the 13th International Symposium on Veterinary Epidemiology and Economics; International Society for Veterinary Epidemiology and Economics: Maastricht, The Netherlands, 2012; p. 379. [Google Scholar]
- Sparkes, J.; Fleming, P.J.S.; Ballard, G.; Scott-Orr, H.; Dürr, S.E.; Ward, M.P. Canine Rabies in Australia: A Review of Preparedness and Research Needs. Zoonoses Public Health 2014, 62, 237–253. [Google Scholar] [CrossRef] [PubMed]
- Brookes, V.; Hernández-Jover, M.; Cowled, B.; Holyoake, P.; Ward, M. Building a picture: Prioritisation of exotic diseases for the pig industry in Australia using multi-criteria decision analysis. Prev. Vet. Med. 2014, 113, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Howard, K.; Wilson, A.; Ward, M.; Gilbert, G.L.; Degeling, C. Public preferences for One Health approaches to emerging infectious diseases: A discrete choice experiment. Soc. Sci. Med. 2019, 228, 164–171. [Google Scholar] [CrossRef]
- Brookes, V.J.; Vilas, V.J.D.R.; Ward, M.P. Disease prioritization: What is the state of the art? Epidemiol. Infect. 2015, 143, 2911–2922. [Google Scholar] [CrossRef] [PubMed]
- Klamer, S.; Working Group Disease and Criteria Selection; Van Goethem, N.; Thomas, D.; Duysburgh, E.; Braeye, T.; Quoilin, S. Working group Epidemiologists Prioritisation for future surveillance, prevention and control of 98 communicable diseases in Belgium: A 2018 multi-criteria decision analysis study. BMC Public Health 2021, 21, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Vilas, V.J.D.R.; Burgeño, A.; Montibeller, G.; Clavijo, A.; Vigilato, M.A.; Cosivi, O. Prioritization of capacities for the elimination of dog-mediated human rabies in the Americas: Building the framework. Pathog. Glob. Health 2013, 107, 340–345. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hernández-Jover, M.; Roche, S.; Ward, M.P. The human and animal health impacts of introduction and spread of an exotic strain of West Nile virus in Australia. Prev. Vet. Med. 2013, 109, 186–204. [Google Scholar] [CrossRef]
- Ward, M.P.; Hernández-Jover, M. A generic rabies risk assessment tool to support surveillance. Prev. Vet. Med. 2015, 120, 4–11. [Google Scholar] [CrossRef]
- Goddard, A.D.; Donaldson, N.M.; Horton, D.; Kosmider, R.; Kelly, L.A.; Sayers, A.R.; Breed, A.; Freuling, C.M.; Muller, T.; Shaw, S.E.; et al. A Quantitative Release Assessment for the Noncommercial Movement of Companion Animals: Risk of Rabies Reintroduction to the United Kingdom. Risk Anal. 2012, 32, 1769–1783. [Google Scholar] [CrossRef] [PubMed]
- Hudson, E.G.; Brookes, V.J.; Ward, M.P. Assessing the Risk of a Canine Rabies Incursion in Northern Australia. Front. Vet. Sci. 2017, 4, 141. [Google Scholar] [CrossRef] [PubMed]
