
Targeting Gametocytes of the Malaria Parasite Plasmodium falciparum in a Functional Genomics Era: Next Steps




Abstract

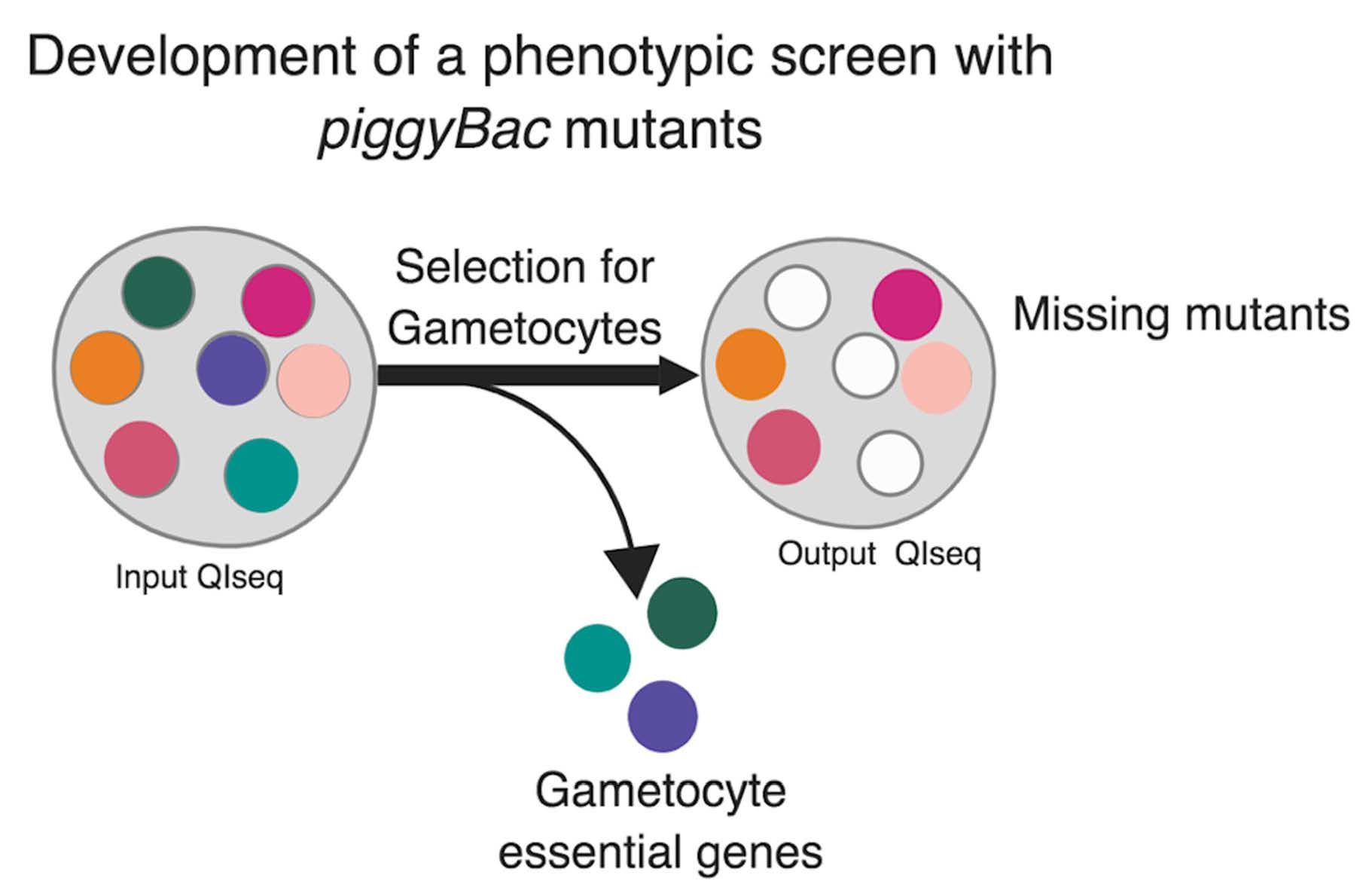{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Understanding P. falciparum Gametocyte Biology
2.1. Gametocyte Conversion
2.2. Gametocyte Sequestration
2.3. Gametocyte Development and Maturation
2.3.1. Molecular Biology
2.3.2. Metabolism
2.3.3. Cellular Biology
2.4. Gametocyte Sex Ratio and Exflagellation
3. Antimalarial Drug Therapies and Gametocytes
4. Transmission-Blocking Vaccines and Gametocytes
5. Identifying Gametocyte Essential Genes in the Age of Functional Genomics
6. Application of Forward Genetic Screens Using piggyBac Transposon Mutagenesis in Malaria Transmission
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2020: 20 Years of Global Progress and Challenges; World Health Organization: Geneva, Switzerland, 2020; ISBN 978-92-4-001579-1. [Google Scholar]
- Nkumama, I.N.; O’Meara, W.P.; Osier, F.H. Changes in Malaria Epidemiology in Africa and New Challenges for Elimination. Trends Parasitol. 2017, 33, 128–140. [Google Scholar] [CrossRef]
- Triglia, T.; Thompson, J.; Caruana, S.R.; Delorenzi, M.; Speed, T.; Cowman, A.F. Identification of Proteins from Plasmodium falciparum That Are Homologous to Reticulocyte Binding Proteins inPlasmodium vivax. Infect. Immun. 2001, 69, 1084–1092. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miller, L.H.; Baruch, D.I.; Marsh, K.; Doumbo, O.K. The pathogenic basis of malaria. Nature 2002, 415, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Wassmer, S.C.; Taylor, T.E.; Rathod, P.K.; Mishra, S.K.; Mohanty, S.; Arevalo-Herrera, M.; Duraisingh, M.T.; Smith, J.D. Investigating the Pathogenesis of Severe Malaria: A Multidisciplinary and Cross-Geographical Approach. Am. J. Trop. Med. Hyg. 2015, 93 (Suppl. S3), 42–56. [Google Scholar] [CrossRef]
- Bennink, S.; Kiesow, M.J.; Pradel, G. The development of malaria parasites in the mosquito midgut. Cell. Microbiol. 2016, 18, 905–918. [Google Scholar] [CrossRef]
- Smith, R.C.; Vega-Rodríguez, J.; Jacobs-Lorena, M. The Plasmodium bottleneck: Malaria parasite losses in the mosquito vector. Memorias do Instituto Oswaldo Cruz 2014, 109, 644–661. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, J.F.; Barreau, C.; Vernick, K.D. Efficiency of salivary gland invasion by malaria sporozoites is controlled by rapid sporozoite destruction in the mosquito haemocoel. Int. J. Parasitol. 2007, 37, 673–681. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, F.E.; Sirichaisinthop, J.; Miller, R.S.; Gasser, R.A.J.; Wongsrichanalai, C. Dependence of malaria detection and species diagnosis by microscopy on parasite density. Am. J. Trop. Med. Hyg. 2003, 69, 372–376. Available online: https://www.ncbi.nlm.nih.gov/pubmed/14640495 (accessed on 9 March 2021). [CrossRef] [PubMed]
- Bousema, T.; Okell, L.; Felger, I.; Drakeley, C. Asymptomatic malaria infections: Detectability, transmissibility and public health relevance. Nat. Rev. Genet. 2014, 12, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, R.N.; Drakeley, C.; Djimde, A.A.; Hall, B.F.; Hay, S.I.; Hemingway, J.; Kaslow, D.C.; Noor, A.; Okumu, F.; Steketee, R.; et al. malERA: An updated research agenda for malaria elimination and eradication. PLoS Med. 2017, 14, e1002456. [Google Scholar] [CrossRef] [PubMed]
- Ashley, E.A.; Dhorda, M.; Fairhurst, R.M.; Amaratunga, C.; Lim, P.; Suon, S.; Sreng, S.; Anderson, J.M.; Mao, S.; Sam, B.; et al. Spread of Artemisinin Resistance in Plasmodium falciparum Malaria. N. Engl. J. Med. 2014, 371, 411–423. [Google Scholar] [CrossRef]
- Noedl, H.; Se, Y.; Schaecher, K.; Smith, B.L.; Socheat, D.; Fukuda, M.M. Evidence of Artemisinin-Resistant Malaria in Western Cambodia. N. Engl. J. Med. 2008, 359, 2619–2620. [Google Scholar] [CrossRef] [PubMed]
- Ariey, F.; Witkowski, B.; Amaratunga, C.; Beghain, J.; Langlois, A.-C.; Khim, N.; Kim, S.; Duru, V.; Bouchier, C.; Ma, L.; et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 2013, 505, 50–55. [Google Scholar] [CrossRef]
- Uwimana, A.; Legrand, E.; Stokes, B.H.; Ndikumana, J.-L.M.; Warsame, M.; Umulisa, N.; Ngamije, D.; Munyaneza, T.; Mazarati, J.-B.; Munguti, K.; et al. Emergence and clonal expansion of in vitro artemisinin-resistant Plasmodium falciparum kelch13 R561H mutant parasites in Rwanda. Nat. Med. 2020, 26, 1602–1608. [Google Scholar] [CrossRef] [PubMed]
- Hancock, P.A.; Hendriks, C.J.M.; Tangena, J.-A.; Gibson, H.; Hemingway, J.; Coleman, M.; Gething, P.W.; Cameron, E.; Bhatt, S.; Moyes, C.L. Mapping trends in insecticide resistance phenotypes in African malaria vectors. PLoS Biol. 2020, 18, e3000633. [Google Scholar] [CrossRef]
- Moyes, C.L.; Athinya, D.K.; Seethaler, T.; Battle, K.E.; Sinka, M.; Hadi, M.P.; Hemingway, J.; Coleman, M.; Hancock, P.A. Evaluating insecticide resistance across African districts to aid malaria control decisions. Proc. Natl. Acad. Sci. USA 2020, 117, 22042–22050. [Google Scholar] [CrossRef] [PubMed]
- Wellems, T.E.; Plowe, C.V. Chloroquine-Resistant Malaria. J. Infect. Dis. 2001, 184, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Roper, C.; Pearce, R.; Nair, S.; Sharp, B.; Nosten, F.; Anderson, T. Intercontinental Spread of Pyrimethamine-Resistant Malaria. Science 2004, 305, 1124. [Google Scholar] [CrossRef]
- Nosten, F.; Van Vugt, M.; Price, R.; Luxemburger, C.; Thway, K.L.; Brockman, A.; Mcgready, R.; Ter Kuile, F.; Looareesuwan, S.; White, N.J. Effects of artesunate-mefloquine combination on incidence of Plasmodium falciparum malaria and mefloquine resistance in western Thailand: A prospective study. Lancet 2000, 356, 297–302. [Google Scholar] [CrossRef]
- Thriemer, K.; Van Hong, N.; Rosanas-Urgell, A.; Phuc, B.Q.; Ha, D.M.; Pockele, E.; Guetens, P.; Van Van, N.; Duong, T.T.; Amambua-Ngwa, A.; et al. Delayed Parasite Clearance after Treatment with Dihydroartemisinin-Piperaquine in Plasmodium falciparum Malaria Patients in Central Vietnam. Antimicrob. Agents Chemother. 2014, 58, 7049–7055. [Google Scholar] [CrossRef]
- Jaramillo-Gutierrez, G.; Rodrigues, J.; Ndikuyeze, G.; Povelones, M.; Molina-Cruz, A.; Barillas-Mury, C. Mosquito immune responses and compatibility between Plasmodium parasites and anopheline mosquitoes. BMC Microbiol. 2009, 9, 154. [Google Scholar] [CrossRef]
- Sinden, R.E.; Carter, R.; Drakeley, C.; Leroy, D. The biology of sexual development of Plasmodium: The design and implementation of transmission-blocking strategies. Malar. J. 2012, 11, 70. [Google Scholar] [CrossRef] [PubMed]
- Dyer, M.; Day, K. Commitment to Gametocytogenesis in Plasmodium falciparum. Parasitol. Today 2000, 16, 102–107. [Google Scholar] [CrossRef]
- Kafsack, B.F.C.; Rovira-Graells, N.; Clark, T.G.; Bancells, C.; Crowley, V.M.; Campino, S.G.; Williams, A.E.; Drought, L.G.; Kwiatkowski, D.P.; Baker, D.A.; et al. A transcriptional switch underlies commitment to sexual development in malaria parasites. Nature 2014, 507, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Hughes, K.R.; Modrzynska, K.K.; Otto, T.D.; Pfander, C.; Dickens, N.J.; Religa, A.A.; Bushell, E.; Graham, A.L.; Cameron, R.; et al. A cascade of DNA-binding proteins for sexual commitment and development in Plasmodium. Nature 2014, 507, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Bechtsi, D.; Waters, A. Genomics and epigenetics of sexual commitment in Plasmodium. Int. J. Parasitol. 2017, 47, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Bruce, M.C.; Alano, P.; Duthie, S.; Carter, R. Commitment of the malaria parasite Plasmodium falciparum to sexual and asexual development. Parasitology 1990, 100, 191–200. [Google Scholar] [CrossRef]
- Bancells, C.; Llorà-Batlle, O.; Poran, A.; Nötzel, C.; Rovira-Graells, N.; Elemento, O.; Kafsack, B.F.C.; Cortés, A. Revisiting the initial steps of sexual development in the malaria parasite Plasmodium falciparum. Nat. Microbiol. 2018, 4, 144–154. [Google Scholar] [CrossRef]
