
Changes in Rice and Livestock Production and the Potential Emergence of Japanese Encephalitis in Africa


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
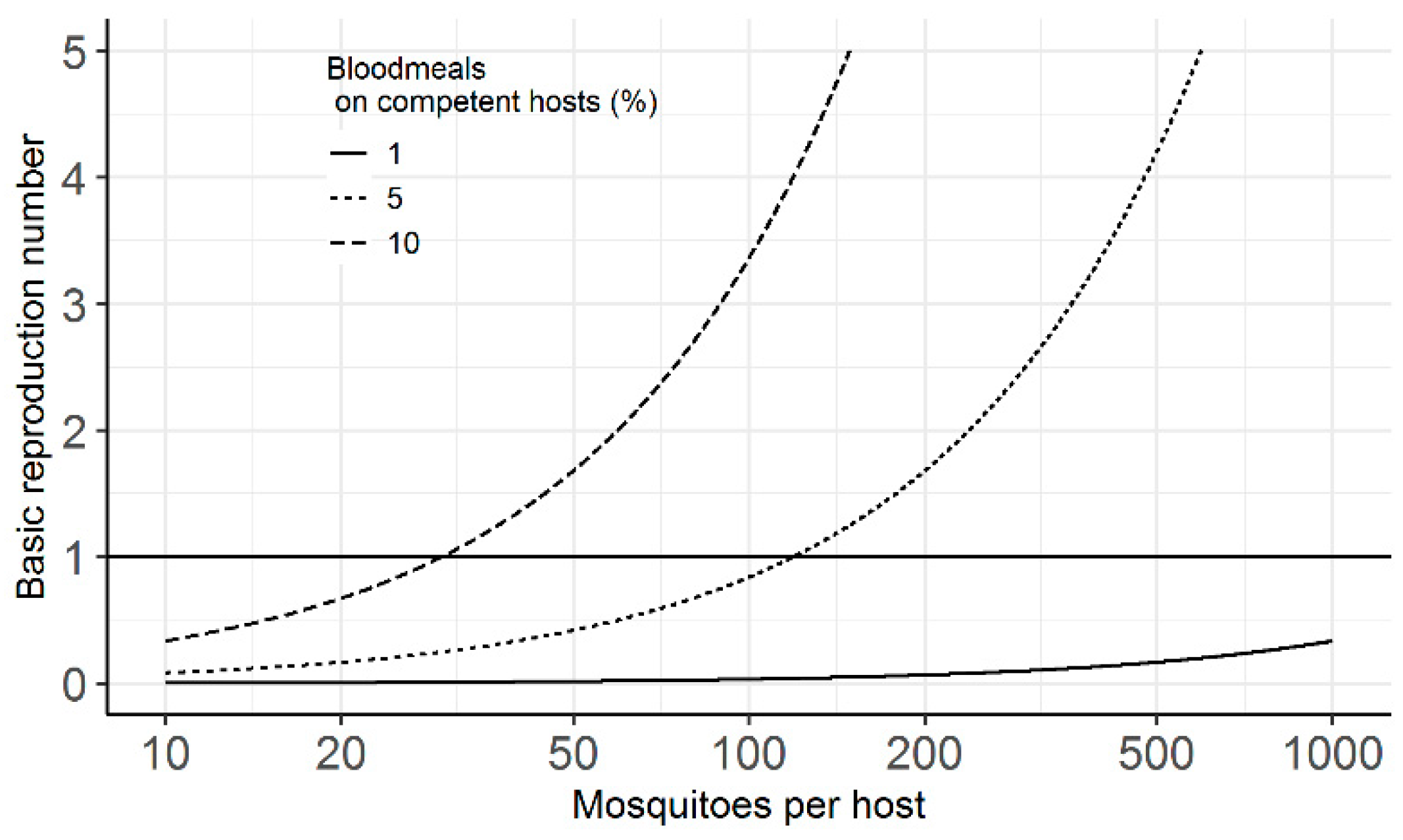
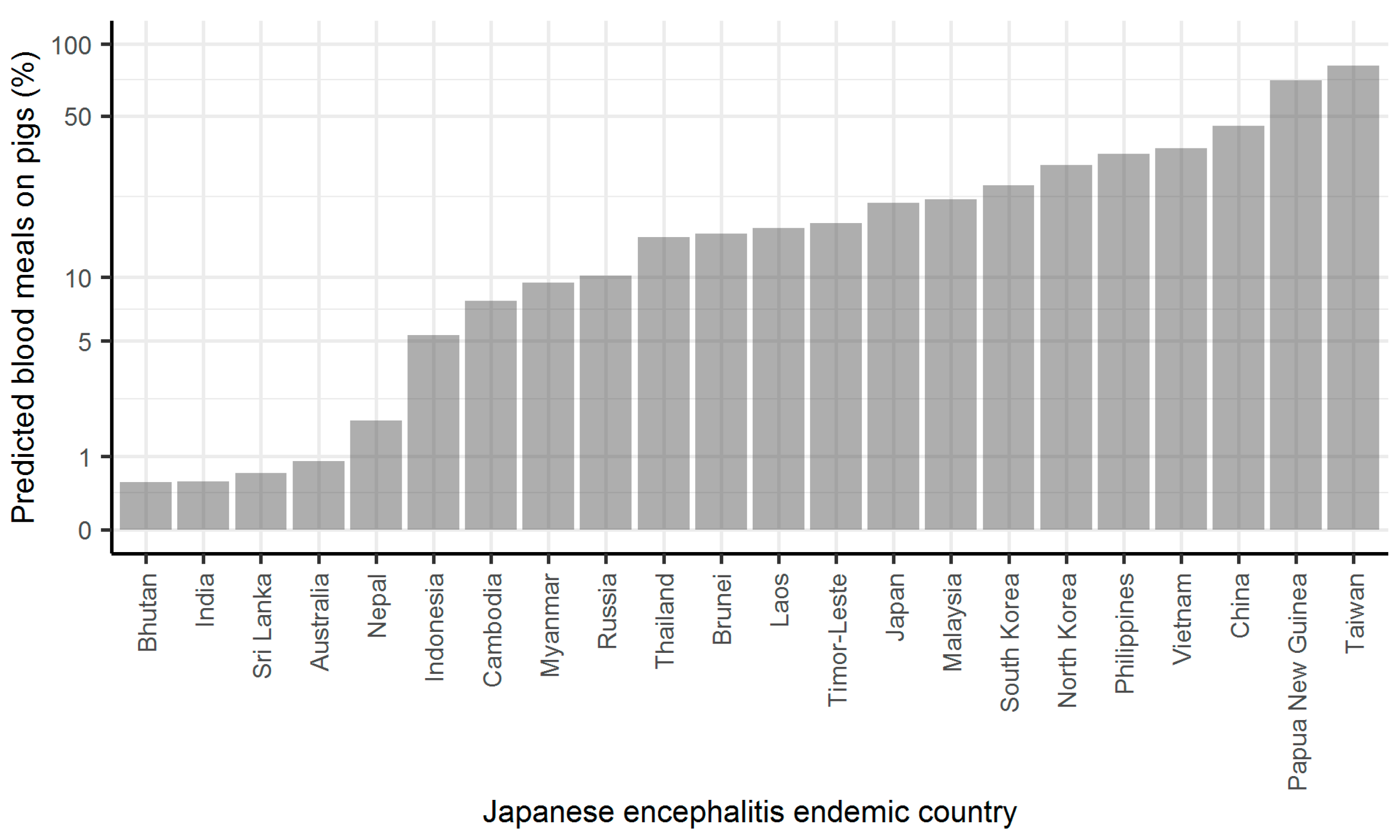
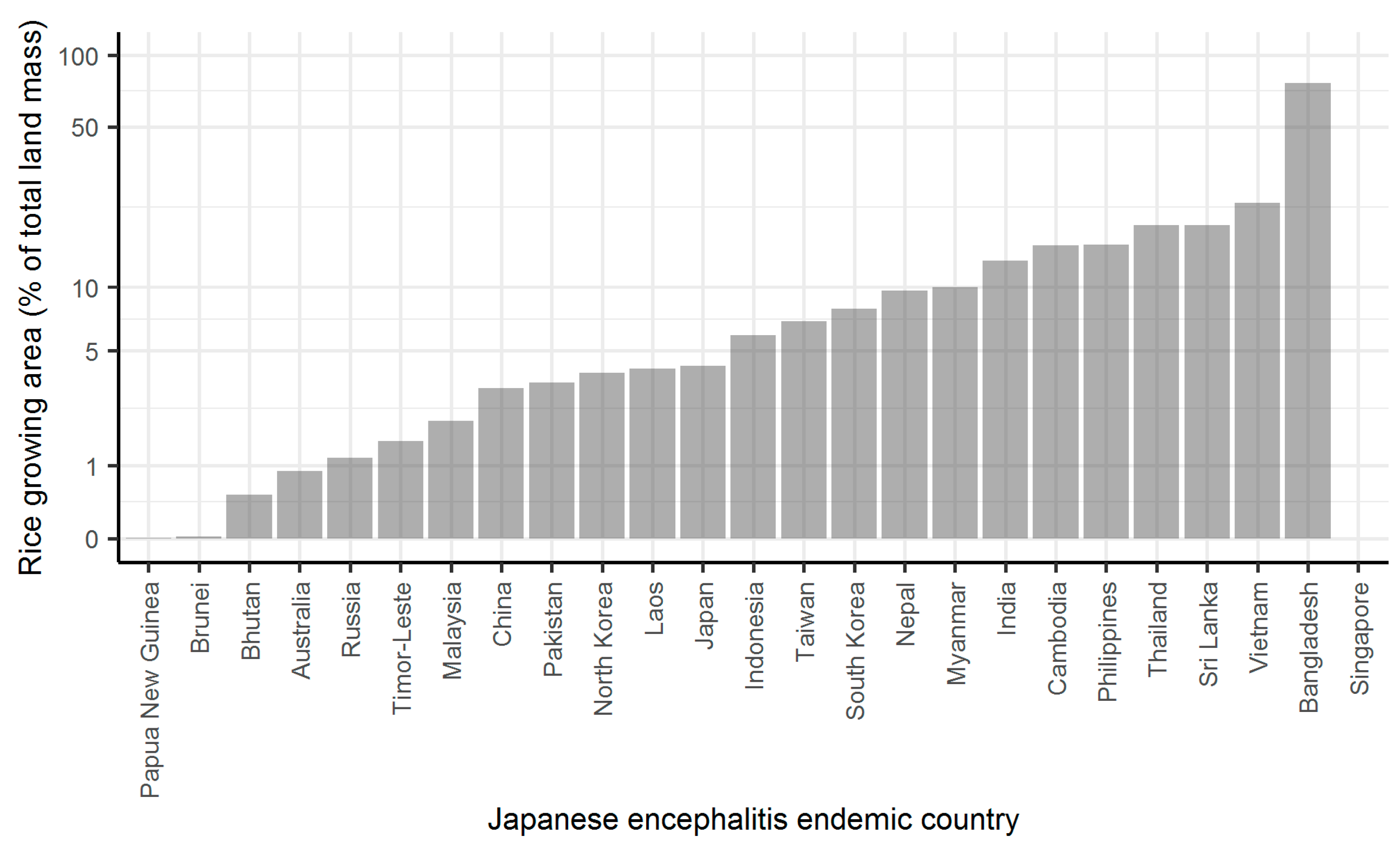
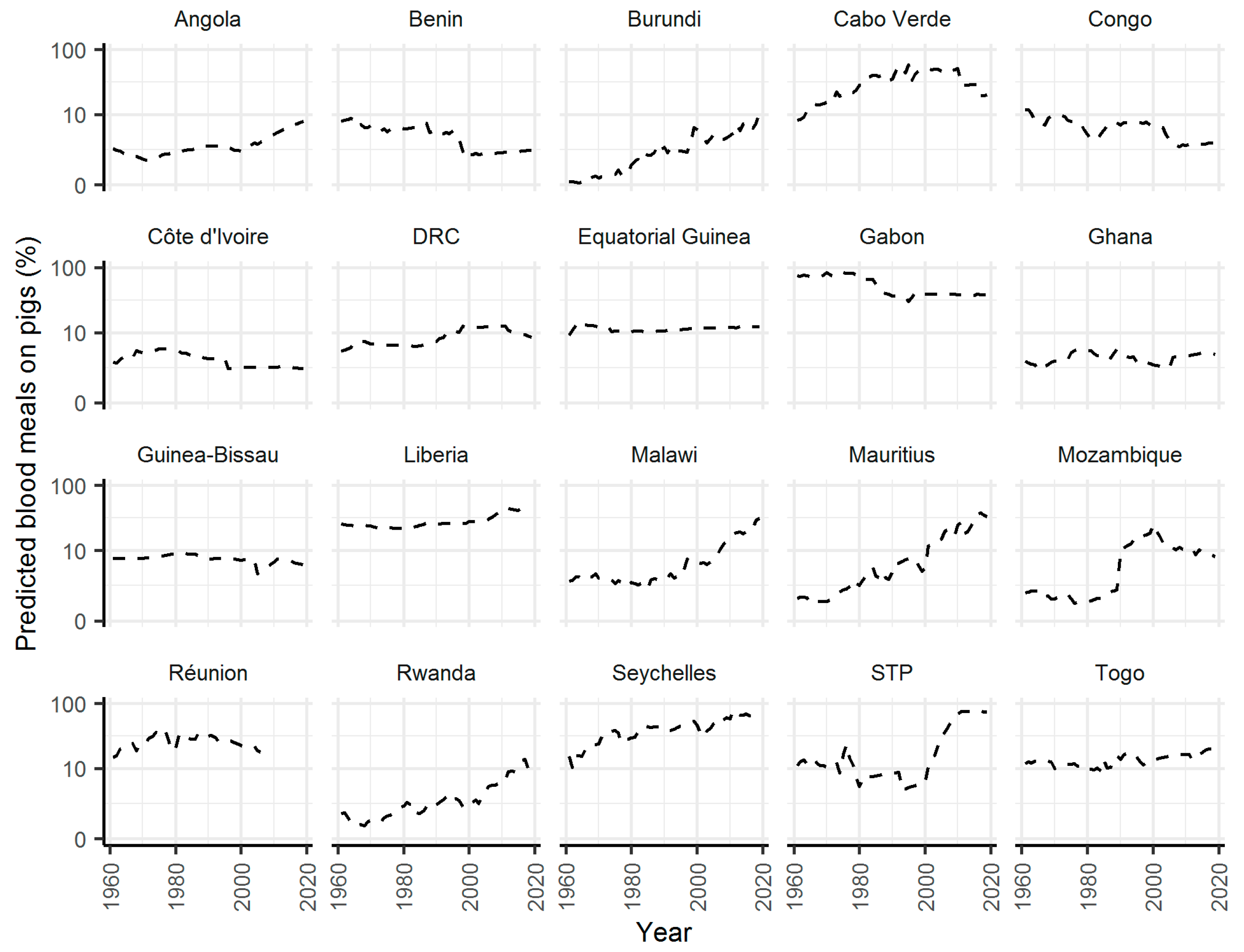
2. Results
3. Discussion
4. Materials and Methods
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Quan, T.M.; Thao, T.T.N.; Duy, N.M.; Nhat, T.M.; Clapham, H.E. Estimates of the global burden of japanese encephalitis and the impact of vaccination from 2000-2015. Elife 2020, 9, 1–187. [Google Scholar] [CrossRef]
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Platonov, A.E.; Rossi, G.; Karan, L.S.; Mironov, K.O.; Busani, L.; Rezza, G. Does the Japanese encephalitis virus (JEV) represent a threat for human health in Europe? Detection of JEV RNA sequences in birds collected in Italy. Eurosurveillance 2012, 17, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Simon-Loriere, E.; Faye, O.; Prot, M.; Casdemont, I.; Fall, G.; Fernandez-Garcia, M.; Diagne, M.; Kipela, J.; Fall, I.; Holmes, E.; et