- Brookes, V.J.; Ward, M. Expert Opinion to Identify High-Risk Entry Routes of Canine Rabies into Papua New Guinea. Zoonoses Public Health 2016, 64, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Brookes, V.J.; Keponge-Yombo, A.; Thomson, D.; Ward, M.P. Risk assessment of the entry of canine-rabies into Papua New Guinea via sea and land routes. Prev. Vet. Med. 2017, 145, 49–66. [Google Scholar] [CrossRef]
- Gabriele-Rivet, V.; Arsenault, J.; Wilhelm, B.; Brookes, V.J.; Newsome, T.M.; Ward, M.P. A Scoping Review of Dingo and Wild-Living Dog Ecology and Biology in Australia to Inform Parameterisation for Disease Spread Modelling. Front. Vet. Sci. 2019, 6, 47. [Google Scholar] [CrossRef] [PubMed]
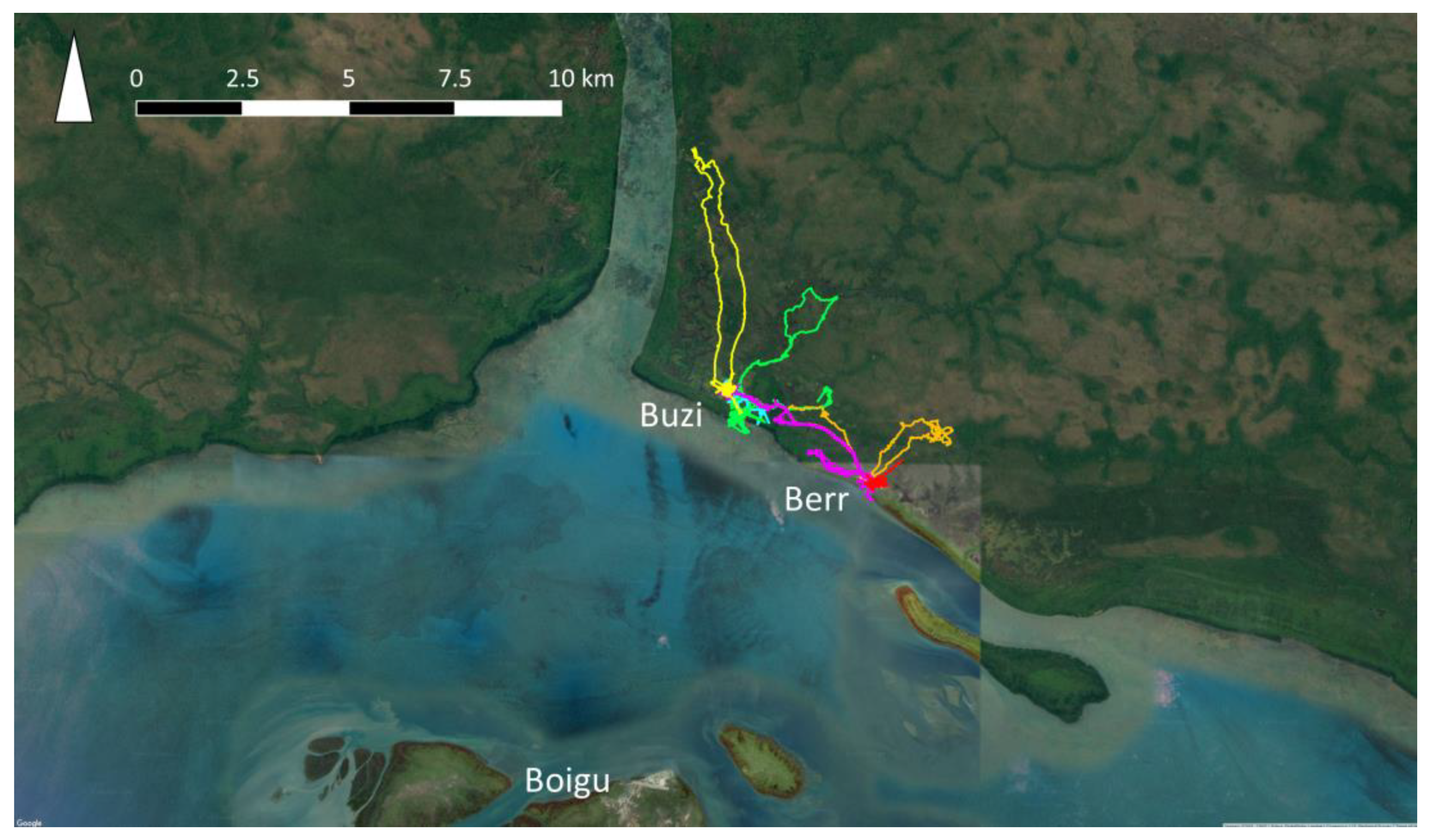
- Hudson, E.; Brookes, V.; Ward, M. Demographic studies of owned dogs in the Northern Peninsula Area, Australia, to inform population and disease management strategies. Aust. Vet. J. 2018, 96, 487–494. [Google Scholar] [CrossRef]
- Brookes, V.; VanderWaal, K.; Ward, M. The social networks of free-roaming domestic dogs in island communities in the Torres Strait, Australia. Prev. Vet. Med. 2018, 181, 104534. [Google Scholar] [CrossRef] [PubMed]
- Dürr, S.; Ward, M.P. Roaming behaviour and home range estimation of domestic dogs in Aboriginal and Torres Strait Islander communities in northern Australia using four different methods. Prev. Vet. Med. 2014, 117, 340–357. [Google Scholar] [CrossRef] [PubMed]
- Hudson, E.G.; Brookes, V.J.; Dürr, S.; Ward, M.P. Domestic dog roaming patterns in remote northern Australian indigenous communities and implications for disease modelling. Prev. Vet. Med. 2017, 146, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Gabriele-Rivet, V.; Arsenault, J.; Brookes, V.J.; Fleming, P.J.S.; Nury, C.; Ward, M.P. Dingo Density Estimates and Movements in Equatorial Australia: Spatially Explicit Mark–Resight Models. Anim. 2020, 10, 865. [Google Scholar] [CrossRef]
- Ward, M.P.; Sivieng, K.Q.-L.; Brookes, V. The dingo-domestic dog interface: Implications for disease spread. Aust. Zool. 2020. [Google Scholar] [CrossRef]
- Gabriele-Rivet, V.; Brookes, V.J.; Arsenault, J.; Ward, M.P. Hunting practices in northern Australia and their implication for disease transmission between community dogs and wild dogs. Aust. Vet. J. 2019, 97, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Bombara, C.; Dürr, S.; Gongora, J.; Ward, M. Roaming of dogs in remote Indigenous communities in northern Australia and potential interaction between community and wild dogs. Aust. Vet. J. 2017, 95, 182–188. [Google Scholar] [CrossRef]
