
Assessing Trypanosoma cruzi Parasite Diversity through Comparative Genomics: Implications for Disease Epidemiology and Diagnostics



Abstract
1. Introduction
2. Results
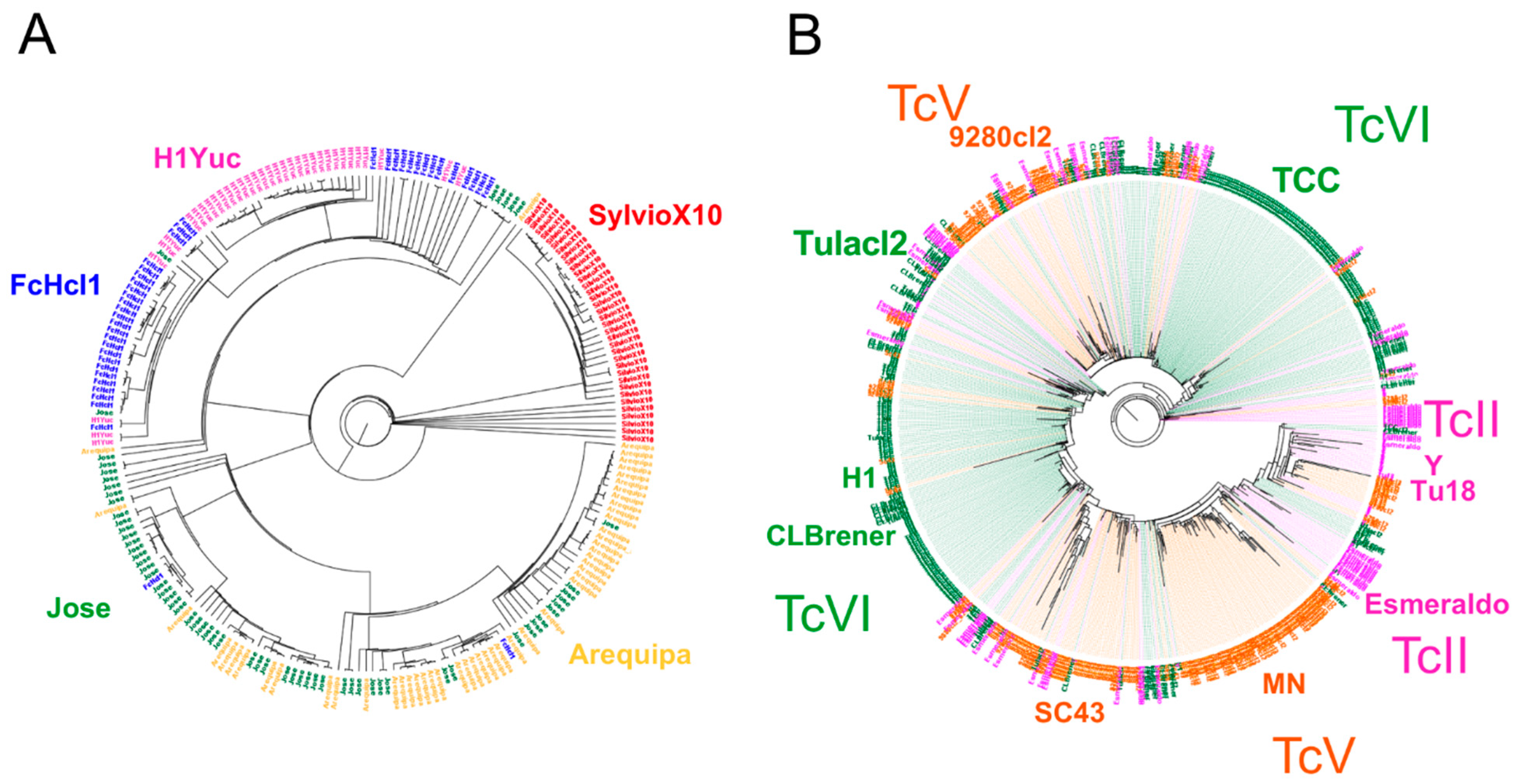
2.1. Whole Genome Comparison
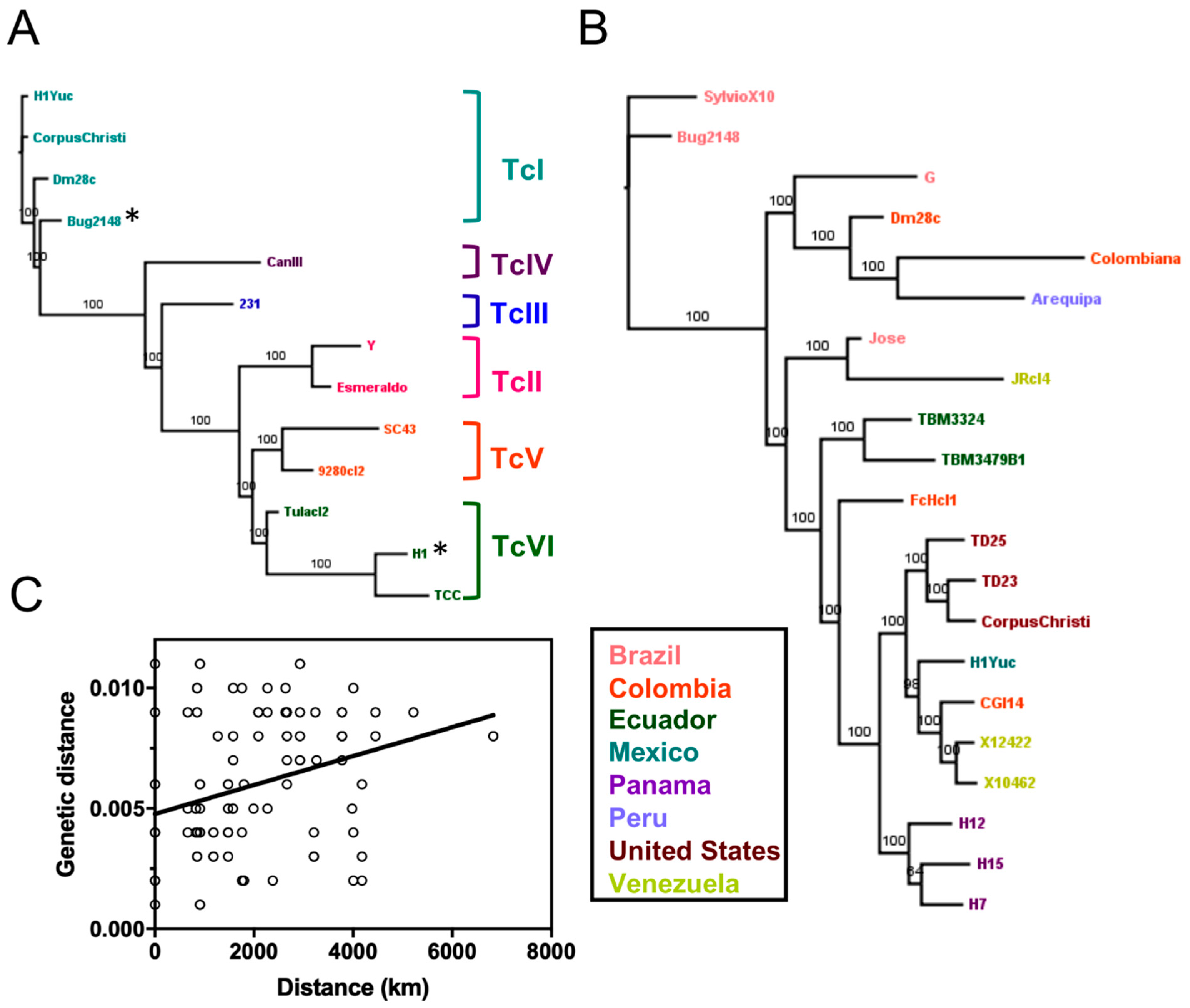
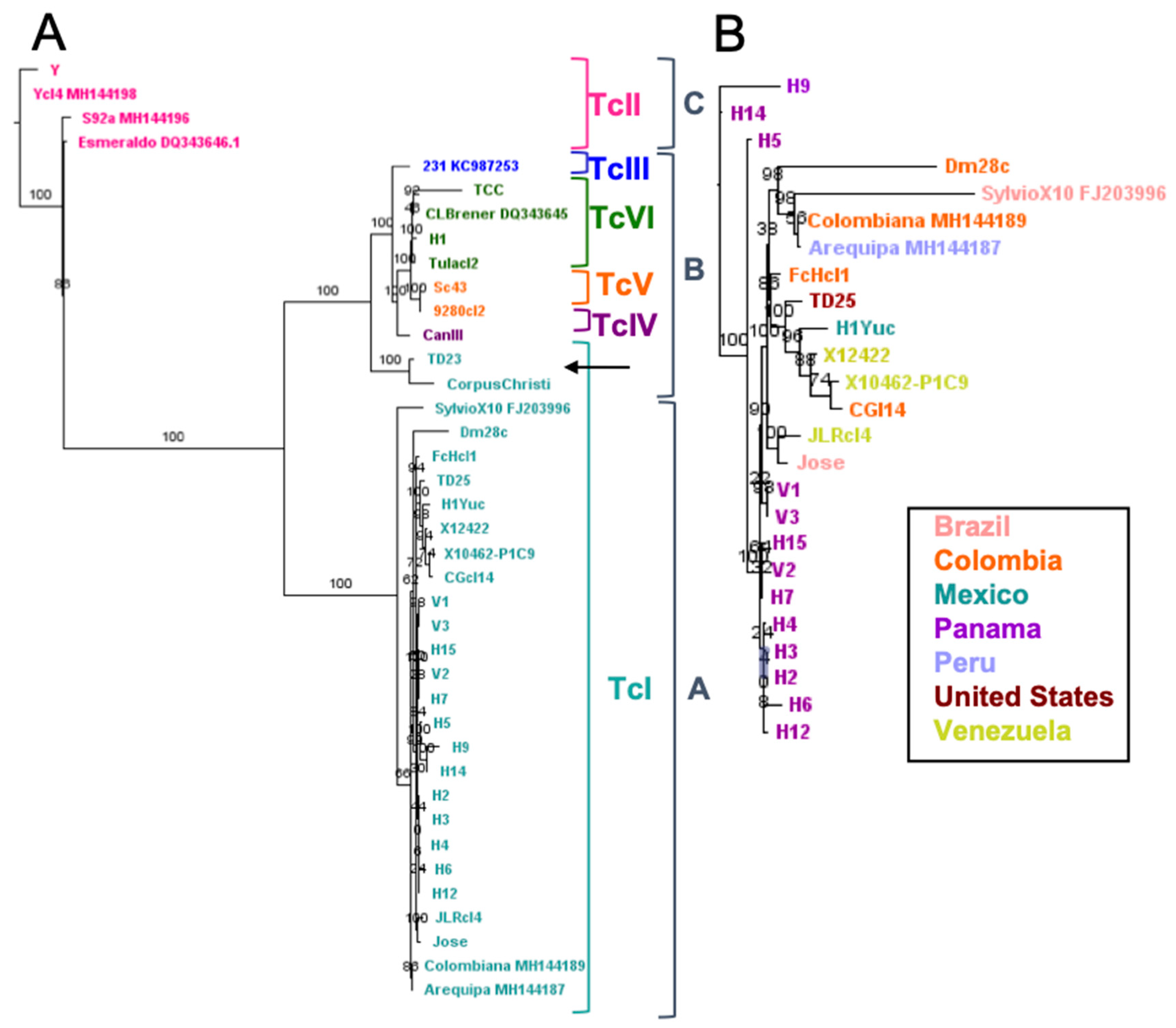
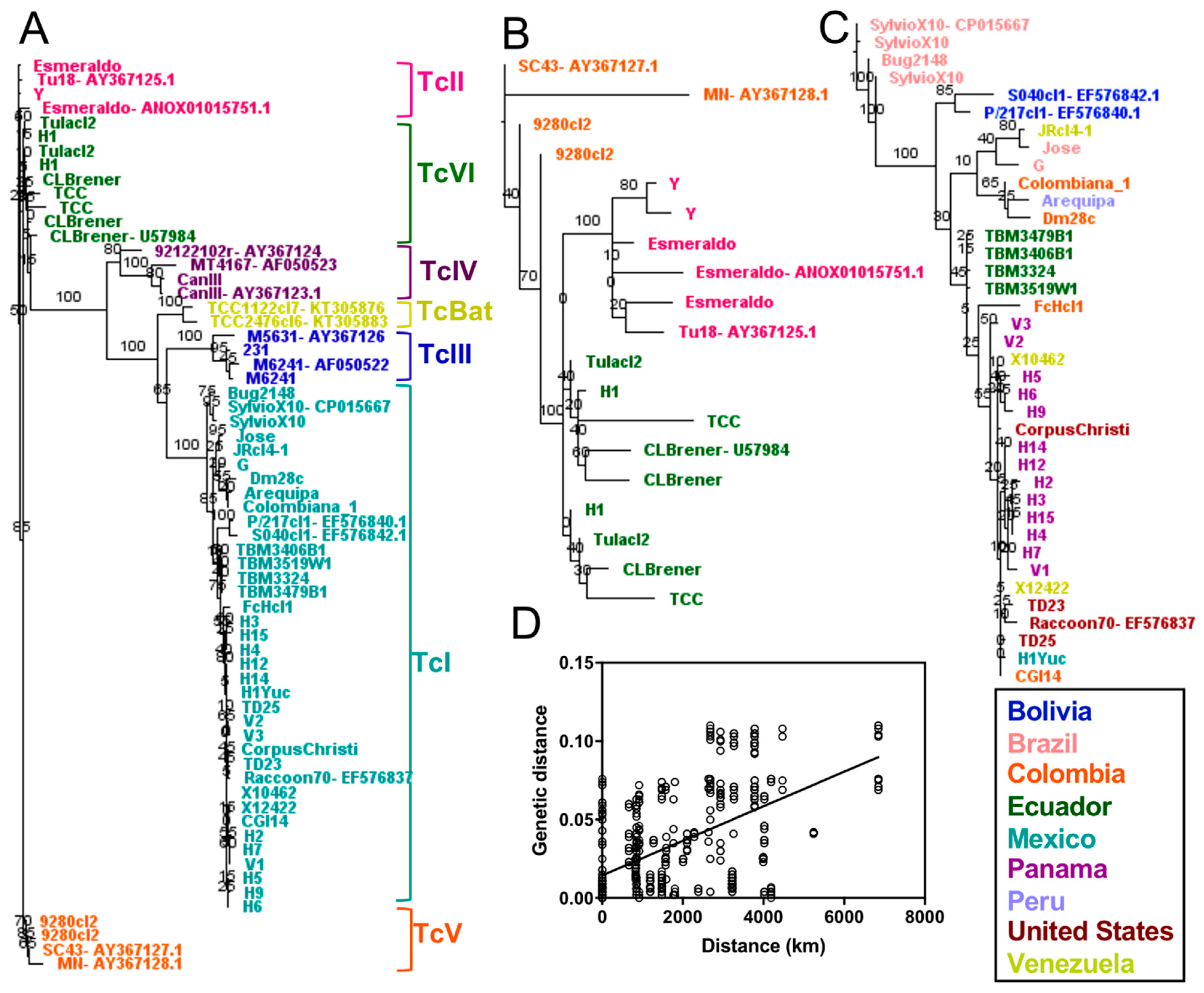
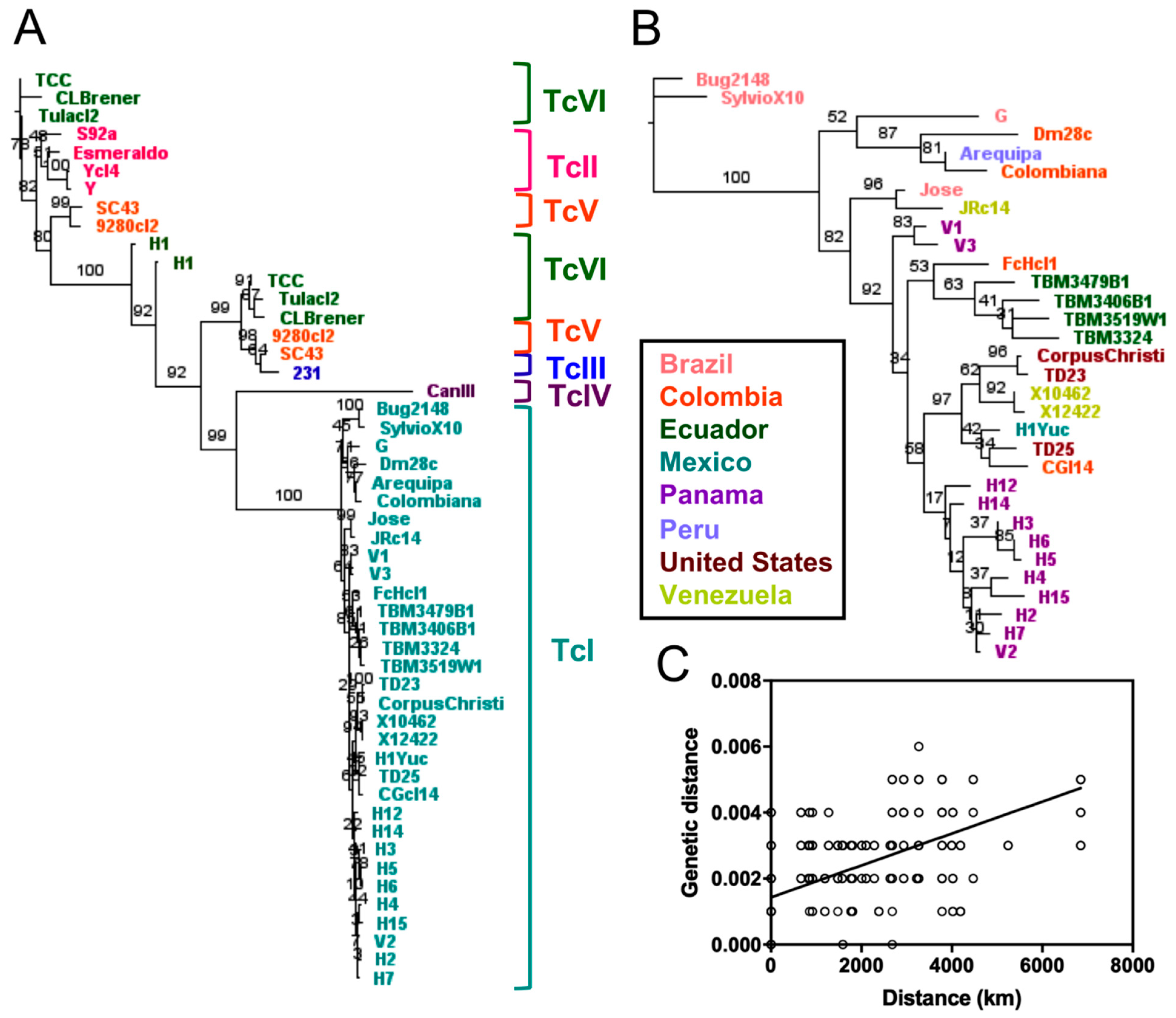
2.2. Phylogenetic Relationships Using Single and Multiple Markers
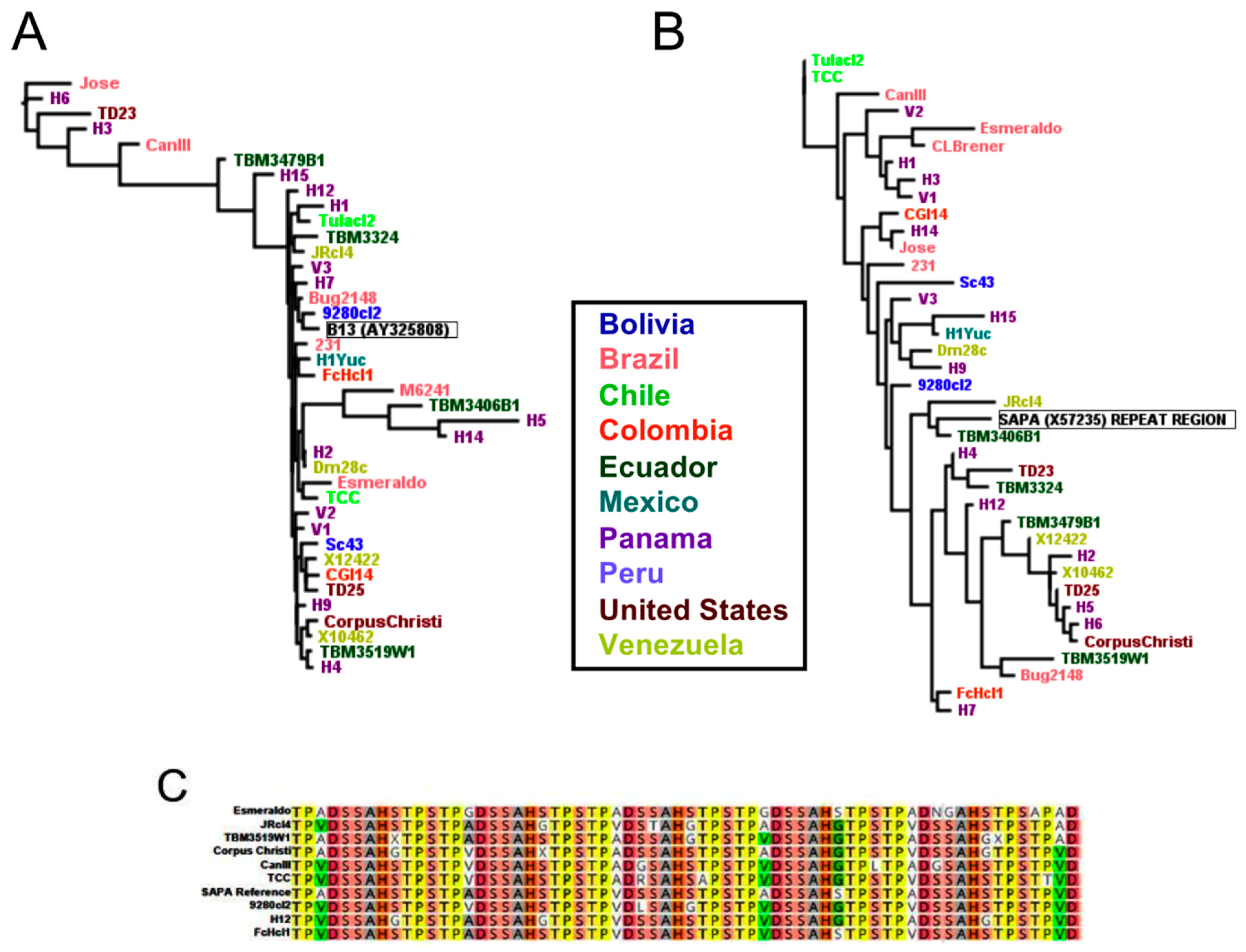
2.3. Diagnostic Antigen Conservation
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Trypanosoma cruzi Genome Sequences and Markers
5.2. Whole Genome Alignments
5.3. Single Marker Analysis
5.4. Concatenated Markers Analysis
5.5. Antigen Conservation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Browne, A.J.; Guerra, C.A.; Alves, R.V.; da Costa, V.M.; Wilson, A.L.; Pigott, D.M.; Hay, S.I.; Lindsay, S.W.; Golding, N.; Moyes, C.L. The contemporary distribution of Trypanosoma cruzi infection in humans, alternative hosts and vectors. Sci. Data 2017, 4, 170050. [Google Scholar] [CrossRef] [PubMed]
- Messenger, L.A.; Miles, M.A.; Bern, C. Between a bug and a hard place: Trypanosoma cruzi genetic diversity and the clinical outcomes of Chagas disease. Expert Rev. Anti. Infect. Ther. 2015, 13, 995–1029. [Google Scholar] [CrossRef] [PubMed]
- Vandermark, C.; Zieman, E.; Boyles, E.; Nielsen, C.K.; Davis, C.; Jiménez, F.A. Trypanosoma cruzi strain TcIV infects raccoons from Illinois. Memórias Inst. Oswaldo Cruz 2018, 113, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Fact Sheet: Chagas Disease [Internet]: PAHO. 2017. Available online: https://www.paho.org/en/documents/chagas-americas-public-health-workers-2017 (accessed on 18 December 2020).
- Lee, B.Y.; Bacon, K.M.; Bottazzi, M.E.; Hotez, P.J. Global economic burden of Chagas disease: A computational simulation model. Lancet Infect. Dis. 2013, 13, 342–348. [Google Scholar] [CrossRef]
- Messenger, L.A.; Llewellyn, M.S.; Bhattacharyya, T.; Franzén, O.; Lewis, M.D.; Ramírez, J.D.; Carrasco, H.J.; Andersson, B.; Miles, M.A. Multiple Mitochondrial Introgression Events and Heteroplasmy in Trypanosoma cruzi Revealed by Maxicircle MLST and Next Generation Sequencing. PLoS Negl. Trop. Dis. 2012, 6, e1584. [Google Scholar] [CrossRef] [PubMed]
- Zingales, B.; Miles, M.A.; Campbell, D.A.; Tibayrenc, M.; Macedo, A.M.; Teixeira, M.M.; Schijman, A.G.; Llewellyn, M.S.; Lages-Silva, E.; Machado, C.R.; et al. The revised Trypanosoma cruzi subspecific nomenclature: Rationale, epidemiological relevance and research applications. Infect. Genet. Evol. 2012, 12, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Marcili, A.; Lima, L.; Cavazzana, M.J.; Junqueira, A.C.V.; Veludo, H.H.; Da Silva, F.M.; Campaner, M.; Paiva, F.; Nunes, V.L.B.; Teixeira, M.M.G. A new genotype of Trypanosoma cruzi associated with bats evidenced by phylogenetic analyses using SSU rDNA, cytochrome b and Histone H2B genes and genotyping based on ITS1 rDNA. Parasitology 2009, 136, 641–655. [Google Scholar] [CrossRef]
- Brenière, S.F.; Waleckx, E.; Barnabé, C. Over Six Thousand Trypanosoma cruzi Strains Classified into Discrete Typing Units (DTUs): Attempt at an Inventory. PLoS Negl. Trop. Dis. 2016, 10, e0004792. [Google Scholar] [CrossRef]
- Ramírez, J.C.; Torres, C.; Curto, M.D.L.A.; Schijman, A.G. New insights into Trypanosoma cruzi evolution, genotyping and molecular diagnostics from satellite DNA sequence analysis. PLoS Negl. Trop. Dis. 2017, 11, e0006139. [Google Scholar] [CrossRef]