- Gautret, P.; Motard, A. Periodic infectivity of Plasmodium gametocytes to the vector. A review. Parasite 1999, 6, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Hawking, F.; Wilson, M.E.; Gammage, K. Evidence for cyclic development and short-lived maturity in the gametocytes of Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 1971, 65, 549–559. [Google Scholar] [CrossRef]
- Field, J.W.; Shute, P.G. The Microscopic Diagnosis of Human Malaria. II. A Morphological Study of the Erythroeytic Parasites. In Studies from the Institute for Medical Research, Federated Malay States; Government Press: Kuala Lumpur, Malaysia, 1956. [Google Scholar]
- Sinden, R.E. Gametocytogenesis of Plasmodium falciparum in vitro: An electron microscopic study. Parasitology 1982, 84, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Smalley, M.; Abdalla, S.; Brown, J. The distribution of Plasmodium falciparum in the peripheral blood and bone marrow of Gambian children. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 103–105. [Google Scholar] [CrossRef]
- Smalley, M.E.; Sinden, R.E. Plasmodium falciparum gametocytes: Their longevity and infectivity. Parasitology 1977, 74, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nixon, C.P. Plasmodium falciparum gametocyte transit through the cutaneous microvasculature: A new target for malaria transmission blocking vaccines? Hum. Vaccines Immunother. 2016, 12, 3189–3195. [Google Scholar] [CrossRef]
- Pick, C.; Ebersberger, I.; Spielmann, T.; Bruchhaus, I.; Burmester, T. Phylogenomic analyses of malaria parasites and evolution of their exported proteins. BMC Evol. Biol. 2011, 11, 167. [Google Scholar] [CrossRef] [PubMed]
- Waters, A.P.; Higgins, D.G.; McCutchan, T.F. Plasmodium falciparum appears to have arisen as a result of lateral transfer between avian and human hosts. Proc. Natl. Acad. Sci. USA 1991, 88, 3140–3144. [Google Scholar] [CrossRef] [PubMed]
- Ngotho, P.; Soares, A.B.; Hentzschel, F.; Achcar, F.; Bertuccini, L.; Marti, M. Revisiting gametocyte biology in malaria parasites. FEMS Microbiol. Rev. 2019, 43, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Drakeley, C.; Sutherland, C.; Bousema, J.T.; Sauerwein, R.W.; Targett, G.A. The epidemiology of Plasmodium falciparum gametocytes: Weapons of mass dispersion. Trends Parasitol. 2006, 22, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Shekalaghe, S.A.; Bousema, J.T.; Kunei, K.K.; Lushino, P.; Masokoto, A.; Wolters, L.R.; Mwakalinga, S.; Mosha, F.W.; Sauerwein, R.W.; Drakeley, C.J. Submicroscopic Plasmodium falciparum gametocyte carriage is common in an area of low and seasonal transmission in Tanzania. Trop. Med. Int. Health 2007, 12, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Ouédraogo, A.L.; Bousema, T.; Schneider, P.; De Vlas, S.J.; Ilboudo-Sanogo, E.; Cuzin-Ouattara, N.; Nébié, I.; Roeffen, W.; Verhave, J.P.; Luty, A.J.F.; et al. Substantial Contribution of Submicroscopical Plasmodium falciparum Gametocyte Carriage to the Infectious Reservoir in an Area of Seasonal Transmission. PLoS ONE 2009, 4, e8410. [Google Scholar] [CrossRef]
- Silvestrini, F.; Alano, P.; Williams, J.L. Commitment to the production of male and female gametocytes in the human malaria parasite Plasmodium falciparum. Parasitology 2000, 121, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Talman, A.M.; Domarle, O.; McKenzie, F.E.; Ariey, F.; Robert, V. Gametocytogenesis: The puberty of Plasmodium falciparum. Malar. J. 2004, 3, 24. [Google Scholar] [CrossRef] [PubMed]
- Maccallum, W. On the Flagellated Form of the Malarial Parasite. Lancet 1897, 150, 1240–1241. [Google Scholar] [CrossRef][Green Version]
- Janse, C.J.; Ponnudurai, T.; Lensen, A.H.W.; Meuwissen, J.H.E.T.; Ramesar, J.; Van Der Ploeg, M.; Overdulve, J.P. DNA synthesis in gametocytes of Plasmodium falciparum. Parasitology 1988, 96, 1–7. [Google Scholar] [CrossRef]
- Sinden, R.E.; Talman, A.; Marques, S.R.; Wass, M.N.; Sternberg, M.J.E. The flagellum in malarial parasites. Curr. Opin. Microbiol. 2010, 13, 491–500. [Google Scholar] [CrossRef]
- Carter, L.M.; Kafsack, B.F.; Llinás, M.; Mideo, N.; Pollitt, L.C.; Reece, S.E. Stress and sex in malaria parasites: Why does commitment vary? Evol. Med. Public Health 2013, 2013, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Sinden, R. Sexual Development of Malarial Parasites. Adv. Parasitol. 1983, 22, 153–216. [Google Scholar] [CrossRef] [PubMed]
- Graves, P.M.; McNeill, K.M.; Carter, R. Gametocyte Production in Cloned Lines of Plasmodium falciparum. Am. J. Trop. Med. Hyg. 1984, 33, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- Josling, G.A.; Llinás, M. Sexual development in Plasmodium parasites: Knowing when it’s time to commit. Nat. Rev. Genet. 2015, 13, 573–587. [Google Scholar] [CrossRef]
- Meibalan, E.; Marti, M. Biology of Malaria Transmission. Cold Spring Harb. Perspect. Med. 2016, 7, a025452. [Google Scholar] [CrossRef] [PubMed]
- Kengne-Ouafo, J.A.; Sutherland, C.J.; Binka, F.N.; Awandare, G.A.; Urban, B.C.; Dinko, B. Immune Responses to the Sexual Stages of Plasmodium falciparum Parasites. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Smalley, M.; Brown, J. Plasmodium falciparum gametocytogenesis stimulated by lymphocytes and serum from infected Gambian children. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 316–317. [Google Scholar] [CrossRef]
- Ono, T.; Nakai, T.; Nakabayashi, T. Induction of gametocytogenesis in Plasmodium falciparum by the culture supernatant of hybridoma cells producing anti-P. falciparum antibody. Biken J. 1986, 29, 77–81. Available online: https://www.ncbi.nlm.nih.gov/pubmed/3304278 (accessed on 9 March 2021). [PubMed]
- Nacher, M.; Singhasivanon, P.; Silachamroon, U.; Treeprasertsuk, S.; Tosukhowong, T.; Vannaphan, S.; Gay, F.; Mazier, D.; Looareesuwan, S. Decreased hemoglobin concentrations, hyperparasitemia, and severe malaria are associated with increased Plasmodium falciparum gametocyte carriage. J. Parasitol. 2002, 88, 97–101. [Google Scholar] [CrossRef]
- Gbotosho, G.O.; Sowunmi, A.; Okuboyejo, T.M.; Happi, C.T.; Michael, O.S.; Folarin, O.A.; Adewoye, E.O. Plasmodium falciparum gametocyte carriage, emergence, clearance and population sex ratios in anaemic and non-anaemic malarious children. Mem. Inst. Oswaldo Cruz 2011, 106, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Brancucci, N.M.; Gerdt, J.P.; Wang, C.; De Niz, M.; Philip, N.; Adapa, S.R.; Zhang, M.; Hitz, E.; Niederwieser, I.; Boltryk, S.D.; et al. Lysophosphatidylcholine Regulates Sexual Stage Differentiation in the Human Malaria Parasite Plasmodium falciparum. Cell 2017, 171, 1532–1544.e15. [Google Scholar] [CrossRef] [PubMed]
- Regev-Rudzki, N.; Wilson, D.W.; Carvalho, T.G.; Sisquella, X.; Coleman, B.M.; Rug, M.; Bursac, D.; Angrisano, F.; Gee, M.; Hill, A.F.; et al. Cell-Cell Communication between Malaria-Infected Red Blood Cells via Exosome-like Vesicles. Cell 2013, 153, 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Coleman, B.I.; Skillman, K.M.; Jiang, R.H.; Childs, L.M.; Altenhofen, L.M.; Ganter, M.; Leung, Y.; Goldowitz, I.; Kafsack, B.F.; Marti, M.; et al. A Plasmodium falciparum Histone Deacetylase Regulates Antigenic Variation and Gametocyte Conversion. Cell Host Microbe 2014, 16, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Brancucci, N.M.; Bertschi, N.L.; Zhu, L.; Niederwieser, I.; Chin, W.H.; Wampfler, R.; Freymond, C.; Rottmann, M.; Felger, I.; Bozdech, Z.; et al. Heterochromatin Protein 1 Secures Survival and Transmission of Malaria Parasites. Cell Host Microbe 2014, 16, 165–176. [Google Scholar] [CrossRef]
- Bui, H.T.N.; Passecker, A.; Brancucci, N.M.B.; Voss, T.S. Investigation of Heterochromatin Protein 1 Function in the Malaria Parasite Plasmodium falciparum Using a Conditional Domain Deletion and Swapping Approach. mSphere 2021, 6. [Google Scholar] [CrossRef]