- Bombara, C.B.; Dürr, S.; Machovsky-Capuska, G.E.; Jones, P.W.; Ward, M.P. A preliminary study to estimate contact rates between free-roaming domestic dogs using novel miniature cameras. PLoS ONE 2017, 12, e0181859. [Google Scholar] [CrossRef] [PubMed]
- Brookes, V.J.; Dürr, S.; Ward, M.P. Rabies-induced behavioural changes are key to rabies persistence in dog populations: Investigation using a network-based model. PLOS Negl. Trop. Dis. 2019, 13, e0007739. [Google Scholar] [CrossRef] [PubMed]
- Dürr, S.; Ward, M.P. Development of a Novel Rabies Simulation Model for Application in a Non-endemic Environment. PLOS Negl. Trop. Dis. 2015, 9, e0003876. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hudson, E.G.; Brookes, V.J.; Dürr, S.; Ward, M.P. Targeted pre-emptive rabies vaccination strategies in a susceptible domestic dog population with heterogeneous roaming patterns. Prev. Vet. Med. 2019, 172, 104774. [Google Scholar] [CrossRef] [PubMed]
- Gabriele-Rivet, V.; Ward, M.P.; Arsenault, J.; London, D.; Brookes, V.J. Could a rabies incursion spread in the northern Australian dingo population? Development of a spatial stochastic simulation model. PLOS Negl. Trop. Dis. 2021, 15, e0009124. [Google Scholar] [CrossRef]
- Johnstone-Robertson, S.P.; Fleming, P.J.S.; Ward, M.P.; Davis, S.A. Predicted Spatial Spread of Canine Rabies in Australia. PLOS Negl. Trop. Dis. 2017, 11, e0005312. [Google Scholar] [CrossRef]
- Sparkes, J.; McLeod, S.; Ballard, G.; Fleming, P.J.; Körtner, G.; Brown, W.Y. Rabies disease dynamics in naïve dog populations in Australia. Prev. Vet. Med. 2016, 131, 127–136. [Google Scholar] [CrossRef]
- Brookes, V.J.; Kennedy, E.; Dhagapan, P.; Ward, M.P. Qualitative research to design sustainable community-based surveillance for rabies in northern Australia and Papua New Guinea. Front. Vet. Sci. 2017, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Brookes, V.J.; Degeling, C.; Ward, M.P. Going viral in PNG—Exploring routes and circumstances of entry of a rabies-infected dog into Papua New Guinea. Soc. Sci. Med. 2018, 196, 10–18. [Google Scholar] [CrossRef]
- Degeling, C.; Brookes, V.; Lea, T.; Ward, M. Rabies response, One Health and more-than-human considerations in Indigenous communities in northern Australia. Soc. Sci. Med. 2018, 212, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Ward, M.; Brookes, V.; Hidayat, M.M.; Jatikusumah, A.; Abila, R. A study of human behavior and spread of rabies in West Kalimantan, Indonesia. In Proceedings of the The Conference of International Society for Economics and Social Science of Animal Health−Southeast Asia, Bogor, Indonesia, 17−18 October 2019. [Google Scholar]