- Lewis, M.D.; Llewellyn, M.S.; Yeo, M.; Acosta, N.; Gaunt, M.W.; Miles, M.A. Recent, Independent and Anthropogenic Origins of Trypanosoma cruzi Hybrids. PLoS Negl. Trop. Dis. 2011, 5, e1363. [Google Scholar] [CrossRef]
- Barnabé, C.; Mobarec, H.I.; Jurado, M.R.; Cortez, J.A.; Brenière, S.F. Reconsideration of the seven discrete typing units within the species Trypanosoma cruzi a new proposal of three reliable mitochondrial clades. Infect. Genet. Evol. 2016, 39, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Burgos, J.M.; Altcheh, J.; Bisio, M.; Duffy, T.; Valadares, H.M.; Seidenstein, M.E.; Piccinali, R.; Freitas, J.M.; Levin, M.J.; Macchi, L.; et al. Direct molecular profiling of minicircle signatures and lineages of Trypanosoma cruzi bloodstream populations causing congenital Chagas disease. Int. J. Parasitol. 2007, 37, 1319–1327. [Google Scholar] [CrossRef]
- Herrera, C.; Bargues, M.D.; Fajardo, A.; Montilla, M.; Triana, O.; Vallejo, G.A.; Guhl, F. Identifying four Trypanosoma cruzi I isolate haplotypes from different geographic regions in Colombia. Infect. Genet. Evol. 2007, 7, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Falla, A.; Herrera, C.; Fajardo, A.; Montilla, M.; Vallejo, G.A.; Guhl, F. Haplotype identification within Trypanosoma cruzi I in Colombian isolates from several reservoirs, vectors and humans. Acta Trop. 2009, 110, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Cura, C.I.; Mejía-Jaramillo, A.M.; Duffy, T.; Burgos, J.M.; Rodriguero, M.; Cardinal, M.V.; Kjos, S.; Gurgel-Gonçalves, R.; Blanchet, D.; De Pablos, L.M.; et al. Trypanosoma cruzi I genotypes in different geographical regions and transmission cycles based on a microsatellite motif of the intergenic spacer of spliced-leader genes. Int. J. Parasitol. 2010, 40, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- Talavera-López, C.; Messenger, L.A.; Lewis, M.D.; Yeo, M.; Reis-Cunha, J.L.; Bartholomeu, D.C.; Calzada, J.E.; Saldaña, A.; Ramírez, J.D.; Guhl, F.; et al. Repeat-driven generation of antigenic diversity in a major human pathogen Trypanosoma cruzi. bioRxiv 2018, 283531. [Google Scholar]
- Reis-Cunha, J.L.; Baptista, R.P.; Rodrigues-Luiz, G.F.; Coqueiro-Dos-Santos, A.; Valdivia, H.O.; De Almeida, L.V.; Cardoso, M.S.; D’Ávila, D.A.; Dias, F.H.C.; Fujiwara, R.T.; et al. Whole genome sequencing of Trypanosoma cruzi field isolates reveals extensive genomic variability and complex aneuploidy patterns within TcII DTU. BMC Genom. 2018, 19, 816. [Google Scholar] [CrossRef]
- Barnabé, C.; Brisse, S.; Tibayrenc, M. Population structure and genetic typing of Trypanosoma cruzi, the agent of Chagas disease: A multilocus enzyme electrophoresis approach. Parasitology 2000, 120, 513–526. [Google Scholar] [CrossRef]
- Baptista, R.d.P.; D’Ávila, D.A.; Segatto, M.; do Valle, Í.F.; Franco, G.R.; Valadares, H.M.S.; Dias Gontijo, E.; da Cunha Galvão, L.M.; Junho Pena, S.D.; Chiari, E.; et al. Evidence of substantial recombination among Trypanosoma cruzi II strains from Minas Gerais. Infect. Genet. Evol. 2014, 22, 183–191. [Google Scholar] [CrossRef]
- Yeo, M.; Mauricio, I.L.; Messenger, L.A.; Lewis, M.D.; Llewellyn, M.S.; Acosta, N.; Bhattacharyya, T.; Diosque, P.; Carrasco, H.J.; Miles, M.A. Multilocus Sequence Typing (MLST) for Lineage Assignment and High Resolution Diversity Studies in Trypanosoma cruzi. PLoS Negl. Trop. Dis. 2011, 5, e1049. [Google Scholar] [CrossRef]