- Filarsky, M.; Fraschka, S.A.; Niederwieser, I.; Brancucci, N.M.B.; Carrington, E.; Carrió, E.; Moes, S.; Jenoe, P.; Bártfai, R.; Voss, T.S. GDV1 induces sexual commitment of malaria parasites by antagonizing HP1-dependent gene silencing. Science 2018, 359, 1259–1263. [Google Scholar] [CrossRef] [PubMed]
- Tibúrcio, M.; Hitz, E.; Niederwieser, I.; Kelly, G.; Davies, H.; Doerig, C.; Billker, O.; Voss, T.S.; Treeck, M. GDV1 C-terminal truncation of 39 amino acids disrupts sexual commitment in Plasmodium falciparum. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kilejian, A. Characterization of a protein correlated with the production of knob-like protrusions on membranes of erythrocytes infected with Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 1979, 76, 4650–4653. [Google Scholar] [CrossRef]
- Baruch, D.I.; Pasloske, B.L.; Singh, H.B.; Bi, X.; Ma, X.C.; Feldman, M.; Taraschi, T.F.; Howard, R.J. Cloning the P. falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell 1995, 82, 77–87. [Google Scholar] [CrossRef]
- Thomson, J.; Robertson, A. The structure and development of Plasmodium falciparum gametocytes in the internal organs and peripheral circulation. Trans. R. Soc. Trop. Med. Hyg. 1935, 29, 31–40. [Google Scholar] [CrossRef]
- Aguilar, R.; Magallon-Tejada, A.; Achtman, A.H.; Moraleda, C.; Joice, R.; Cisteró, P.; Suen, C.S.N.L.W.; Nhabomba, A.; Macete, E.; Mueller, I.; et al. Molecular evidence for the localization of Plasmodium falciparum immature gametocytes in bone marrow. Blood 2014, 123, 959–966. [Google Scholar] [CrossRef]
- Joice, R.; Nilsson, S.K.; Montgomery, J.; Dankwa, S.; Egan, E.; Morahan, B.; Seydel, K.B.; Bertuccini, L.; Alano, P.; Williamson, K.C.; et al. Plasmodium falciparum transmission stages accumulate in the human bone marrow. Sci. Transl. Med. 2014, 6, 244re5. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, K.; Hentzschel, F.; Valkiūnas, G.; Marti, M. Plasmodium asexual growth and sexual development in the haematopoietic niche of the host. Nat. Rev. Genet. 2020, 18, 177–189. [Google Scholar] [CrossRef]
- Nemazee, D. Mechanisms of central tolerance for B cells. Nat. Rev. Immunol. 2017, 17, 281–294. [Google Scholar] [CrossRef]
- De Niz, M.; Meibalan, E.; Mejia, P.; Ma, S.; Brancucci, N.M.B.; Agop-Nersesian, C.; Mandt, R.; Ngotho, P.; Hughes, K.R.; Waters, A.P.; et al. Plasmodium gametocytes display homing and vascular transmigration in the host bone marrow. Sci. Adv. 2018, 4, eaat3775. [Google Scholar] [CrossRef]
- Tibúrcio, M.; Silvestrini, F.; Bertuccini, L.; Sander, A.F.; Turner, L.; Lavstsen, T.; Alano, P. Early gametocytes of the malaria parasite Plasmodium falciparum specifically remodel the adhesive properties of infected erythrocyte surface. Cell. Microbiol. 2012, 15, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Silvestrini, F.; Lasonder, E.; Olivieri, A.; Camarda, G.; van Schaijk, B.; Sanchez, M.; Younis, S.Y.; Sauerwein, R.; Alano, P. Protein Export Marks the Early Phase of Gametocytogenesis of the Human Malaria Parasite Plasmodium falciparum. Mol. Cell. Proteom. 2010, 9, 1437–1448. [Google Scholar] [CrossRef] [PubMed]
- Dantzler, K.W.; Ma, S.; Ngotho, P.; Stone, W.J.R.; Tao, D.; Rijpma, S.; De Niz, M.; Bark, S.K.N.; Jore, M.M.; Raaijmakers, T.K.; et al. Naturally acquired immunity against immature Plasmodium falciparum gametocytes. Sci. Transl. Med. 2019, 11, eaav3963. [Google Scholar] [CrossRef] [PubMed]
- Hermand, P.; Cicéron, L.; Pionneau, C.; Vaquero, C.; Combadiere, C.; Deterre, P. Plasmodium falciparum proteins involved in cytoadherence of infected erythrocytes to chemokine CX3CL1. Sci. Rep. 2016, 6, 33786. [Google Scholar] [CrossRef]
- Sinden, R.; Butcher, G.; Billker, O.; Fleck, S. Regulation of Infectivity of Plasmodium to the Mosquito Vector. Adv. Parasitol. 1996, 38, 53–117. [Google Scholar] [CrossRef] [PubMed]
- Raabe, A.C.; Billker, O.; Vial, H.J.; Wengelnik, K. Quantitative assessment of DNA replication to monitor microgametogenesis in Plasmodium berghei. Mol. Biochem. Parasitol. 2009, 168, 172–176. [Google Scholar] [CrossRef]
- Baker, D.A. Malaria gametocytogenesis. Mol. Biochem. Parasitol. 2010, 172, 57–65. [Google Scholar] [CrossRef]
- Canning, E.U.; Sinden, R.E. Nuclear organisation in gametocytes of Plasmodium and Hepatocystis: A cytochemical study. Parasitol. Res. 1975, 46, 297–299. [Google Scholar] [CrossRef]
- Sinden, R.E.; Canning, E.U.; Bray, R.S.; Smalley, M.E. Gametocyte and gamete development in Plasmodium falciparum. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1978, 201, 375–399. [Google Scholar] [CrossRef]
- Alano, P. Plasmodium falciparum gametocytes: Still many secrets of a hidden life. Mol. Microbiol. 2007, 66, 291–302. [Google Scholar] [CrossRef]
- Le Roch, K.G.; Johnson, J.R.; Florens, L.; Zhou, Y.; Santrosyan, A.; Grainger, M.; Yan, S.F.; Williamson, K.C.; Holder, A.A.; Carucci, D.J.; et al. Global analysis of transcript and protein levels across the Plasmodium falciparum life cycle. Genome Res. 2004, 14, 2308–2318. [Google Scholar] [CrossRef]
- Hall, N.; Karras, M.; Raine, J.D.; Carlton, J.M.; Kooij, T.W.A.; Berriman, M.; Florens, L.; Janssen, C.S.; Pain, A.; Christophides, G.K.; et al. A Comprehensive Survey of the Plasmodium Life Cycle by Genomic, Transcriptomic, and Proteomic Analyses. Science 2005, 307, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Lindner, S.; Miao, J. Translational regulation during stage transitions in malaria parasites. Ann. N. Y. Acad. Sci. 2014, 1342, 1–9. [Google Scholar] [CrossRef]
- Mair, G.R.; Braks, J.A.M.; Garver, L.S.; Wiegant, J.C.A.G.; Hall, N.; Dirks, R.W.; Khan, S.M.; Dimopoulos, G.; Janse, C.J.; Waters, A.P. Regulation of Sexual Development of Plasmodium by Translational Repression. Science 2006, 313, 667–669. [Google Scholar] [CrossRef]
- Mair, G.R.; Lasonder, E.; Garver, L.S.; Franke-Fayard, B.M.D.; Carret, C.K.; Wiegant, J.C.A.G.; Dirks, R.W.; Dimopoulos, G.; Janse, C.J.; Waters, A.P. Universal Features of Post-Transcriptional Gene Regulation Are Critical for Plasmodium Zygote Development. PLoS Pathog. 2010, 6, e1000767. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Fan, Q.; Parker, D.; Li, X.; Li, J.; Cui, L. Puf Mediates Translation Repression of Transmission-Blocking Vaccine Candidates in Malaria Parasites. PLoS Pathog. 2013, 9, e1003268. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Li, J.; Fan, Q.; Li, X.; Li, X.; Cui, L. The Puf-family RNA-binding protein PfPuf2 regulates sexual development and sex differentiation in the malaria parasite Plasmodium falciparum. J. Cell Sci. 2010, 123, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Bunnik, E.M.; Batugedara, G.; Saraf, A.; Prudhomme, J.; Florens, L.; Le Roch, K.G. The mRNA-bound proteome of the human malaria parasite Plasmodium falciparum. Genome Biol. 2016, 17, 147. [Google Scholar] [CrossRef]
- Rios, K.T.; Lindner, S.E. Protein–RNA interactions important for Plasmodium transmission. PLoS Pathog. 2019, 15, e1008095. [Google Scholar] [CrossRef]
- Creasey, A.; Mendis, K.; Carlton, J.; Williamson, N.; Wilson, I.; Carter, R. Maternal inheritance of extrachromosomal DNA in malaria parasites. Mol. Biochem. Parasitol. 1994, 65, 95–98. [Google Scholar] [CrossRef]
- Okamoto, N.; Spurck, T.P.; Goodman, C.D.; McFadden, G.I. Apicoplast and Mitochondrion in Gametocytogenesis of Plasmodium falciparum. Eukaryot. Cell 2008, 8, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Petmitr, S.; Krungkrai, J. Mitochondrial cytochrome b gene in two developmental stages of human malarial parasite Plasmodium falciparum. Southeast Asian J. Trop. Med. Public Health 1995, 26, 600–605. Available online: https://www.ncbi.nlm.nih.gov/pubmed/9139360 (accessed on 9 March 2021).