al. Autochthonous Japanese encephalitis with yellow fever coinfection in Africa. N. Engl. J. Med. 2017, 1386–1388. [Google Scholar] [CrossRef]
- Gao, X.; Liu, H.; Li, X.; Fu, S.; Cao, L.; Shao, N.; Zhang, W.; Wang, Q.; Lu, Z.; Lei, W.; et al. Changing Geographic Distribution of Japanese Encephalitis Virus Genotypes, 1935-2017. Vector-Borne Zoonotic Dis. 2019, 19, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Buescher, E.; Scherer, W. Ecologic studies of Japanese encephalitis virus in Japan. IX. Epidemiologic correlations and conclusions. Am. J. Trop. Med. Hyg. 1959, 8, 719–722. [Google Scholar] [CrossRef] [PubMed]
- Konno, J.; Endo, K.; Agatsuma, H.; Ishida, N. Cyclic outbreaks of japanese encephalitis among pigs and humans. Am. J. Epidemiol. 1966, 84, 292–300. [Google Scholar] [CrossRef]
- Keiser, J.; Maltese, M.F.; Erlanger, T.E.; Bos, R.; Tanner, M.; Singer, B.H.; Utzinger, J. Effect of irrigated rice agriculture on Japanese encephalitis, including challenges and opportunities for integrated vector management. Acta Trop. 2005, 95, 40–57. [Google Scholar] [CrossRef] [PubMed]