- Roellig, D.M.; Savage, M.Y.; Fujita, A.W.; Barnabé, C.; Tibayrenc, M.; Steurer, F.J.; Yabsley, M.J. Genetic Variation and Exchange in Trypanosoma cruzi Isolates from the United States. PLoS ONE 2013, 8, e56198. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, M.S.; Lewis, M.D.; Acosta, N.; Yeo, M.; Carrasco, H.J.; Segovia, M.; Vargas, J.; Torrico, F.; Miles, M.A.; Gaunt, M.W. Trypanosoma cruzi IIc: Phylogenetic and Phylogeographic Insights from Sequence and Microsatellite Analysis and Potential Impact on Emergent Chagas Disease. PLoS Negl. Trop. Dis. 2009, 3, e510. [Google Scholar] [CrossRef]
- Izeta-Alberdi, A.; Ibarra-Cerdeña, C.N.; Moo-Llanes, D.A.; Ramsey, J.M. Geographical, landscape and host associations of Trypanosoma cruzi DTUs and lineages. Parasites Vectors 2016, 9, 631. [Google Scholar] [CrossRef]
- Pronovost, H.; Peterson, A.C.; Chavez, B.G.; Blum, M.J.; Dumonteil, E.; Herrera, C.P. Deep sequencing reveals multiclonality and new discrete typing units of Trypanosoma cruzi in rodents from the southern United States. J. Microbiol. Immunol. Infect. 2020, 53, 622–633. [Google Scholar] [CrossRef] [PubMed]
- Monje-Rumi, M.; Floridia-Yapur, N.; Zago, M.; Ragone, P.; Brandán, C.P.; Nuñez, S.; Barrientos, N.; Tomasini, N.; Diosque, P. Potential association of Trypanosoma cruzi DTUs TcV and TcVI with the digestive form of Chagas disease. Infect. Genet. Evol. 2020, 84, 104329. [Google Scholar] [CrossRef] [PubMed]
- Miles, M.A.; Póvoa, M.M.; Prata, A.; Cedillos, R.A.; De Souza, A.A.; Macedo, V. Do radically dissimilar Trypanosoma cruziI strains (zymodemes) cause Venezuelan and Brazilian forms of Chagas’ Disease? Lancet 1981, 317, 1338–1340. [Google Scholar] [CrossRef]
- Gruber, A.; Zingales, B. Trypanosoma cruzi: Characterization of Two Recombinant Antigens with Potential Application in the Diagnosis of Chagas? Disease. Exp. Parasitol. 1993, 76, 1–12. [Google Scholar] [CrossRef]
- Carmona, S.J.; Nielsen, M.; Schafer-Nielsen, C.; Mucci, J.; Altcheh, J.; Balouz, V.; Tekiel, V.; Frasch, A.C.; Campetella, O.; Buscaglia, C.A.; et al. Towards High-throughput Immunomics for Infectious Diseases: Use of Next-generation Peptide Microarrays for Rapid Discovery and Mapping of Antigenic Determinants. Mol. Cell. Proteom. 2015, 14, 1871–1884. [Google Scholar] [CrossRef]
- Guzmán-Gómez, D.; López-Monteon, A.; Lagunes-Castro, M.D.L.S.; Álvarez-Martínez, C.; Hernández-Lutzon, M.J.; Dumonteil, E.; Ramos-Ligonio, A. Highly discordant serology against Trypanosoma cruzi in central Veracruz, Mexico: Role of the antigen used for diagnostic. Parasites Vectors 2015, 8, 466. [Google Scholar] [CrossRef]
- Buekens, P.; Althabe, F.; López, B.; Graiff, O.; Ramírez-Sierra, M.J.; Sosa-Estani, S.; Wesson, D.M.; Bustamante, N.; Alger, J.; Zúniga, C.; et al. Congenital Transmission of Trypanosoma cruzi in Argentina, Honduras, and Mexico: An Observational Prospective Study. Am. J. Trop. Med. Hyg. 2018, 98, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Elmayan, A.; Tu, W.; Duhon, B.; Marx, P.; Wolfson, W.; Balsamo, G.; Herrera, C.; Dumonteil, E. High prevalence of Trypanosoma cruzi infection in shelter dogs from southern Louisiana, USA. Parasites Vectors 2019, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Whitman, J.D.; Bulman, C.A.; Gunderson, E.L.; Irish, A.M.; Townsend, R.L.; Stramer, S.L.; Sakanari, J.A.; Bern, C. Chagas Disease Serological Test Performance in U.S. Blood Donor Specimens. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef]
- Martin, D.L.; Weatherly, D.B.; A Laucella, S.; A Cabinian, M.; Crim, M.T.; Sullivan, S.; Heiges, M.; Craven, S.H.; Rosenberg, C.S.; Collins, M.H.; et al. CD8+ T-Cell Responses to Trypanosoma cruzi Are Highly Focused on Strain-Variant trans-Sialidase Epitopes. PLOS Pathog. 2006, 2, e77. [Google Scholar] [CrossRef] [PubMed]