- MacRae, J.I.; Dixon, M.W.; Dearnley, M.K.; Chua, H.H.; Chambers, J.M.; Kenny, S.; Bottova, I.; Tilley, L.; McConville, M.J. Mitochondrial metabolism of sexual and asexual blood stages of the malaria parasite Plasmodium falciparum. BMC Biol. 2013, 11, 1–67. [Google Scholar] [CrossRef] [PubMed]
- Gulati, S.; Ekland, E.H.; Ruggles, K.V.; Chan, R.B.; Jayabalasingham, B.; Zhou, B.; Mantel, P.-Y.; Lee, M.C.S.; Spottiswoode, N.; Coburn-Flynn, O.; et al. Profiling the Essential Nature of Lipid Metabolism in Asexual Blood and Gametocyte Stages of Plasmodium falciparum. Cell Host Microbe 2015, 18, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.N.; Brown, S.H.J.; Rug, M.; Ridgway, M.C.; Mitchell, T.W.; Maier, A.G. Changes in lipid composition during sexual development of the malaria parasite Plasmodium falciparum. Malar. J. 2016, 15, 73. [Google Scholar] [CrossRef]
- Ben Mamoun, C.; Prigge, S.T.; Vial, H. Targeting the lipid metabolic pathways for the treatment of malaria. Drug Dev. Res. 2009, 71, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Dearnley, M.; Chu, T.; Zhang, Y.; Looker, O.; Huang, C.; Klonis, N.; Yeoman, J.; Kenny, S.; Arora, M.; Osborne, J.M.; et al. Reversible host cell remodeling underpins deformability changes in malaria parasite sexual blood stages. Proc. Natl. Acad. Sci. USA 2016, 113, 4800–4805. [Google Scholar] [CrossRef] [PubMed]
- Dixon, M.W.; Dearnley, M.K.; Hanssen, E.; Gilberger, T.; Tilley, L. Shape-shifting gametocytes: How and why does P. falciparum go banana-shaped? Trends Parasitol. 2012, 28, 471–478. [Google Scholar] [CrossRef] [PubMed]
- De Koning-Ward, T.F.; Gilson, P.R.; Boddey, J.A.; Rug, M.; Smith, B.J.; Papenfuss, A.T.; Sanders, P.R.; Lundie, R.J.; Maier, A.G.; Cowman, A.F.; et al. A newly discovered protein export machine in malaria parasites. Nature 2009, 459, 945–949. [Google Scholar] [CrossRef]
- Dearnley, M.K.; Yeoman, J.A.; Hanssen, E.; Kenny, S.; Turnbull, L.; Whitchurch, C.B.; Tilley, L.; Dixon, M.W.A. Origin, composition, organization and function of the inner membrane complex of Plasmodium falciparum gametocytes. J. Cell Sci. 2012, 125, 2053–2063. [Google Scholar] [CrossRef] [PubMed]
- Kono, M.; Herrmann, S.; Loughran, N.B.; Cabrera, A.; Engelberg, K.; Lehmann, C.; Sinha, D.; Prinz, B.; Ruch, U.; Heussler, V.; et al. Evolution and Architecture of the Inner Membrane Complex in Asexual and Sexual Stages of the Malaria Parasite. Mol. Biol. Evol. 2012, 29, 2113–2132. [Google Scholar] [CrossRef] [PubMed]
- Boucher, L.E.; Bosch, J. The apicomplexan glideosome and adhesins—Structures and function. J. Struct. Biol. 2015, 190, 93–114. [Google Scholar] [CrossRef] [PubMed]
- Neveu, G.; Lavazec, C. Erythrocyte Membrane Makeover by Plasmodium falciparum Gametocytes. Front. Microbiol. 2019, 10, 2652. [Google Scholar] [CrossRef]
- Tibúrcio, M.; Niang, M.; Deplaine, G.; Perrot, S.; Bischoff, E.; Ndour, P.A.; Silvestrini, F.; Khattab, A.; Milon, G.; David, P.H.; et al. A switch in infected erythrocyte deformability at the maturation and blood circulation of Plasmodium falciparum transmission stages. Blood 2012, 119, e172–e180. [Google Scholar] [CrossRef] [PubMed]
- Naissant, B.; Dupuy, F.; Duffier, Y.; Lorthiois, A.; Duez, J.; Scholz, J.; Buffet, P.; Merckx, A.; Bachmann, A.; Lavazec, C. Plasmodium falciparum STEVOR phosphorylation regulates host erythrocyte deformability enabling malaria parasite transmission. Blood 2016, 127, e42–e53. [Google Scholar] [CrossRef]
- Ramdani, G.; Naissant, B.; Thompson, E.; Breil, F.; Lorthiois, A.; Dupuy, F.; Cummings, R.; Duffier, Y.; Corbett, Y.; Mercereau-Puijalon, O.; et al. cAMP-Signalling Regulates Gametocyte-Infected Erythrocyte Deformability Required for Malaria Parasite Transmission. PLoS Pathog. 2015, 11, e1004815. [Google Scholar] [CrossRef]
- Smith, T.G.; Lourenço, P.; Carter, R.; Walliker, D.; Ranford-Cartwright, L.C. Commitment to sexual differentiation in the human malaria parasite, Plasmodium falciparum. Parasitology 2000, 121, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.G.; Walliker, D.; Ranford-Cartwright, L.C. Sexual differentiation and sex determination in the Apicomplexa. Trends Parasitol. 2002, 18, 315–323. [Google Scholar] [CrossRef]
- Creasey, A.M.; Ranford-Cartwright, L.C.; Moore, D.J.; Williamson, D.H.; Wilson, R.J.M.; Walliker, D.; Carter, R. Uniparental inheritance of the mitochondrial gene cytochrome b in Plasmodium falciparum. Curr. Genet. 1993, 23, 360–364. [Google Scholar] [CrossRef]
- Lasonder, E.; Rijpma, S.R.; Van Schaijk, B.C.; Hoeijmakers, W.A.; Kensche, P.R.; Gresnigt, M.S.; Italiaander, A.; Vos, M.W.; Woestenenk, R.; Bousema, T.; et al. Integrated transcriptomic and proteomic analyses of P. falciparum gametocytes: Molecular insight into sex-specific processes and translational repression. Nucleic Acids Res. 2016, 44, 6087–6101. [Google Scholar] [CrossRef]
- Schneider, P.; Reece, S.E.; Van Schaijk, B.C.; Bousema, T.; Lanke, K.H.; Meaden, C.S.; Gadalla, A.; Ranford-Cartwright, L.C.; Babiker, H.A. Quantification of female and male Plasmodium falciparum gametocytes by reverse transcriptase quantitative PCR. Mol. Biochem. Parasitol. 2015, 199, 29–33. [Google Scholar] [CrossRef]
- Santolamazza, F.; Avellino, P.; Siciliano, G.; Yao, F.A.; Lombardo, F.; Ouédraogo, J.B.; Modiano, D.; Alano, P.; Mangano, V.D. Detection of Plasmodium falciparum male and female gametocytes and determination of parasite sex ratio in human endemic populations by novel, cheap and robust RTqPCR assays. Malar. J. 2017, 16, 468. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.; Sawa, P.; Lanke, K.; Rijpma, S.; Oriango, R.; Nyaurah, M.; Osodo, P.; Osoti, V.; Mahamar, A.; Diawara, H.; et al. A Molecular Assay to Quantify Male and Female Plasmodium falciparum Gametocytes: Results From 2 Randomized Controlled Trials Using Primaquine for Gametocyte Clearance. J. Infect. Dis. 2017, 216, 457–467. [Google Scholar] [CrossRef]
- Meerstein-Kessel, L.; Andolina, C.; Carrio, E.; Mahamar, A.; Sawa, P.; Diawara, H.; Van De Vegte-Bolmer, M.; Stone, W.; Collins, K.A.; Schneider, P.; et al. A multiplex assay for the sensitive detection and quantification of male and female Plasmodium falciparum gametocytes. Malar. J. 2018, 17, 441. [Google Scholar] [CrossRef]
- Tadesse, F.G.; Meerstein-Kessel, L.; Gonçalves, B.P.; Drakeley, C.; Ranford-Cartwright, L.; Bousema, T. Gametocyte Sex Ratio: The Key to Understanding Plasmodium falciparum Transmission? Trends Parasitol. 2019, 35, 226–238. [Google Scholar] [CrossRef]
- Henry, N.B.; Sermé, S.S.; Siciliano, G.; Sombié, S.; Diarra, A.; Sagnon, N.; Traoré, A.S.; Sirima, S.B.; Soulama, I.; Alano, P. Biology of Plasmodium falciparum gametocyte sex ratio and implications in malaria parasite transmission. Malar. J. 2019, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sowunmi, A.; Balogun, S.T.; Gbotosho, G.O.; Happi, C.T. Plasmodium falciparum gametocyte sex ratios in children with acute, symptomatic, uncomplicated infections treated with amodiaquine. Malar. J. 2008, 7, 169. [Google Scholar] [CrossRef] [PubMed]
- Sowunmi, A.; Balogun, S.T.; Gbotosho, G.O.; Happi, C.T. Influence of anaemia on Plasmodium falciparum gametocyte sex ratios in acutely symptomatic children. Open Trop. Med. J. 2008, 1, 13–20. [Google Scholar] [CrossRef][Green Version]
- Reece, S.E.; Drew, D.R.; Gardner, A. Sex ratio adjustment and kin discrimination in malaria parasites. Nature 2008, 453, 609–614. [Google Scholar] [CrossRef]
