
Discovery of a Novel Member of the Carlavirus Genus from Soybean (Glycine max L. Merr.)
 , ,
, , {kind=link}
{kind=link}
Abstract
1. Introduction
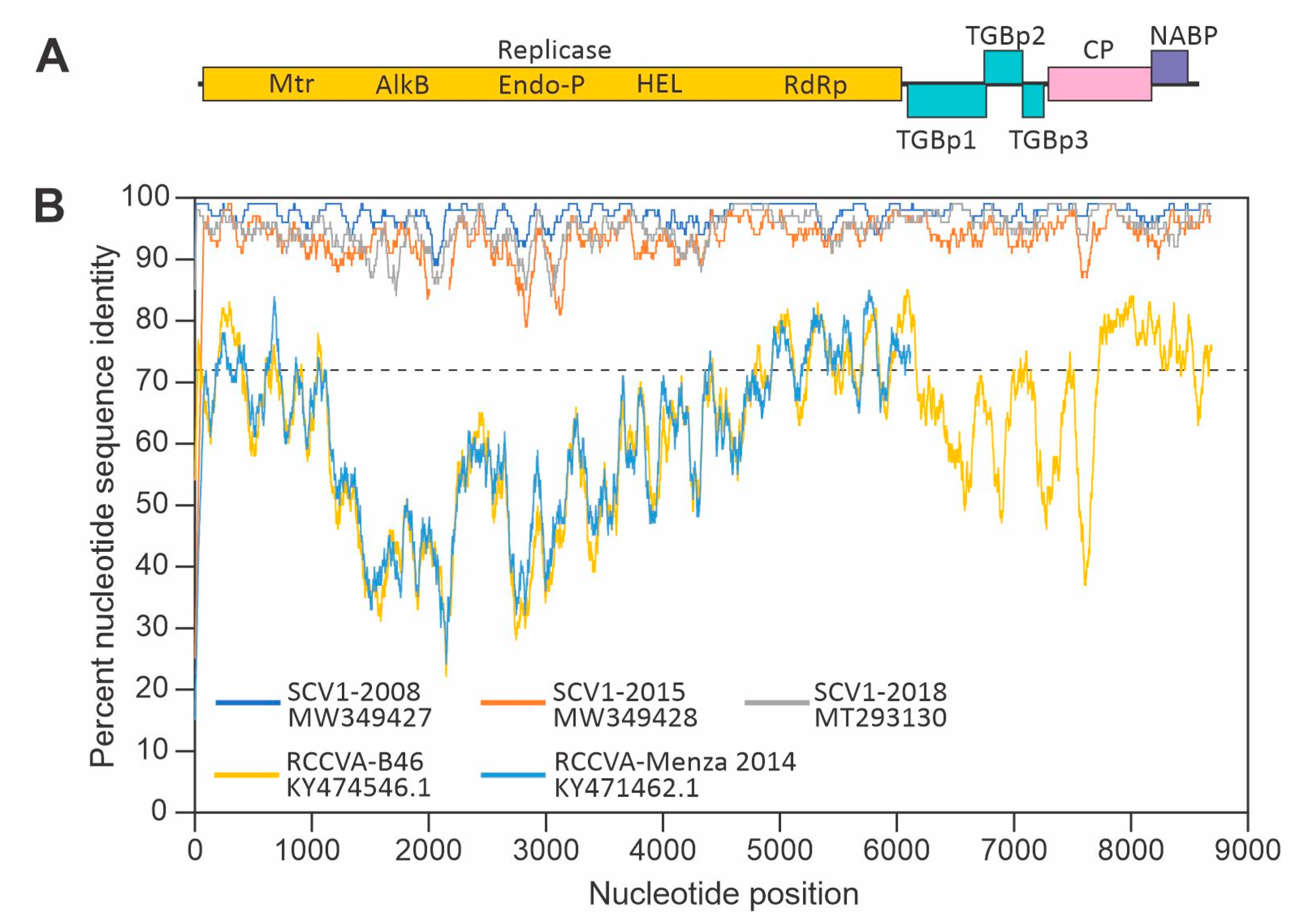
2. Results
2.1. High-Throughput Sequencing
2.2. Soybean Carlavirus 1 Genome Organization
2.3. Pylogenetic Analyses
3. Discussion
4. Materials and Methods
4.1. High-Throughput Sequencing
4.2. Random Amplification of cDNA ends (RACE)
4.3. Construction of Full-Length Clones of SCV1
4.4. Genome comparison
4.5. Phylogenetic Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Agricultural Statistics Service National Statistics for Soybeans. Available online: https://www.nass.usda.gov/Data_and_Statistics/ (accessed on 21 October 2020).