- Weatherly, D.B.; Peng, D.; Tarleton, R.L. Recombination-driven generation of the largest pathogen repository of antigen variants in the protozoan Trypanosoma cruzi. BMC Genom. 2016, 17, 1–17. [Google Scholar] [CrossRef]
- Haolla, F.A.; Claser, C.; De Alencar, B.C.; Tzelepis, F.; De Vasconcelos, J.R.; De Oliveira, G.; Silvério, J.C.; Machado, A.V.; Lannes-Vieira, J.; Bruna-Romero, O.; et al. Strain-specific protective immunity following vaccination against experimental Trypanosoma cruzi infection. Vaccine 2009, 27, 5644–5653. [Google Scholar] [CrossRef]
- Arnal, A.; Villanueva-Lizama, L.; Teh-Poot, C.; Herrera, C.; Dumonteil, E. Extent of polymorphism and selection pressure on the Trypanosoma cruzi vaccine candidate antigen Tc24. Evol. Appl. 2020, 13, 2663–2672. [Google Scholar] [CrossRef]
- Macedo, A.M.; Oliveira, R.P.; Pena, S.D. Chagas disease: Role of parasite genetic variation in pathogenesis. Expert Rev. Mol. Med. 2002, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Andrade, S.G.; Magalhães, J.B.; Pontes, A.L. Evaluation of chemotherapy with benznidazole and nifurtimox in mice infected with Trypanosoma cruzi strains of different types*. Bull. World Heal. Organ. 1985, 63, 721–726. [Google Scholar]
- Filardi, L.; Brener, Z. Susceptibility and natural resistance of Trypanosoma cruzi strains to drugs used clinically in Chagas disease. Trans. R. Soc. Trop. Med. Hyg. 1987, 81, 755–759. [Google Scholar] [CrossRef]
- Zingales, B. Trypanosoma cruzi genetic diversity: Something new for something known about Chagas disease manifestations, serodiagnosis and drug sensitivity. Acta Trop. 2018, 184, 38–52. [Google Scholar] [CrossRef]
- Villanueva-Lizama, L.; Teh-Poot, C.; Majeau, A.; Herrera, C.; Dumonteil, E. Molecular Genotyping of Trypanosoma cruzi by Next-Generation Sequencing of the Mini-Exon Gene Reveals Infections With Multiple Parasite Discrete Typing Units in Chagasic Patients From Yucatan, Mexico. J. Infect. Dis. 2019, 219, 1980–1988. [Google Scholar] [CrossRef]
- Callejas-Hernández, F.; Gìronés, N.; Fresno, M. Genome Sequence of Trypanosoma cruzi Strain Bug2148. Genome Announc. 2018, 6, e01497-17. [Google Scholar] [CrossRef]
- DeCuir, J.T.; Tu, W.; Dumonteil, E.; Herrera, C. Sequence of Trypanosoma cruzi reference strain SC43 nuclear genome and kinetoplast maxicircle confirms a strong genetic structure among closely related parasite discrete typing units. Genome 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- A Minning, T.; Weatherly, D.B.; Flibotte, S.; Tarleton, R.L. Widespread, focal copy number variations (CNV) and whole chromosome aneuploidies in Trypanosoma cruzi strains revealed by array comparative genomic hybridization. BMC Genom. 2011, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.T.; Lima, F.M.; Barros, R.M.; Cortez, D.R.; Santos, M.F.; Cordero, E.M.; Ruiz, J.C.; Goldenberg, S.; Teixeira, M.M.G.; Da Silveira, J.F. Genome Size, Karyotype Polymorphism and Chromosomal Evolution in Trypanosoma cruzi. PLoS ONE 2011, 6, e23042. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, N.M.; Myler, P.J.; Bartholomeu, D.C.; Nilsson, D.; Aggarwal, G.; Tran, A.-N.; Ghedin, E.; Worthey, E.A.; Delcher, A.L.; Blandin, G.; et al. The Genome Sequence of Trypanosoma cruzi, Etiologic Agent of Chagas Disease. Science 2005, 309, 409–415. [Google Scholar] [CrossRef]