- Sowunmi, A.; Fateye, B.A. Gametocyte sex ratios in children with asymptomatic, recrudescent, pyrimethamine-sulfadoxine-resistant, Plasmodium falciparummalaria. Ann. Trop. Med. Parasitol. 2003, 97, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Sowunmi, A.; Balogun, S.T.; Gbotosho, G.O.; Happi, C.T. Plasmodium falciparum gametocyte sex ratios in symptomatic children treated with antimalarial drugs. Acta Trop. 2009, 109, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Billker, O.; Alano, P. Molecular Approaches to Malaria. Mol. Microbiol. 2005, 219, 191–219. [Google Scholar]
- Nijhout, M.M.; Carter, R. Gamete development in malaria parasites: Bicarbonate-dependent stimulation by pH in vitro. Parasitology 1978, 76, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Garcia, G.E.; Wirtz, R.A.; Barr, J.R.; Woolfitt, A.; Rosenberg, R. Xanthurenic Acid Induces Gametogenesis in Plasmodium, the Malaria Parasite. J. Biol. Chem. 1998, 273, 12003–12005. [Google Scholar] [CrossRef]
- Kuehn, A.; Pradel, G. The Coming-Out of Malaria Gametocytes. J. Biomed. Biotechnol. 2010, 2010, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Delves, M.J.; Miguel-Blanco, C.; Matthews, H.; Molina, I.; Ruecker, A.; Yahiya, S.; Straschil, U.; Abraham, M.; León, M.L.; Fischer, O.J.; et al. A high throughput screen for next-generation leads targeting malaria parasite transmission. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Colmenarejo, G.; Lozano, S.; González-Cortés, C.; Calvo, D.; Sanchez-Garcia, J.; Matilla, J.-L.P.; Leroy, D.; Rodrigues, J. Predicting transmission blocking potential of anti-malarial compounds in the Mosquito Feeding Assay using Plasmodium falciparum Male Gamete Inhibition Assay. Sci. Rep. 2018, 8, 7764. [Google Scholar] [CrossRef]
- Delves, M.J.; Ruecker, A.; Straschil, U.; Lelièvre, J.; Marques, S.; López-Barragán, M.J.; Herreros, E.; Sinden, R.E. Male and Female Plasmodium falciparum Mature Gametocytes Show Different Responses to Antimalarial Drugs. Antimicrob. Agents Chemother. 2013, 57, 3268–3274. [Google Scholar] [CrossRef] [PubMed]
- Plowe, C.V. The evolution of drug-resistant malaria. Trans. R. Soc. Trop. Med. Hyg. 2009, 103 (Suppl. S1), S11–S14. [Google Scholar] [CrossRef]
- Mita, T.; Tanabe, K. Evolution of Plasmodium falciparum drug resistance: Implications for the development and containment of artemisinin resistance. Jpn. J. Infect. Dis. 2012, 65, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Delves, M.; Plouffe, D.; Scheurer, C.; Meister, S.; Wittlin, S.; Winzeler, E.A.; Sinden, R.E.; Leroy, D. The Activities of Current Antimalarial Drugs on the Life Cycle Stages of Plasmodium: A Comparative Study with Human and Rodent Parasites. PLoS Med. 2012, 9, e1001169. [Google Scholar] [CrossRef]
- Beaudoin, R.L.; Aikawa, M. Primaquine-Induced Changes in Morphology of Exoerythrocytic Stages of Malaria. Science 1968, 160, 1233–1234. [Google Scholar] [CrossRef] [PubMed]
- Lanners, H.N. Effect of the 8-aminoquinoline primaquine on culture-derived gametocytes of the malaria parasite Plasmodium falciparum. Parasitol. Res. 1991, 77, 478–481. [Google Scholar] [CrossRef]
- Camarda, G.; Jirawatcharadech, P.; Priestley, R.S.; Saif, A.; March, S.; Wong, M.H.L.; Leung, S.; Miller, A.B.; Baker, D.A.; Alano, P.; et al. Antimalarial activity of primaquine operates via a two-step biochemical relay. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Van Malderen, C.; Van Geertruyden, J.-P.; Machevo, S.; González, R.; Bassat, Q.; Talisuna, A.; Yeka, A.; Nabasumba, C.; Piola, P.; Daniel, A.; et al. Glucose-6-phosphate dehydrogenase deficiency, chlorproguanil-dapsone with artesunate and post-treatment haemolysis in African children treated for uncomplicated malaria. Malar. J. 2012, 11, 139. [Google Scholar] [CrossRef]
- Recht, J.; Ashley, E.A.; White, N.J. Use of primaquine and glucose-6-phosphate dehydrogenase deficiency testing: Divergent policies and practices in malaria endemic countries. PLoS Negl. Trop. Dis. 2018, 12, e0006230. [Google Scholar] [CrossRef] [PubMed]
- Krudsood, S.; Wilairatana, P.; Tangpukdee, N.; Chalermrut, K.; Srivilairit, S.; Thanachartwet, V.; Muangnoicharoen, S.; Luplertlop, N.; Brittenham, G.M.; Looareesuwan, S. Safety and tolerability of elubaquine (bulaquine, CDRI 80/53) for treatment of Plasmodium vivax malaria in Thailand. Korean J. Parasitol. 2006, 44, 221–228. [Google Scholar] [CrossRef][Green Version]
- Graves, P.M.; Choi, L.; Gelband, H.; Garner, P. Primaquine or other 8-aminoquinolines for reducing Plasmodium falciparum transmission. Cochrane Database Syst. Rev. 2018, 2, CD008152. [Google Scholar] [CrossRef] [PubMed]
- Adjalley, S.H.; Johnston, G.L.; Li, T.; Eastman, R.T.; Ekland, E.H.; Eappen, A.G.; Richman, A.; Sim, B.K.L.; Lee, M.C.S.; Hoffman, S.L.; et al. Quantitative assessment of Plasmodium falciparum sexual development reveals potent transmission-blocking activity by methylene blue. Proc. Natl. Acad. Sci. USA 2011, 108, E1214–E1223. [Google Scholar] [CrossRef] [PubMed]
- Wadi, I.; Pillai, C.R.; Anvikar, A.R.; Sinha, A.; Nath, M.; Valecha, N. Methylene blue induced morphological deformations in Plasmodium falciparum gametocytes: Implications for transmission-blocking. Malar. J. 2018, 17, 11. [Google Scholar] [CrossRef]
- Mandi, G.; Witte, S.; Meissner, P.; Coulibaly, B.; Mansmann, U.; Rengelshausen, J.; Schiek, W.; Jahn, A.; Sanon, M.; Wüst, K.; et al. Safety of the combination of chloroquine and methylene blue in healthy adult men with G6PD deficiency from rural Burkina Faso. Trop. Med. Int. Health 2005, 10, 32–38. [Google Scholar] [CrossRef]
- Coulibaly, B.; Zoungrana, A.; Mockenhaupt, F.P.; Schirmer, R.H.; Klose, C.; Mansmann, U.; Meissner, P.E.; Müller, O. Strong Gametocytocidal Effect of Methylene Blue-Based Combination Therapy against Falciparum Malaria: A Randomised Controlled Trial. PLoS ONE 2009, 4, e5318. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Nagbanshi, M.; Goldau, N.; Jorge, M.M.; Meissner, P.; Jahn, A.; Mockenhaupt, F.P.; Müller, O.; Lu, G.; Nagbanshi, M.; et al. Efficacy and safety of methylene blue in the treatment of malaria: A systematic review. BMC Med. 2018, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Price, R.; Nosten, F.; Luxemburger, C.; Ter Kuile, F.; Paiphun, L.; Chongsuphajaisiddhi, T.; White, N. Effects of artemisinin derivatives on malaria transmissibility. Lancet 1996, 347, 1654–1658. [Google Scholar] [CrossRef]
- Bousema, T.; Okell, L.; Shekalaghe, S.; Griffin, J.T.; Omar, S.; Sawa, P.; Sutherland, C.; Sauerwein, R.; Ghani, A.C.; Drakeley, C. Revisiting the circulation time of Plasmodium falciparum gametocytes: Molecular detection methods to estimate the duration of gametocyte carriage and the effect of gametocytocidal drugs. Malar. J. 2010, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Makanga, M. A review of the effects of artemether-lumefantrine on gametocyte carriage and disease transmission. Malar. J. 2014, 13, 291. [Google Scholar] [CrossRef] [PubMed]
- WWARN Gametocyte Study Group. Gametocyte carriage in uncomplicated Plasmodium falciparum malaria following treatment with artemisinin combination therapy: A systematic review and meta-analysis of individual patient data. BMC Med. 2016, 14, 1–18. [Google Scholar] [CrossRef]
- Meshnick, S.R. Artemisinin antimalarials: Mechanisms of action and resistance. Med. Trop. Rev. Corps Sante Colon. 1998, 58 (Suppl. S3), 13–17. [Google Scholar]