- Richards, E.E.; Masuoka, P.; Brett-Major, D.; Smith, M.; Klein, T.A.; Kim, H.C.; Anyamba, A.; Grieco, J. The relationship between mosquito abundance and rice field density in the Republic of Korea. Int. J. Health Geogr. 2010, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Suwonkerd, W.; Tsuda, Y.; Sugiyama, A.; Wada, Y. Effects of rice culture practices on the abundance of Culex mosquitoes (Diptera: Culicidae) in Northern Thailand. J. Med. Entomol. 1997, 34, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Cha, G.W.; Jeong, Y.E.; Lee, W.G.; Chang, K.S.; Roh, J.Y.; Yang, S.C.; Park, M.Y.; Park, C.; Shin, E.H. Detection of Japanese encephalitis virus genotype V in Culex orientalis and Culex pipiens (Diptera: Culicidae) in Korea. PLoS ONE 2015, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, J.; Choeung, R.; Peng, B.; Pring, L.; Pang, S.; Ong, S.; Sorn, S.; Tarantola, A.; Fontenille, D.; Cappelle, J. Comparison of the dynamics of Japanese encephalitis virus circulation in sentinel pigs between a rural and a peri-urban setting in Cambodia. PLoS Negl. Trop. Dis. 2018, 12, 1–18. [Google Scholar] [CrossRef]