- Berná, L.; Rodriguez, M.; Chiribao, M.L.; Parodi-Talice, A.; Pita, S.; Rijo, G.; Alvarez-Valin, F.; Robello, C. Expanding an expanded genome: Long-read sequencing of Trypanosoma cruzi. Microb. Genom. 2018, 4, e000177. [Google Scholar] [CrossRef]
- Franzen, O.; Ochaya, S.; Sherwood, E.; Lewis, M.D.; Llewellyn, M.S.; Miles, M.A.; Andersson, B. Shotgun sequencing analysis of Trypanosoma cruzi I Sylvio X10/1 and comparison with T. cruzi VI CL Brener. PLoS Negl. Trop. Dis. 2011, 5, 984. [Google Scholar] [CrossRef]
- Díaz-Viraqué, F.; Pita, S.; Greif, G.; Souza, R.D.C.M.D.; Iraola, G.; Robello, C. Nanopore Sequencing Significantly Improves Genome Assembly of the Protozoan Parasite Trypanosoma cruzi. Genome Biol. Evol. 2019, 11, 1952–1957. [Google Scholar] [CrossRef]
- Tibayrenc, M.; Ayala, F.J. The population genetics of Trypanosoma cruzi revisited in the light of the predominant clonal evolution model. Acta Trop. 2015, 151, 156–165. [Google Scholar] [CrossRef]
- Bartholomeu, D.C.; Buck, G.A.; Teixeira, S.M.R.; El-Sayed, N.M.A. Genetics of Trypanosoma cruzi: Nuclear genome. In American Trypanosomiasis Chagas Disease One Hundred Years of Research; Telleria, J., Tibayrenc, M., Eds.; Elsevir: London, UK, 2010; pp. 443–448. [Google Scholar]
- Berry, A.S.F.; Salazar-Sánchez, R.; Castillo-Neyra, R.; Borrini-Mayorí, K.; Chipana-Ramos, C.; Vargas-Maquera, M.; Ancca-Juarez, J.; Náquira-Velarde, C.; Levy, M.Z.; Brisson, D.; et al. Sexual reproduction in a natural Trypanosoma cruzi population. PLoS Negl. Trop. Dis. 2019, 13, e0007392. [Google Scholar] [CrossRef]
- Schwabl, P.; Imamura, H.; Broeck, F.V.D.; Costales, J.A.; Maiguashca-Sánchez, J.; Miles, M.A.; Andersson, B.; Grijalva, M.J.; Llewellyn, M.S. Meiotic sex in Chagas disease parasite Trypanosoma cruzi. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Cosentino, R.O.; Agüero, F. A Simple Strain Typing Assay for Trypanosoma cruzi: Discrimination of Major Evolutionary Lineages from a Single Amplification Product. PLoS Negl. Trop. Dis. 2012, 6, e1777. [Google Scholar] [CrossRef]
- Augusto-Pinto, L.; Teixeira, S.M.R.; Pena, S.D.J.; Machado, C.R. Single-nucleotide polymorphisms of the Trypanosoma cruzi MSH2 gene support the existence of three phylogenetic lineages presenting differences in mismatch-repair efficiency. Genet. 2003, 164, 117–126. [Google Scholar]
- De Freitas, J.M.; Augusto-Pinto, L.; Pimenta, J.R.; Bastos-Rodrigues, L.; Goncalves, V.F.; Teixeira, S.M.; Chiari, E.; Junqueira, A.C.V.; Fernandes, O.; Macedo, A.M.; et al. Ancestral genomes, sex, and the population structure of Trypanosoma cruzi. PLoS Pathog. 2006, 2, e24. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.D.; Ma, J.; Yeo, M.; Carrasco, H.J.; Llewellyn, M.S.; Miles, M.A. Genotyping of Trypanosoma cruzi: Systematic Selection of Assays Allowing Rapid and Accurate Discrimination of All Known Lineages. Am. J. Trop. Med. Hyg. 2009, 81, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Souto, R.P.; Fernandes, O.; Macedo, A.M.; Campbell, D.A.; Zingales, B. DNA markers define two major phylogenetic lineages of Trypanosoma cruzi. Mol Biochem Parasitol. 1996, 83, 141–152. [Google Scholar] [CrossRef]
- Fernandes, O.; Sturm, N.R.; Derré, R.; A Campbell, D. The mini-exon gene: A genetic marker for zymodeme III of Trypanosoma cruzi. Mol. Biochem. Parasitol. 1998, 95, 129–133. [Google Scholar] [CrossRef]
- Tomasini, N.; Lauthier, J.J.; Rumi, M.M.M.; Ragone, P.G.; D’Amato, A.A.A.; Brandan, C.P.; Cura, C.I.; Schijman, A.G.; Barnabé, C.; Tibayrenc, M.; et al. Interest and limitations of Spliced Leader Intergenic Region sequences for analyzing Trypanosoma cruzi I phylogenetic diversity in the Argentinean Chaco. Infect. Genet. Evol. 2011, 11, 300–307. [Google Scholar] [CrossRef] [PubMed]