- Butterworth, A.S.; Skinner-Adams, T.S.; Gardiner, D.L.; Trenholme, K.R. Plasmodium falciparum gametocytes: With a view to a kill. Parasitology 2013, 140, 1718–1734. [Google Scholar] [CrossRef]
- Phyo, A.P.; Jittamala, P.; Nosten, F.H.; Pukrittayakamee, S.; Imwong, M.; White, N.J.; Duparc, S.; MacIntyre, F.; Baker, M.; Möhrle, J.J. Antimalarial activity of artefenomel (OZ439), a novel synthetic antimalarial endoperoxide, in patients with Plasmodium falciparum and Plasmodium vivax malaria: An open-label phase 2 trial. Lancet Infect. Dis. 2016, 16, 61–69. [Google Scholar] [CrossRef]
- Collins, K.A.; Abd-Rahman, A.N.; Marquart, L.; Ballard, E.; Gobeau, N.; Griffin, P.; Chalon, S.; Möhrle, J.J.; McCarthy, J.S. Antimalarial Activity of Artefenomel Against Asexual Parasites and Transmissible Gametocytes During Experimental Blood-Stage Plasmodium vivax Infection. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- McCarthy, J.S.; Rückle, T.; Elliott, S.L.; Ballard, E.; Collins, K.A.; Marquart, L.; Griffin, P.; Chalon, S.; Möhrle, J.J. A Single-Dose Combination Study with the Experimental Antimalarials Artefenomel and DSM265 To Determine Safety and Antimalarial Activity against Blood-StagePlasmodium falciparumin Healthy Volunteers. Antimicrob. Agents Chemother. 2019, 64. [Google Scholar] [CrossRef]
- Fleck, S.; Pudney, M.; Sinden, R. The effect of atovaquone (566C80) on the maturation and viability of Plasmodium falciparum gametocytes in vitro. Trans. R. Soc. Trop. Med. Hyg. 1996, 90, 309–312. [Google Scholar] [CrossRef]
- Ittarat, I.; Asawamahasakda, W.; Meshnick, S. The Effects of Antimalarials on the Plasmodium falciparum Dihydroorotate Dehydrogenase. Exp. Parasitol. 1994, 79, 50–56. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Siregar, J.E.; Kurisu, G.; Kobayashi, T.; Matsuzaki, M.; Sakamoto, K.; Mi-Ichi, F.; Watanabe, Y.-I.; Hirai, M.; Matsuoka, H.; Syafruddin, D.; et al. Direct evidence for the atovaquone action on the Plasmodium cytochrome bc 1 complex. Parasitol. Int. 2015, 64, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Korsinczky, M.; Chen, N.; Kotecka, B.; Saul, A.; Rieckmann, K.; Cheng, Q. Mutations in Plasmodium falciparum Cytochrome b That Are Associated with Atovaquone Resistance Are Located at a Putative Drug-Binding Site. Antimicrob. Agents Chemother. 2000, 44, 2100–2108. [Google Scholar] [CrossRef]
- Wojnarski, M.; Lon, C.; Vanachayangkul, P.; Gosi, P.; Sok, S.; Rachmat, A.; Harrison, D.; Berjohn, C.M.; Spring, M.; Chaoratanakawee, S.; et al. Atovaquone-Proguanil in Combination With Artesunate to Treat Multidrug-Resistant P. falciparum Malaria in Cambodia: An Open-Label Randomized Trial. Open Forum Infect. Dis. 2019, 6, ofz314. [Google Scholar] [CrossRef] [PubMed]
- Czesny, B.; Goshu, S.; Cook, J.L.; Williamson, K.C. The Proteasome Inhibitor Epoxomicin Has Potent Plasmodium falciparum Gametocytocidal Activity. Antimicrob. Agents Chemother. 2009, 53, 4080–4085. [Google Scholar] [CrossRef] [PubMed]
- Aminake, M.N.; Schoof, S.; Sologub, L.; Leubner, M.; Kirschner, M.; Arndt, H.-D.; Pradel, G. Thiostrepton and Derivatives Exhibit Antimalarial and Gametocytocidal Activity by Dually Targeting Parasite Proteasome and Apicoplast. Antimicrob. Agents Chemother. 2011, 55, 1338–1348. [Google Scholar] [CrossRef]
- Stocks, P.A.; Barton, V.; Antoine, T.; Biagini, G.A.; Ward, S.A.; O’Neill, P.M. Novel inhibitors of the Plasmodium falciparum electron transport chain. Parasitology 2014, 141, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Elliott, P.J.; Ross, J.S. The Proteasome: A new target for novel drug therapies. Am. J. Clin. Pathol. 2001, 116, 637–646. [Google Scholar] [CrossRef]
- Krishnan, K.M.; Williamson, K.C. The proteasome as a target to combat malaria: Hits and misses. Transl. Res. 2018, 198, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Moyo, P.; Mugumbate, G.; Eloff, J.N.; Louw, A.I.; Maharaj, V.J.; Birkholtz, L.-M. Natural Products: A Potential Source of Malaria Transmission Blocking Drugs? Pharmaceuticals 2020, 13, 251. [Google Scholar] [CrossRef]
- Hogh, B.; Dgedge, M.; Gamage-Mendis, A.; Barreto, J.; Butcher, G.A.; Eling, W.; Sinden, R.E.; Mendis, C.; Enosse, S.M.; Thompson, R.; et al. The differing impact of chloroquine and pyrimethamine/sulfadoxine upon the infectivity of malaria species to the mosquito vector. Am. J. Trop. Med. Hyg. 1998, 58, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Buckling, A.G.J.; Taylor, L.H.; Carlton, J.M.; Read, A.F. Adaptive changes in Plasmodium transmission strategies following chloroquine chemotherapy. Proc. R. Soc. B Biol. Sci. 1997, 264, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Von Seidlein, L.; Jawara, M.; Coleman, R.; Doherty, T.; Walraven, G.; Targett, G. Parasitaemia and gametocytaemia after treatment with chloroquine, pyrimethamine/sulfadoxine, and pyrimethamine/sulfadoxine combined with artesunate in young Gambians with uncomplicated malaria. Trop. Med. Int. Health 2001, 6, 92–98. [Google Scholar] [CrossRef]
- Tjitra, E.; Suprianto, S.; Anstey, N.M. Higher gametocyte prevalence following failure of treatment of Plasmodium falciparum malaria with sulfadoxine-pyrimethamine and the combination of chloroquine plus sulfadoxine-pyrimethamine: Implications for progression of anti-folate resistance. Trans. R. Soc. Trop. Med. Hyg. 2002, 96, 434–437. [Google Scholar] [CrossRef]
- Barnes, K.I.; Little, F.; Mabuza, A.; Mngomezulu, N.; Govere, J.; Durrheim, D.; Roper, C.; Watkins, B.; White, N.J. Increased Gametocytemia after Treatment: An Early Parasitological Indicator of Emerging Sulfadoxine-Pyrimethamine Resistance in Falciparum Malaria. J. Infect. Dis. 2008, 197, 1605–1613. [Google Scholar] [CrossRef] [PubMed]
- Ali, E.; MacKinnon, M.J.; Abdel-Muhsin, A.-M.A.; Ahmed, S.; Walliker, D.; Babiker, H.A. Increased density but not prevalence of gametocytes following drug treatment of Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 176–183. [Google Scholar] [CrossRef]
- Witmer, K.; Dahalan, F.A.; Delves, M.J.; Yahiya, S.; Watson, O.J.; Straschil, U.; Chiwcharoen, D.; Sornboon, B.; Pukrittayakamee, S.; Baum, J. Artemisinin-resistant malaria parasites show enhanced transmission to mosquitoes under drug pressure. bioRxiv 2020. [Google Scholar] [CrossRef]
- Carter, R. Transmission blocking malaria vaccines. Vaccine 2001, 19, 2309–2314. [Google Scholar] [CrossRef]
- Healer, J.; McGuinness, D.; Hopcroft, P.; Haley, S.; Carter, R.; Riley, E. Complement-mediated lysis of Plasmodium falciparum gametes by malaria-immune human sera is associated with antibodies to the gamete surface antigen Pfs230. Infect. Immun. 1997, 65, 3017–3023. [Google Scholar] [CrossRef]
- Ponnudurai, T.; Van Gemert, G.; Bensink, T.; Lensen, A.; Meuwissen, J. Transmission blockade of Plasmodium falciparum: Its variability with gametocyte numbers and concentration of antibody. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 491–493. [Google Scholar] [CrossRef]
- Sinden, R. A biologist’s perspective on malaria vaccine development. Hum. Vaccines 2010, 6, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Mendis, K.N.; Munesinghe, Y.D.; De Silva, Y.N.; Keragalla, I.; Carter, R. Malaria transmission-blocking immunity induced by natural infections of Plasmodium vivax in humans. Infect. Immun. 1987, 55, 369–372. [Google Scholar] [CrossRef]
