
Genomic Characteristics of Stx2e-Producing Escherichia coli Strains Derived from Humans, Animals, and Meats

 ,
, 
 ,
,
Abstract
1. Introduction
2. Results
2.1. Prevalence of stx2e-Carrying STEC Strains in Diverse Hosts
2.2. Human-Derived Stx2e-STEC Strains
2.3. Molecular Characteristics of Stx2e-STEC Strains from Different Sources
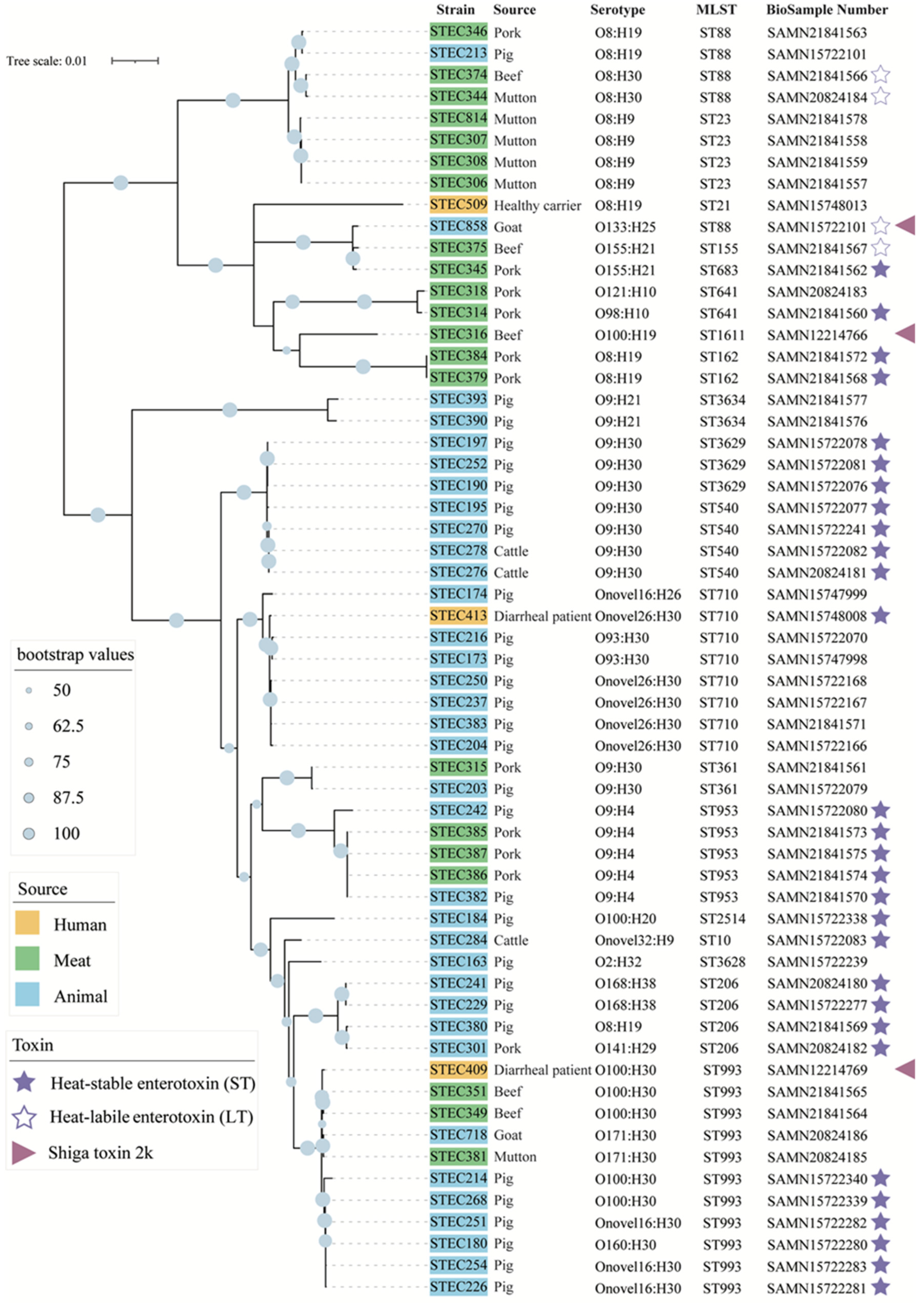
2.4. Phylogenetic Relationships of the Stx2e-STEC Strains from Different Hosts
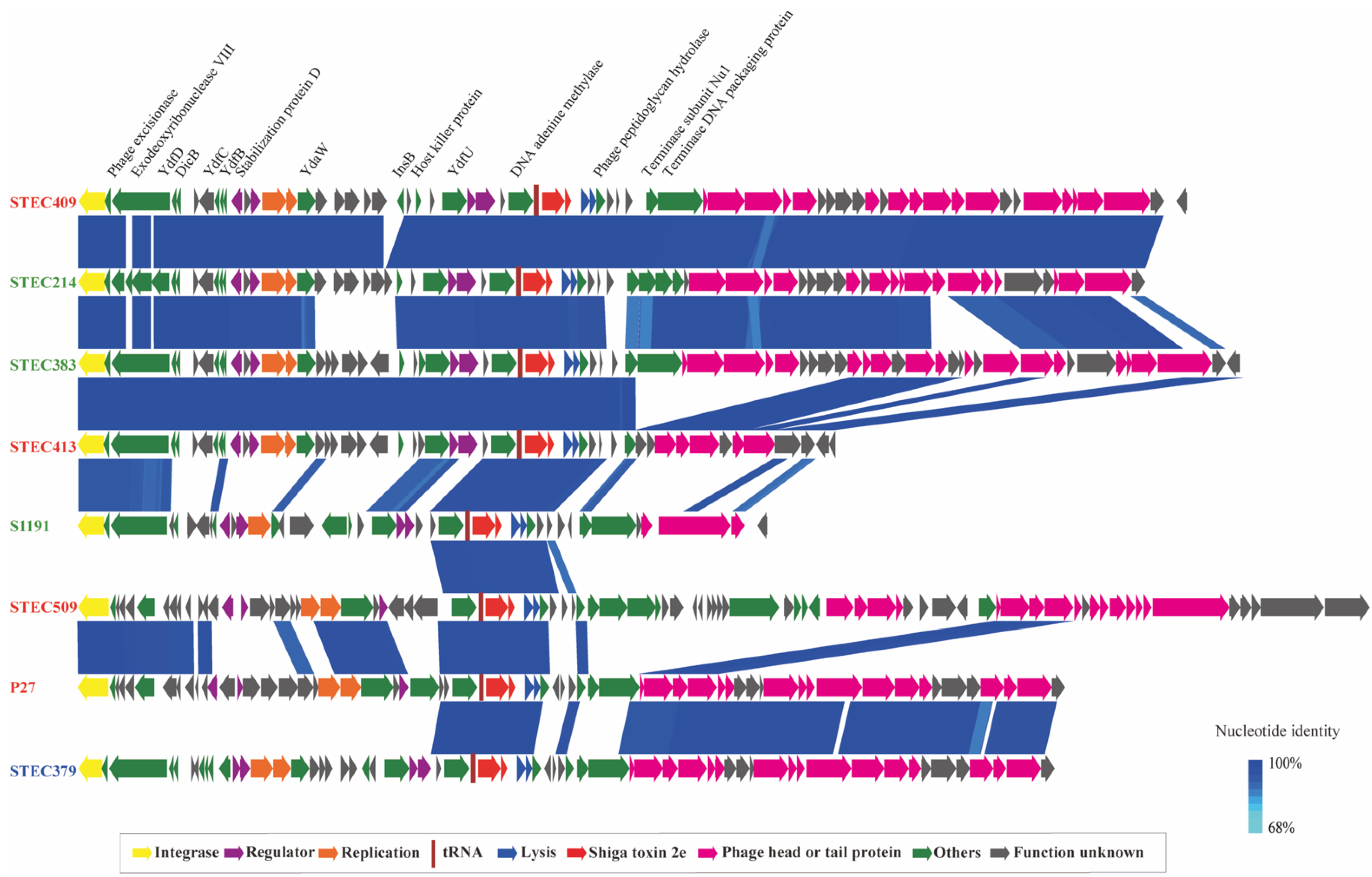
2.5. Genetic Feature of Stx2e-Converting Prophages
2.6. Antimicrobial Resistance of Stx2e-STEC Strains
3. Discussion
4. Materials and Methods
4.1. Stx2e-STEC Strains Collection
4.2. Whole-Genome Sequencing (WGS), Assembly, and Annotation
4.3. Determination of stx2 Subtype, Serotype, MLST, Virulence Factor Genes and Antimicrobial Resistance Genes
4.4. Single-Nucleotide Polymorphism (SNP)-Based Phylogeny
4.5. Genomic Characterization of Stx2e-Converting Prophages
4.6. Antimicrobial Resistance Testing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bryan, A.; Youngster, I.; McAdam, A.J. Shiga toxin producing Escherichia coli. Clin. Lab. Med. 2015, 35, 247–272. [Google Scholar] [CrossRef]