- McCarthy-Burke, C.; Taylor, Z.A.; Buck, G.A. Characterization of the spliced leader genes and transcripts in Trypanosoma cruzi. Gene 1989, 82, 177–189. [Google Scholar] [CrossRef]
- Houghton, R.L.; Stevens, Y.Y.; Hjerrild, K.; Guderian, J.; Okamoto, M.; Kabir, M.; Reed, S.G.; Leiby, D.A.; Morrow, W.J.W.; Lorca, M.; et al. Lateral Flow Immunoassay for Diagnosis of Trypanosoma cruzi Infection with High Correlation to the Radioimmunoprecipitation Assay. Clin. Vaccine Immunol. 2009, 16, 515–520. [Google Scholar] [CrossRef]
- Ibáñez, C.F.; Affranchino, J.L.; Macina, R.A.; Reyes, M.B.; Leguizamon, S.; Camargo, M.E.; Åslund, L.; Pettersson, U.; Frasch, A.C. Multiple Trypanosoma cruzi antigens containing tandemly repeated amino acid sequence motifs. Mol. Biochem. Parasitol. 1988, 30, 27–33. [Google Scholar] [CrossRef]
- Thomas, M.C.; Longobardo, M.V.; Carmelo, E.; Marañón, C.; Planelles, L.; E Patarroyo, M.; Alonso, C.; López, M.C. Mapping of the antigenic determinants of the T. cruzi kinetoplastid membrane protein-11. Identification of a linear epitope specifically recognized by human Chagasic sera. Clin. Exp. Immunol. 2001, 123, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Ponce, C.; Ponce, E.; Vinelli, E.; Montoya, A.; De Aguilar, V.; Gonzalez, A.; Zingales, B.; Rangel-Aldao, R.; Levin, M.J.; Esfandiari, J.; et al. Validation of a Rapid and Reliable Test for Diagnosis of Chagas’ Disease by Detection of Trypanosoma cruzi-Specific Antibodies in Blood of Donors and Patients in Central America. J. Clin. Microbiol. 2005, 43, 5065–5068. [Google Scholar] [CrossRef]
- Umezawa, E.S.; Nascimento, M.S.; Kesper, N., Jr.; Coura, J.R.; Borges-Pereira, J.; Junqueira, A.C.; Camargo, M.E. Immunoblot assay using excreted-secreted antigens of Trypanosoma cruzi in serodiagnosis of congenital, acute, and chronic Chagas’ disease. J. Clin. Microbiol. 1996, 34, 2143–2147. [Google Scholar] [CrossRef] [PubMed]
- Miles, M.A.; De Souza, A.A.A.; Povoa, M.; Shaw, J.J.; Lainson, R.; Toye, P.J. Isozymic heterogeneity of Trypanosoma cruzi in the first autochthonous patients with Chagas’ disease in Amazonian Brazil. Nat. Cell Biol. 1978, 272, 819–821. [Google Scholar] [CrossRef]
- Tibayrenc, M.; Ward, P.; Moya, A.; Ayala, F.J. Natural populations of Trypanosoma cruzi, the agent of Chagas disease, have a complex multiclonal structure. Proc. Natl. Acad. Sci. USA 1986, 83, 115–119. [Google Scholar] [CrossRef]
- Tibayrenc, M.; Ayala, F. Trypanosoma cruzi populations: More clonal than sexual. Parasitol. Today 1987, 3, 189–190. [Google Scholar] [CrossRef][Green Version]
- Tibayrenc, M.; Kjellberg, F.; Ayala, F.J. A clonal theory of parasitic protozoa: The population structures of Entamoeba, Giardia, Leishmania, Naegleria, Plasmodium, Trichomonas, and Trypanosoma and their medical and taxonomical consequences. Proc. Natl. Acad. Sci. USA 1990, 87, 2414–2418. [Google Scholar] [CrossRef]
- Schimanski, B.; Nguyen, T.N.; Gunzl, A. Characterization of a Multisubunit Transcription Factor Complex Essential for Spliced-Leader RNA Gene Transcription in Trypanosoma brucei. Mol. Cell. Biol. 2005, 25, 7303–7313. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kramer, S.; Marnef, A.; Standart, N.; Carrington, M. Inhibition of mRNA maturation in trypanosomes causes the formation of novel foci at the nuclear periphery containing cytoplasmic regulators of mRNA fate. J. Cell Sci. 2012, 125, 2896–2909. [Google Scholar] [CrossRef]