- Stone, W.J.R.; Campo, J.J.; Ouédraogo, A.L.; Meerstein-Kessel, L.; Morlais, I.; Da, D.; Cohuet, A.; Nsango, S.; Sutherland, C.J.; Van De Vegte-Bolmer, M.; et al. Unravelling the immune signature of Plasmodium falciparum transmission-reducing immunity. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Acquah, F.K.; Adjah, J.; Williamson, K.C.; Amoah, L.E. Transmission-Blocking Vaccines: Old Friends and New Prospects. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.C.; Criscio, M.D.; Kaslow, D.C. Cloning and expression of the gene for Plasmodium falciparum transmission-blocking target antigen, Pfs230. Mol. Biochem. Parasitol. 1993, 58, 355–358. [Google Scholar] [CrossRef]
- Williamson, K.C. Pfs230: From malaria transmission-blocking vaccine candidate toward function. Parasite Immunol. 2003, 25, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Kaslow, D.C.; Quakyi, I.A.; Syin, C.; Raum, M.G.; Keister, D.B.; Coligan, J.E.; McCutchan, T.F.; Miller, L.H. A vaccine candidate from the sexual stage of human malaria that contains EGF-like domains. Nature 1988, 333, 74–76. [Google Scholar] [CrossRef]
- Eksi, S.; Czesny, B.; Van Gemert, G.-J.; Sauerwein, R.W.; Eling, W.; Williamson, K.C. Malaria transmission-blocking antigen, Pfs230, mediates human red blood cell binding to exflagellating male parasites and oocyst production. Mol. Microbiol. 2006, 61, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Burkhardt, M.; Nakuchima, S.; Herrera, R.; Muratova, O.; Gittis, A.G.; Kelnhofer, E.; Reiter, K.; Smelkinson, M.; Veltri, D.; et al. Structure and function of a malaria transmission blocking vaccine targeting Pfs230 and Pfs230-Pfs48/45 proteins. Commun. Biol. 2020, 3, 1–12. [Google Scholar] [CrossRef]
- Barr, P.J.; Green, K.M.; Gibson, H.L.; Bathurst, I.C.; Quakyi, I.A.; Kaslow, D.C. Recombinant pfs25 protein of Plasmodium fal-ciparum elicits malaria transmission-blocking immunity in experimental animals. J. Exp. Med. 1991, 174, 1203–1208. [Google Scholar] [CrossRef]
- Tomas, A.M.; Margos, G.; Dimopoulos, G.; Van Lin, L.H.; De Koning-Ward, T.F.; Sinha, R.; Lupetti, P.; Beetsma, A.L.; Rodriguez, M.C.; Karras, M.; et al. P25 and P28 proteins of the malaria ookinete surface have multiple and partially redundant functions. EMBO J. 2001, 20, 3975–3983. [Google Scholar] [CrossRef]
- Sagara, I.; Healy, S.A.; Assadou, M.H.; Gabriel, E.E.; Kone, M.; Sissoko, K.; Tembine, I.; Guindo, M.A.; Doucoure, M.; Niaré, K.; et al. Safety and immunogenicity of Pfs25H-EPA/Alhydrogel, a transmission-blocking vaccine against Plasmodium falciparum: A randomised, double-blind, comparator-controlled, dose-escalation study in healthy Malian adults. Lancet Infect. Dis. 2018, 18, 969–982. [Google Scholar] [CrossRef]
- Chichester, J.A.; Green, B.J.; Jones, R.M.; Shoji, Y.; Miura, K.; Long, C.A.; Lee, C.K.; Ockenhouse, C.F.; Morin, M.J.; Streatfield, S.J.; et al. Safety and immunogenicity of a plant-produced Pfs25 virus-like particle as a transmission blocking vaccine against malaria: A Phase 1 dose-escalation study in healthy adults. Vaccine 2018, 36, 5865–5871. [Google Scholar] [CrossRef] [PubMed]
- Talaat, K.R.; Ellis, R.D.; Hurd, J.; Hentrich, A.; Gabriel, E.; Hynes, N.A.; Rausch, K.M.; Zhu, D.; Muratova, O.; Herrera, R.; et al. Safety and Immunogenicity of Pfs25-EPA/Alhydrogel®, a Transmission Blocking Vaccine against Plasmodium falciparum: An Open Label Study in Malaria Naïve Adults. PLoS ONE 2016, 11, e0163144. [Google Scholar] [CrossRef] [PubMed]
- Mathias, D.K.; Plieskatt, J.L.; Armistead, J.S.; Bethony, J.M.; Abdul-Majid, K.B.; McMillan, A.; Angov, E.; Aryee, M.J.; Zhan, B.; Gillespie, P.; et al. Expression, Immunogenicity, Histopathology, and Potency of a Mosquito-Based Malaria Transmission-Blocking Recombinant Vaccine. Infect. Immun. 2012, 80, 1606–1614. [Google Scholar] [CrossRef]
- Armistead, J.S.; Morlais, I.; Mathias, D.K.; Jardim, J.G.; Joy, J.; Fridman, A.; Finnefrock, A.C.; Bagchi, A.; Plebanski, M.; Scorpio, D.G.; et al. Antibodies to a Single, Conserved Epitope in Anopheles APN1 Inhibit Universal Transmission of Plasmodium falciparum and Plasmodium vivax Malaria. Infect. Immun. 2013, 82, 818–829. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, S.C.; Armistead, J.S.; Mathias, D.K.; Sandeu, M.M.; Tao, D.; Borhani-Dizaji, N.; Tarimo, B.B.; Morlais, I.; Dinglasan, R.R.; Borg, N.A. The Anopheles-midgut APN1 structure reveals a new malaria transmission–blocking vaccine epitope. Nat. Struct. Mol. Biol. 2015, 22, 532–539. [Google Scholar] [CrossRef]
- Dinglasan, R.R.; Kalume, D.E.; Kanzok, S.M.; Ghosh, A.K.; Muratova, O.; Pandey, A.; Jacobs-Lorena, M. Disruption of Plasmodium falciparum development by antibodies against a conserved mosquito midgut antigen. Proc. Natl. Acad. Sci. USA 2007, 104, 13461–13466. [Google Scholar] [CrossRef] [PubMed]
- Kapulu, M.C.; Da, D.F.; Miura, K.; Li, Y.; Blagborough, A.M.; Churcher, T.S.; Nikolaeva, D.; Williams, A.; Goodman, A.L.; Sangare, I.; et al. Comparative Assessment of Transmission-Blocking Vaccine Candidates against Plasmodium falciparum. Sci. Rep. 2015, 5, 11193. [Google Scholar] [CrossRef]
- Saeed, M.; Roeffen, W.; Alexander, N.; Drakeley, C.J.; Targett, G.A.T.; Sutherland, C.J. Plasmodium falciparum Antigens on the Surface of the Gametocyte-Infected Erythrocyte. PLoS ONE 2008, 3, e2280. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, C.J. Surface antigens of Plasmodium falciparum gametocytes—A new class of transmission-blocking vaccine targets? Mol. Biochem. Parasitol. 2009, 166, 93–98. [Google Scholar] [CrossRef]
- Chan, J.-A.; Fowkes, F.J.I.; Beeson, J.G. Surface antigens of Plasmodium falciparum-infected erythrocytes as immune targets and malaria vaccine candidates. Cell. Mol. Life Sci. 2014, 71, 3633–3657. [Google Scholar] [CrossRef]
- Dinko, B.; King, E.; Targett, G.A.T.; Sutherland, C.J. Antibody responses to surface antigens of Plasmodium falciparum gametocyte-infected erythrocytes and their relation to gametocytaemia. Parasite Immunol. 2016, 38, 352–364. [Google Scholar] [CrossRef]
- Trager, W.; Jensen, J.B. Human malaria parasites in continuous culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef] [PubMed]
- Ifediba, T.; Vanderberg, J.P. Complete in vitro maturation of Plasmodium falciparum gametocytes. Nature 1981, 294, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Fivelman, Q.L.; McRobert, L.; Sharp, S.; Taylor, C.J.; Saeed, M.; Swales, C.A.; Sutherland, C.J.; Baker, D.A. Improved synchronous production of Plasmodium falciparum gametocytes in vitro. Mol. Biochem. Parasitol. 2007, 154, 119–123. [Google Scholar] [CrossRef]
- Khan, S.M.; Franke-Fayard, B.; Mair, G.R.; Lasonder, E.; Janse, C.J.; Mann, M.; Waters, A.P. Proteome Analysis of Separated Male and Female Gametocytes Reveals Novel Sex-Specific Plasmodium. Biol. Cell 2005, 121, 675–687. [Google Scholar] [CrossRef]
- Day, K.P.; Karamalis, F.; Thompson, J.; Barnes, D.A.; Peterson, C.; Brown, H.; Brown, G.V.; Kemp, D.J. Genes necessary for expression of a virulence determinant and for transmission of Plasmodium falciparum are located on a 0.3-megabase region of chromosome 9. Proc. Natl. Acad. Sci. USA 1993, 90, 8292–8296. [Google Scholar] [CrossRef] [PubMed]