- Le Flohic, G.; Porphyre, V.; Barbazan, P.; Gonzalez, J.-P. Review of climate, landscape, and viral genetics as drivers of the Japanese encephalitis virus ecology. PLoS Negl. Trop. Dis. 2013, 7, e2208. [Google Scholar] [CrossRef]
- Lord, J.S.; Gurley, E.S.; Pulliam, J.R.C. Rethinking Japanese encephalitis virus transmission: A framework for implicating host and vector species. PLoS Negl. Trop. Dis. 2015, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ladreyt, H.; Durand, B.; Dussart, P.; Chevalier, V. How central is the domestic pig in the epidemiological cycle of Japanese encephalitis virus? A review of scientific evidence and implications for disease control. Viruses 2019, 11, 949. [Google Scholar] [CrossRef]
- Walter Reed Biosystematics Unit. Culex Tritaeniorhynchus Species Page. Available online: https://www.wrbu.si.edu/vectorspecies/mosquitoes/tritaeniorhynchus (accessed on 14 February 2021).
- Mwandawiro, C.; Tuno, N.; Suwonkerd, W.; Tsuda, Y.; Yanagi, T.; Takagi, M. Host preference of Japanese encephalitis vectors in Chiangmai, Northern Thailand. Med. Entomol. Zool. 1999, 50, 323–333. [Google Scholar] [CrossRef]
- Ilkal, M.A.; Dhanda, V.; Rae, B.U.; George, S.; Mishra, A.C.; Prasanna, Y.; Gopall, S. Absence of viraemia in cattle after experimental infection with Japanese encephalitis virus. Trans. R. Soc. Trop. Med. Hyg. 1988, 82, 628–631. [Google Scholar] [CrossRef]
- Longbottom, J.; Browne, A.J.; Pigott, D.M.; Sinka, M.E.; Golding, N.; Hay, S.I.; Moyes, C.L.; Shearer, F.M. Mapping the spatial distribution of the Japanese encephalitis vector, Culex tritaeniorhynchus Giles, 1901 (Diptera: Culicidae) within areas of Japanese encephalitis risk. Parasites Vectors 2017, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.; Nicolas, G.; Cinardi, G.; Van Boeckel, T.P.; Vanwambeke, S.O.; Wint, G.R.W.; Robinson, T.P. Global distribution data for cattle, buffaloes, horses, sheep, goats, pigs, chickens and ducks in 2010. Sci. Data 2018, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- FAO. FAOSTAT Database. Available online: http://www.fao.org/faostat/en/#data/RF (accessed on 5 February 2018).