- Bhattacharyya, T.; Brooks, J.; Yeo, M.; Carrasco, H.J.; Lewis, M.D.; Llewellyn, M.S.; Miles, M.A. Analysis of molecular diversity of the Trypanosoma cruzi trypomastigote small surface antigen reveals novel epitopes, evidence of positive selection and potential implications for lineage-specific serology. Int. J. Parasitol. 2010, 40, 921–928. [Google Scholar] [CrossRef]
- Oury, B.; Tarrieu, F.; Monte-Alegre, A.; Ouaissi, A. Trypanosoma cruzi: Sequence polymorphism of the gene encoding the Tc52 immunoregulatory-released factor in relation to the phylogenetic diversity of the species. Exp. Parasitol. 2005, 111, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Zingales, B.; Andrade, S.G.; Briones, M.R.S.; A Campbell, D.; Chiari, E.; Fernandes, O.; Guhl, F.; Lages-Silva, E.; Macedo, A.M.; Machado, C.R.; et al. A new consensus for Trypanosoma cruzi intraspecific nomenclature: Second revision meeting recommends TcI to TcVI. Memórias do Instituto Oswaldo Cruz 2009, 104, 1051–1054. [Google Scholar] [CrossRef] [PubMed]
- Callejas-Hernández, F.; Rastrojo, A.; Poveda, C.; Gironès, N.; Fresno, M. Genomic assemblies of newly sequenced Trypanosoma cruzi strains reveal new genomic expansion and greater complexity. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Baptista, R.P.; Reis-Cunha, J.L.; DeBarry, J.D.; Chiari, E.; Kissinger, J.C.; Bartholomeu, D.C.; Macedo, A.M. Assembly of highly repetitive genomes using short reads: The genome of discrete typing unit III Trypanosoma cruzi strain 231. Microb. Genom. 2018, 4, e000156. [Google Scholar] [CrossRef] [PubMed]
- Bradwell, K.R.; Koparde, V.N.; Matveyev, A.V.; Serrano, M.G.; Alves, J.M.P.; Parikh, H.; Huang, B.; Lee, V.; Espinosa-Alvarez, O.; A Ortiz, P.; et al. Genomic comparison of Trypanosoma conorhini and Trypanosoma rangeli to Trypanosoma cruzi strains of high and low virulence. BMC Genom. 2018, 19, 770. [Google Scholar] [CrossRef]
- Leinonen, R.; Sugawara, H.; Shumway, M.; on behalf of the International Nucleotide Sequence Database Collaboration. The Sequence Read Archive. Nucleic Acids Res. 2010, 39, D19–D21. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple Genome Alignment with Gene Gain, Loss and Rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; Von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Ostell, J.; Pruitt, K.D.; Karsch-Mizrachi, I. GenBank. Nucleic Acids Res. 2020, 48, D84–D86. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A. FigTree version 1.4. 0. 2012; (accessed on 9 July 2020). [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | DTU | Accession No. | Host of Origin | Country |
|---|---|---|---|---|
| JRcl4 | TcI | SRX290934, SRX180959, SRX180957, SRX031468, SRX031469 | Human | Venezuela |
| Jose | TcI | SRR2057803 | Human | Brazil |
| Colombiana | TcI | SRX871218 | Human | Colombia |
| Arequipa | TcI | SRR1793866 | Triatomine | Peru |
| Dm28c * | TcI | GCA_003177105.1 | Opossum | Colombia |
| TBM3519W1 | TcI | SRR3676270 | Triatomine | Ecuador |
| TBM3324 | TcI | SRR3676267 | Triatomine | Ecuador |
| TBM3406B1 | TcI | SRR3676268 | Triatomine | Ecuador |
| TBM3479B1 | TcI | SRR3676269 | Triatomine | Ecuador |
| FcHcl1 | TcI | SRR3676318 | Human | Colombia |
| V1 | TcI | SRR3676313 | Triatomine | Panama |
| V2 | TcI | SRR3676314 | Triatomine | Panama |
| V3 | TcI | SRR3676315 | Triatomine | Panama |
| H1Yuc | TcI | SRX1851500 | Human | Mexico |
| H1 | TcVI ** | SRR3676277 | Human | Panama |
| H2 | TcI | SRR3676278 | Human | Panama |