- Brookes, V.; Ward, M.; Hidayat, M.M.; Jatikusumah, A.; Abila, R.; Yambo, A. Prioritisation of pathways for targeted risk assessment. In Proceedings of the Conference of International Society for Economics and Social Science of Animal Health−Southeast Asia, Bogor, Indonesia, 17−18 October 2019. [Google Scholar]
- Mustiana, A.; Toribio, J.-A.; Abdurrahman, M.; Suadnya, I.W.; Hernandez-Jover, M.; Putra, A.A.G.; Ward, M.P. Owned and Unowned Dog Population Estimation, Dog Management and Dog Bites to Inform Rabies Prevention and Response on Lombok Island, Indonesia. PLoS ONE 2015, 10, e0124092. [Google Scholar] [CrossRef]
- Halliday, J.; Daborn, C.; Auty, H.; Mtema, Z.; Lembo, T.; Bronsvoort, B.M.; Handel, I.; Knobel, D.; Hampson, K.; Cleaveland, S. Bringing together emerging and endemic zoonoses surveillance: Shared challenges and a common solution. Philos. Trans. R. Soc. Ser. B Biol. Sci. 2012, 367, 2872–2880. [Google Scholar] [CrossRef]
- Ward, M.P.; Kelman, M. Companion animal disease surveillance: A new solution to an old problem? Spat. Spatio-Temporal Epidemiol. 2011, 2, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Raynor, B.; Díaz, E.W.; Shinnick, J.; Zegarra, E.; Monroy, Y.; Mena, C.; Castillo-Neyra, R. The impact of the COVID-19 pandemic on rabies reemergence in Latin America: The case of Arequipa, Peru. medRxiv 2020. the preprint server for health sciences. [Google Scholar] [CrossRef]
- Rolfe, B. Leave No Fever Unresolved: The Malaria Pathway to End This Pandemic—And Prevent the Next; Malaria No More: Seattle, WA, USA; United Nations Foundation: New York, NY, USA, 2021; Available online: https://documentcloud.adobe.com/link/track?uri=urn%3Aaaid%3Ascds%3AUS%3Af58a2b97-173f-47fd-a168-2e2967469219 (accessed on 24 February 2021).



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ward, M.P.; Brookes, V.J. Rabies in Our Neighbourhood: Preparedness for an Emerging Infectious Disease. Pathogens 2021, 10, 375. https://doi.org/10.3390/pathogens10030375
Ward MP, Brookes VJ. Rabies in Our Neighbourhood: Preparedness for an Emerging Infectious Disease. Pathogens. 2021; 10(3):375. https://doi.org/10.3390/pathogens10030375
Chicago/Turabian StyleWard, Michael P., and Victoria J. Brookes. 2021. "Rabies in Our Neighbourhood: Preparedness for an Emerging Infectious Disease" Pathogens 10, no. 3: 375. https://doi.org/10.3390/pathogens10030375
APA StyleWard, M. P., & Brookes, V. J. (2021). Rabies in Our Neighbourhood: Preparedness for an Emerging Infectious Disease. Pathogens, 10(3), 375. https://doi.org/10.3390/pathogens10030375