- Gould, L.H.; Mody, R.K.; Ong, K.L.; Clogher, P.; Cronquist, A.B.; Garman, K.N.; Lathrop, S.; Medus, C.; Spina, N.L.; Webb, T.H.; et al. Increased recognition of non-O157 Shiga toxin-producing Escherichia coli infections in the United States during 2000–2010: Epidemiologic features and comparison with E. coli O157 infections. Foodborne Pathog. Dis. 2013, 10, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Bruyand, M.; Mariani-Kurkdjian, P.; Gouali, M.; de Valk, H.; King, L.A.; Le Hello, S.; Bonacorsi, S.; Loirat, C. Hemolytic uremic syndrome due to Shiga toxin-producing Escherichia coli infection. Med. Mal. Infect. 2018, 48, 167–174. [Google Scholar] [CrossRef]
- Rodríguez-Rubio, L.; Haarmann, N.; Schwidder, M.; Muniesa, M.; Schmidt, H. Bacteriophages of Shiga toxin-producing Escherichia coli and their contribution to pathogenicity. Pathogens 2021, 10, 404. [Google Scholar] [CrossRef] [PubMed]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Pierard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Fu, S.; Zhang, J.; Fan, R.; Xu, Y.; Sun, H.; He, X.; Xu, J.; Xiong, Y. Identification and pathogenomic analysis of an Escherichia coli strain producing a novel Shiga toxin 2 subtype. Sci. Rep. 2018, 8, 6756. [Google Scholar] [CrossRef]
- Yang, X.; Bai, X.; Zhang, J.; Sun, H.; Fu, S.; Fan, R.; He, X.; Scheutz, F.; Matussek, A.; Xiong, Y. Escherichia coli strains producing a novel Shiga toxin 2 subtype circulate in China. Int. J. Med. Microbiol. 2020, 310, 151377. [Google Scholar] [CrossRef] [PubMed]