| H3 | TcI | SRR3676279 | Human | Panama |
| H4 | TcI | SRR3676280 | Human | Panama |
| H5 | TcI | SRR3676281 | Human | Panama |
| H6 | TcI | SRR3676282 | Human | Panama |
| H7 | TcI | SRR3676283 | Human | Panama |
| H9 | TcI | SRR3676285 | Human | Panama |
| H12 | TcI | SRR3676309 | Human | Panama |
| H14 | TcI | SRR3676310 | Human | Panama |
| H15 | TcI | SRR3676312 | Human | Panama |
| TD23 | TcI | SRR3676272 | Triatomine | USA |
| TD25 | TcI | SRR3676273 | Triatomine | USA |
| Corpus Christi | TcI | SRX1054555 | Human | USA |
| Bug2148 * | TcI ** | NMZN00000000 | Human | Brazil |
| X12422 | TcI | SRR3676275 | Human | Venezuela |
| X10462 | TcI | SRR3676274 | Human | Venezuela |
| CGl14 | TcI | SRX1851527 | Human | Colombia |
| SylvioX10 | TcI | ADWP00000000 | Human | Brazil |
| G * | TcI | MKKV00000000 | Opossum | Brazil |
| Esmeraldo | TcII | SRX022423, SRX022425, SRX022426, SRX022427, SRX022428, SRX022429, SRX271443, SRX271444, SRX2015243, SRX2015244 | Human | Brazil |
| S11 | TcII | SRX3453751 | Human | Brazil |
| S15 | TcII | SRX3453752 | Human | Brazil |
| S154a | TcII | SRX3453753 | Human | Brazil |
| S23b | TcII | SRX3453755 | Human | Brazil |
| S1162a | TcII | SRX3453754 | Human | Brazil |
| S92a | TcII | SRX3453757 | Human | Brazil |
| S44a | TcII | SRX3453756 | Human | Brazil |
| Y | TcII | SRR1796718, SRR1797819 | Human | Brazil |
| Ycl4 * | TcII | GCA_003594405.1 | Human | Brazil |
| 231 * | TcIII | OGCJ00000000 | Human | Brazil |
| CanIII | TcIV | SRR1996498, SRR1996501 | Human | Brazil |
| 9280 cl2 | TcV | SRR1996492, SRR1996493, SRR1996496, SRR1996497, SRR1996502 | Human | Bolivia |
| SC43 * | TcV | GCA_015455285.1 | Triatomine | Bolivia |
| CLBrener * | TcVI | GCA_000209065.1 | Triatomine (culture derivate) | Brazil |
| Tula cl2 | TcVI | SRX268890, SRX268891, SRX268892, SRX268893, SRX268894, SRX268895, SRX268896 | Human | Chile |
| TCC * | TcVI | GCA_003177095.1 | Human (culture derivate) | Chile |
| Antigen | Accession No. | Current Diagnostic |
|---|---|---|
| Ag1 [63] | M21330 | Chagatest recombinant (Wiener), T-detect (InBios) |
| Ag36 [63] | M21331 | Chagatest recombinant (Wiener), T-detect (InBios) |
| Ag30 [63,64] | n/a | Chagatest recombinant (Wiener), T-detect (InBios) |
| KMP-11 [63,65] | n/a | T-detect (InBios) |
| B13 [66] | AY325808 | Stat-Pak (Chembio) |
| SAPA [63,67] | X57235 | Chagatest recombinant (Wiener), T-detect (InBios) |
| TcH49 [66] | L09564 | Stat-Pak (Chembio) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majeau, A.; Murphy, L.; Herrera, C.; Dumonteil, E. Assessing Trypanosoma cruzi Parasite Diversity through Comparative Genomics: Implications for Disease Epidemiology and Diagnostics. Pathogens 2021, 10, 212. https://doi.org/10.3390/pathogens10020212
Majeau A, Murphy L, Herrera C, Dumonteil E. Assessing Trypanosoma cruzi Parasite Diversity through Comparative Genomics: Implications for Disease Epidemiology and Diagnostics. Pathogens. 2021; 10(2):212. https://doi.org/10.3390/pathogens10020212
Chicago/Turabian StyleMajeau, Alicia, Laura Murphy, Claudia Herrera, and Eric Dumonteil. 2021. "Assessing Trypanosoma cruzi Parasite Diversity through Comparative Genomics: Implications for Disease Epidemiology and Diagnostics" Pathogens 10, no. 2: 212. https://doi.org/10.3390/pathogens10020212
APA StyleMajeau, A., Murphy, L., Herrera, C., & Dumonteil, E. (2021). Assessing Trypanosoma cruzi Parasite Diversity through Comparative Genomics: Implications for Disease Epidemiology and Diagnostics. Pathogens, 10(2), 212. https://doi.org/10.3390/pathogens10020212