- Alano, P.; Roca, L.; Smith, D.; Read, D.; Carter, R.; Day, K. Plasmodium falciparum: Parasites Defective in Early Stages of Gametocytogenesis. Exp. Parasitol. 1995, 81, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.L.; Dixon, M.W.; Spielmann, T.; Skinner-Adams, T.S.; Hawthorne, P.L.; Ortega, M.R.; Kemp, D.J.; Trenholme, K.R. Implication of a Plasmodium falciparum gene in the switch between asexual reproduction and gametocytogenesis. Mol. Biochem. Parasitol. 2005, 140, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Young, J.A.; Fivelman, Q.L.; Blair, P.L.; De La Vega, P.; Le Roch, K.G.; Zhou, Y.; Carucci, D.J.; Baker, D.A.; Winzeler, E.A. The Plasmodium falciparum sexual development transcriptome: A microarray analysis using ontology-based pattern identification. Mol. Biochem. Parasitol. 2005, 143, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Silvestrini, F.; Bozdech, Z.; Lanfrancotti, A.; Di Giulio, E.; Bultrini, E.; Picci, L.; DeRisi, J.L.; Pizzi, E.; Alano, P. Genome-wide identification of genes upregulated at the onset of gametocytogenesis in Plasmodium falciparum. Mol. Biochem. Parasitol. 2005, 143, 100–110. [Google Scholar] [CrossRef][Green Version]
- Llorà-Batlle, O.; Michel-Todó, L.; Witmer, K.; Toda, H.; Fernández-Becerra, C.; Baum, J.; Cortés, A. Conditional expression of PfAP2-G for controlled massive sexual conversion in Plasmodium falciparum. Sci. Adv. 2020, 6, eaaz5057. [Google Scholar] [CrossRef] [PubMed]
- Poran, A.; Nötzel, C.; Aly, O.; Mencia-Trinchant, N.; Harris, C.T.; Guzman, M.L.; Hassane, D.C.; Elemento, O.; Kafsack, B.F.C. Single-cell RNA sequencing reveals a signature of sexual commitment in malaria parasites. Nature 2017, 551, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Modrzynska, K.; Pfander, C.; Chappell, L.; Yu, L.; Suarez, C.; Dundas, K.; Gomes, A.R.; Goulding, D.; Rayner, J.C.; Choudhary, J.; et al. A Knockout Screen of ApiAP2 Genes Reveals Networks of Interacting Transcriptional Regulators Controlling the Plasmodium Life Cycle. Cell Host Microbe 2017, 21, 11–22. [Google Scholar] [CrossRef]
- Van Biljon, R.; Van Wyk, R.; Painter, H.J.; Orchard, L.; Reader, J.; Niemand, J.; Llinás, M.; Birkholtz, L.-M. Hierarchical transcriptional control regulates Plasmodium falciparum sexual differentiation. BMC Gen. 2019, 20, 1–16. [Google Scholar] [CrossRef]
- Hitz, E.; Balestra, A.C.; Brochet, M.; Voss, T.S. PfMAP-2 is essential for male gametogenesis in the malaria parasite Plasmodium falciparum. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Böhme, U.; Otto, T.D.; Sanders, M.; Newbold, C.I.; Berriman, M. Progression of the canonical reference malaria parasite genome from 2002–2019. Wellcome Open Res. 2019, 4, 58. [Google Scholar] [CrossRef]
- Sidik, S.M.; Huet, D.; Ganesan, S.M.; Huynh, M.-H.; Wang, T.; Nasamu, A.S.; Thiru, P.; Saeij, J.P.; Carruthers, V.B.; Niles, J.C.; et al. A Genome-wide CRISPR Screen in Toxoplasma Identifies Essential Apicomplexan Genes. Cell 2016, 166, 1423–1435.e12. [Google Scholar] [CrossRef]
- Sidik, S.M.; Huet, D.; Lourido, S. CRISPR-Cas9-based genome-wide screening of Toxoplasma gondii. Nat. Protoc. 2018, 13, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.R.; Bushell, E.; Schwach, F.; Girling, G.; Anar, B.; Quail, M.A.; Herd, C.; Pfander, C.; Modrzynska, K.; Rayner, J.C.; et al. A Genome-Scale Vector Resource Enables High-Throughput Reverse Genetic Screening in a Malaria Parasite. Cell Host Microbe 2015, 17, 404–413. [Google Scholar] [CrossRef]
- Bushell, E.; Gomes, A.R.; Sanderson, T.; Anar, B.; Girling, G.; Herd, C.; Metcalf, T.; Modrzynska, K.; Schwach, F.; Martin, R.E.; et al. Functional Profiling of a Plasmodium Genome Reveals an Abundance of Essential Genes. Cell 2017, 170, 260–272.e8. [Google Scholar] [CrossRef]
- Waldman, B.S.; Schwarz, D.; Wadsworth, M.H.; Saeij, J.P.; Shalek, A.K.; Lourido, S. Identification of a Master Regulator of Differentiation in Toxoplasma. Cell 2020, 180, 359–372.e16. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.J.; Hall, N.; Fung, E.; White, O.; Berriman, M.; Hyman, R.W.; Carlton, J.M.; Pain, A.; Nelson, K.E.; Bowman, S.; et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 2002, 419, 498–511. [Google Scholar] [CrossRef] [PubMed]
- Skinner-Adams, T.S.; Lawrie, P.M.; Hawthorne, P.L.; Gardiner, D.L.; Trenholme, K.R. Comparison of Plasmodium falciparum transfection methods. Malar. J. 2003, 2, 19. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Balu, B.; Chauhan, C.; Maher, S.P.; Shoue, D.A.; Kissinger, J.C.; Fraser, M.J.; Adams, J.H. piggyBac is an effective tool for functional analysis of the Plasmodium falciparumgenome. BMC Microbiol. 2009, 9, 83. [Google Scholar] [CrossRef] [PubMed]
- Crabb, B.S.; De Koning-Ward, T.F.; Gilson, P.R. Toward forward genetic screens in malaria-causing parasites using the piggyBac transposon. BMC Biol. 2011, 9, 21. [Google Scholar] [CrossRef]
- Munoz-Lopez, M. DNA Transposons: Nature and Applications in Genomics. Curr. Gen. 2010, 11, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, C.; Otto, T.D.; Oberstaller, J.; Liao, X.; Adapa, S.R.; Udenze, K.; Bronner, I.F.; Cassandra, D.; Mayho, M.; et al. Uncovering the essential genes of the human malaria parasitePlasmodium falciparumby saturation mutagenesis. Science 2018, 360, eaap7847. [Google Scholar] [CrossRef] [PubMed]
- Bronner, I.F.; Otto, T.D.; Zhang, M.; Udenze, K.; Wang, C.; Quail, M.A.; Jiang, R.H.; Adams, J.H.; Rayner, J.C. Quantitative insertion-site sequencing (QIseq) for high throughput phenotyping of transposon mutants. Genome Res. 2016, 26, 980–989. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Sedillo, J.; Oberstaller, J.; Li, S.; Zhang, M.; Singh, N.; Wang, C.C.Q.; Udenze, K.; Jiang, R.H.Y.; Adams, J.H. Phenotypic Screens Identify Parasite Genetic Factors Associated with Malarial Fever Response in Plasmodium falciparum piggyBac Mutants. mSphere 2016, 1, e00273-16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, C.; Oberstaller, J.; Thomas, P.; Otto, T.D.; Casandra, D.; Boyapalle, S.; Adapa, S.R.; Xu, S.; Adams, J.H. The endosymbiotic origins of the apicoplast link fever-survival and artemisinin-resistance in the malaria parasite. bioRxiv 2020. [Google Scholar] [CrossRef]
- Pradhan, A.; Siwo, G.H.; Singh, N.; Martens, B.; Balu, B.; Button-Simons, K.A.; Tan, A.; Zhang, M.; Udenze, K.O.; Jiang, R.H.; et al. Chemogenomic profiling of Plasmodium falciparum as a tool to aid antimalarial drug discovery. Sci. Rep. 2015, 5, 15930. [Google Scholar] [CrossRef] [PubMed]
- Ikadai, H.; Saliba, K.S.; Kanzok, S.M.; McLean, K.J.; Tanaka, T.Q.; Cao, J.; Williamson, K.C.; Jacobs-Lorena, M. Transposon mutagenesis identifies genes essential for Plasmodium falciparum gametocytogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, E1676–E1684. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chawla, J.; Oberstaller, J.; Adams, J.H. Targeting Gametocytes of the Malaria Parasite Plasmodium falciparum in a Functional Genomics Era: Next Steps. Pathogens 2021, 10, 346. https://doi.org/10.3390/pathogens10030346
Chawla J, Oberstaller J, Adams JH. Targeting Gametocytes of the Malaria Parasite Plasmodium falciparum in a Functional Genomics Era: Next Steps. Pathogens. 2021; 10(3):346. https://doi.org/10.3390/pathogens10030346
Chicago/Turabian StyleChawla, Jyotsna, Jenna Oberstaller, and John H. Adams. 2021. "Targeting Gametocytes of the Malaria Parasite Plasmodium falciparum in a Functional Genomics Era: Next Steps" Pathogens 10, no. 3: 346. https://doi.org/10.3390/pathogens10030346
APA StyleChawla, J., Oberstaller, J., & Adams, J. H. (2021). Targeting Gametocytes of the Malaria Parasite Plasmodium falciparum in a Functional Genomics Era: Next Steps. Pathogens, 10(3), 346. https://doi.org/10.3390/pathogens10030346