- Smith, D.L.; McKenzie, F.E. Statics and dynamics of malaria infection in Anopheles mosquitoes. Malar. J. 2004, 3, 13. [Google Scholar] [CrossRef]
- Khan, S.U.; Salje, H.; Hannan, A.; Islam, M.A.; Bhuyan, A.A.M.; Islam, M.A.; Rahman, M.Z.; Nahar, N.; Hossain, M.J.; Luby, S.P.; et al. Dynamics of Japanese Encephalitis Virus Transmission among Pigs in Northwest Bangladesh and the Potential Impact of Pig Vaccination. PLoS Negl. Trop. Dis. 2014, 8, e3166. [Google Scholar] [CrossRef]
- Buescher, E.; Scherer, W.; Rosenberg, M.; Gresser, I.; Hardy, J.L.; Bullock, H.R. Ecologic studies of Japanese encephalitis virus in Japan. II. Mosquito infection. Am. J. Trop. Med. Hyg. 1959, 8, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Gajanana, A.; Rajendran, R.; Samuel, P.P.; Thenmozhi, V.; Tsai, T.F.; Kimura-Kuroda, J.; Reuben, R. Japanese Encephalitis in South Arcot District, Tamil Nadu, India: A Three-Year Longitudinal Study of Vector Abundance and Infection Frequency. J. Med. Entomol. 1997, 34, 651–659. [Google Scholar] [CrossRef]
- Cleton, N.B.; Bosco-Lauth, A.; Page, M.J.; Bowen, R. A Age-related susceptibility to Japanese encephalitis virus in domestic ducklings and chicks. Am. J. Trop. Med. Hyg. 2014, 90, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Ladreyt, H.; Auerswald, H.; Tum, S.; Ken, S.; Heng, L.; In, S.; Lay, S.; Top, C.; Ly, S.; Duong, V.; et al. Comparison of Japanese encephalitis force of infection in pigs, poultry and dogs in cambodian villages. Pathogens 2020, 9, 719. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S.; Williams, D.T.; Smith, D.W. Japanese Encephalitis Virus: The Geographic Distribution, Incidence, and Spread of a Virus with a Propensity to Emerge in New Areas. Perspect. Med. Virol. 2006, 16, 201–268. [Google Scholar] [CrossRef]
- Sule, W.F.; Oluwayelu, D.O.; Hernández-Triana, L.M.; Fooks, A.R.; Venter, M.; Johnson, N. Epidemiology and ecology of West Nile virus in sub-Saharan Africa. Parasites Vectors 2018, 11, 1–10. [Google Scholar] [CrossRef]
- Oliveira, A.R.S.; Strathe, E.; Etcheverry, L.; Cohnstaedt, L.W.; Mcvey, D.S. Assessment of data on vector and host competence for Japanese encephalitis virus: A systematic review of the literature. Prev. Vet. Med. 2018, 154, 71–89. [Google Scholar] [CrossRef]
- Seo, H.J.; Kim, H.C.; Klein, T.A.; Ramey, A.M.; Lee, J.H.; Kyung, S.G.; Park, J.Y.; Cho, Y.S.; Cho, I.S.; Yeh, J.Y. Molecular Detection and Genotyping of Japanese Encephalitis Virus in Mosquitoes during a 2010 Outbreak in the Republic of Korea. PLoS ONE 2013, 8, 1–11. [Google Scholar] [CrossRef]
- Mordecai, E.A.; Ryan, S.J.; Caldwell, J.M.; Shah, M.M.; LaBeaud, A.D. Climate change could shift disease burden from malaria to arboviruses in Africa. Lancet Planet. Heal. 2020, 4, e416–e423. [Google Scholar] [CrossRef]
- Ministry of Agriculture and Forestry. National Rice Development Strategy (NRDS) July 2018 Executive Summary; Ministry of Agriculture and Forestry: Luanda, Angola, 2018; Available online: https://riceforafrica.net/images/pdf/NRDS_Eng_Angola_20180910.pdf (accessed on 28 February 2021).
- Ricklin, M.E.; Garcìa-Nicolàs, O.; Brechbühl, D.; Python, S.; Zumkehr, B.; Posthaus, H.; Oevermann, A.; Summerfield, A. Japanese encephalitis virus tropism in experimentally infected pigs. Vet. Res. 2016, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lord, J.S. Changes in Rice and Livestock Production and the Potential Emergence of Japanese Encephalitis in Africa. Pathogens 2021, 10, 294. https://doi.org/10.3390/pathogens10030294
Lord JS. Changes in Rice and Livestock Production and the Potential Emergence of Japanese Encephalitis in Africa. Pathogens. 2021; 10(3):294. https://doi.org/10.3390/pathogens10030294
Chicago/Turabian StyleLord, Jennifer S. 2021. "Changes in Rice and Livestock Production and the Potential Emergence of Japanese Encephalitis in Africa" Pathogens 10, no. 3: 294. https://doi.org/10.3390/pathogens10030294
APA StyleLord, J. S. (2021). Changes in Rice and Livestock Production and the Potential Emergence of Japanese Encephalitis in Africa. Pathogens, 10(3), 294. https://doi.org/10.3390/pathogens10030294