- Domier, L.L. Virus Diseases. In Compendium of Soybean Diseases, 5th ed.; Hartman, G.L., Rupe, J.C., Sikora, E.J., Domier, L.L., Davis, J.A., Steffey, K.L., Eds.; American Phytopathological Society Press: St. Paul, MN, USA, 2015; pp. 104–127. [Google Scholar]
- Zhou, J.; Tzanetakis, I.E. Soybean vein necrosis virus: An emerging virus in North America. Virus Genes 2019, 55, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Kreuze, J.F.; Pearson, M.N. Family Betaflexiviridae. In Virus Taxonomy—Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: New York, NY, USA, 2012; pp. 920–941. [Google Scholar]
- Morozov, S.Y.; Solovyev, A.G. Triple gene block: Modular design of a multifunctional machine for plant virus movement. J. Gen. Virol. 2003, 84, 1351–1366. [Google Scholar] [CrossRef] [PubMed]
- Thekke-Veetil, T.; Lagos-Kutz, D.; McCoppin, N.K.; Hartman, G.L.; Ju, H.-K.; Lim, H.-S.; Domier, L.L. Soybean thrips (Thysanoptera: Thripidae) harbor highly diverse populations of arthropod, fungal and plant viruses. Viruses 2020, 12, 1376. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.M.; Rozanov, M.N.; Hillman, B.I. Autocatalytic processing of the 223-kDa protein of blueberry scorch carlavirus by a papain-like proteinase. Virology 1995, 207, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Lukhovitskaya, N.I.; Ignatovich, I.V.; Savenkov, E.I.; Schiemann, J.; Morozov, S.Y.; Solovyev, A.G. Role of the zinc-finger and basic motifs of chrysanthemum virus B p12 protein in nucleic acid binding, protein localization and induction of a hypersensitive response upon expression from a viral vector. J. Gen. Virol. 2009, 90, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Gramstat, A.; Courtpozanis, A.; Rohde, W. The 12 kDa protein of potato virus M displays properties of a nucleic acid-binding regulatory protein. FEBS Lett. 1990, 276, 34–38. [Google Scholar] [CrossRef]
- Senshu, H.; Yamaji, Y.; Minato, N.; Shiraishi, T.; Maejima, K.; Hashimoto, M.; Miura, C.; Neriya, Y.; Namba, S. A dual strategy for the suppression of host antiviral silencing: Two distinct suppressors for viral replication and viral movement encoded by potato virus M. J. Virol. 2011, 85, 10269–10278. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; Junglen, S.; et al. Changes to virus taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2019). Arch. Virol. 2019, 164, 2417–2429. [Google Scholar] [CrossRef] [PubMed]
- Laguna, I.G.; Arneodo, J.D.; Rodríguez-Pardina, P.; Fiorona, M. Cowpea mild mottle virus infecting soybean crops in northwestern Argentina. Fitopatol. Bras. 2006, 31, 317. [Google Scholar] [CrossRef]
- Rosario, K.; Capobianco, H.; Ng, T.F.F.; Breitbart, M.; Polston, J.E. RNA viral metagenome of whiteflies leads to the discovery and characterization of a whitefly-transmitted carlavirus in North America. PLoS ONE 2014, 9, e86748. [Google Scholar] [CrossRef] [PubMed]
- Ryu, K.H.; Lee, B.Y. Carlavirus . In Encyclopedia of Virology, 3rd ed.; Mahy, B.W.J., van Regenmortel, M.H.V., Eds.; Academic Press: Cambridge, MA, USA, 2008; pp. 448–452. [Google Scholar]
- Nagata, T.; Alves, D.M.; Inoue-Nagata, A.K.; Tian, T.Y.; Kitajima, E.W.; Cardoso, J.E.; de Avila, A.C. A novel melon flexivirus transmitted by whitefly. Arch. Virol. 2005, 150, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Zanardo, L.G.; Carvalho, C.M. Cowpea mild mottle virus (Carlavirus, Betaflexiviridae): A review. Trop. Plant Pathol. 2017, 42, 417–430. [Google Scholar] [CrossRef]
- Martin, R.R.; Bristow, P.R. A carlavirus associated with blueberry scorch disease. Phytopathology 1988, 78, 1636–1640. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tinland, B.; Koukolikova-Nicola, Z.; Hall, M.N.; Hohn, B. The T-DNA-linked VirD2 protein contains two distinct functional nuclear localization signals. Proc. Natl. Acad. Sci. USA 1992, 89, 7442–7446. [Google Scholar] [CrossRef]
- Pattnaik, A.K.; Ball, L.A.; LeGrone, A.W.; Wertz, G.W. Infectious defective interfering particles of VSV from transcripts of a cDNA clone. Cell 1992, 69, 1011–1020. [Google Scholar] [CrossRef]
- Scholthof, H.B. Rapid delivery of foreign genes into plants by direct rub-inoculation with intact plasmid DNA of a tomato bushy stunt virus gene vector. J. Virol. 1999, 73, 7823–7829. [Google Scholar] [CrossRef] [PubMed]
- Greener, A. Expand your library by retrieving toxic clones with ABLE strains. Strategies 1993, 6, 7–9. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evolut. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thekke-Veetil, T.; McCoppin, N.K.; Hobbs, H.A.; Hartman, G.L.; Lambert, K.N.; Lim, H.-S.; Domier, L.L. Discovery of a Novel Member of the Carlavirus Genus from Soybean (Glycine max L. Merr.). Pathogens 2021, 10, 223. https://doi.org/10.3390/pathogens10020223
Thekke-Veetil T, McCoppin NK, Hobbs HA, Hartman GL, Lambert KN, Lim H-S, Domier LL. Discovery of a Novel Member of the Carlavirus Genus from Soybean (Glycine max L. Merr.). Pathogens. 2021; 10(2):223. https://doi.org/10.3390/pathogens10020223
Chicago/Turabian StyleThekke-Veetil, Thanuja, Nancy K. McCoppin, Houston A. Hobbs, Glen L. Hartman, Kris N. Lambert, Hyoun-Sub Lim, and Leslie. L. Domier. 2021. "Discovery of a Novel Member of the Carlavirus Genus from Soybean (Glycine max L. Merr.)" Pathogens 10, no. 2: 223. https://doi.org/10.3390/pathogens10020223
APA StyleThekke-Veetil, T., McCoppin, N. K., Hobbs, H. A., Hartman, G. L., Lambert, K. N., Lim, H.-S., & Domier, L. L. (2021). Discovery of a Novel Member of the Carlavirus Genus from Soybean (Glycine max L. Merr.). Pathogens, 10(2), 223. https://doi.org/10.3390/pathogens10020223