- Probert, W.S.; McQuaid, C.; Schrader, K. Isolation and identification of an Enterobacter cloacae strain producing a novel subtype of Shiga toxin type 1. J. Clin. Microbiol. 2014, 52, 2346–2351. [Google Scholar] [CrossRef]
- Lee, K.S.; Jeong, Y.J.; Lee, M.S. Escherichia coli Shiga toxins and gut microbiota interactions. Toxins 2021, 13, 416. [Google Scholar] [CrossRef]
- Bergan, J.; Dyve Lingelem, A.B.; Simm, R.; Skotland, T.; Sandvig, K. Shiga toxins. Toxicon 2012, 60, 1085–1107. [Google Scholar] [CrossRef]
- Pacheco, A.R.; Sperandio, V. Shiga toxin in enterohemorrhagic E. coli: Regulation and novel anti-virulence strategies. Front. Cell. Infect. Microbiol. 2012, 2, 81. [Google Scholar] [CrossRef] [PubMed]
- Steil, D.; Schepers, C.L.; Pohlentz, G.; Legros, N.; Runde, J.; Humpf, H.U.; Karch, H.; Muthing, J. Shiga toxin glycosphingolipid receptors of Vero-B4 kidney epithelial cells and their membrane microdomain lipid environment. J. Lipid Res. 2015, 56, 2322–2336. [Google Scholar] [CrossRef]
- Baldo, V.; Salogni, C.; Giovannini, S.; D’Incau, M.; Boniotti, M.B.; Birbes, L.; Pitozzi, A.; Formenti, N.; Grassi, A.; Pasquali, P.; et al. Pathogenicity of Shiga toxin type 2e Escherichia coli in pig colibacillosis. Front. Vet. Sci. 2020, 7, 545818. [Google Scholar] [CrossRef] [PubMed]
- Arancia, S.; Iurescia, M.; Lorenzetti, S.; Stravino, F.; Buccella, C.; Caprioli, A.; Franco, A.; Battisti, A.; Morabito, S.; Tozzoli, R. Detection and isolation of Shiga toxin-producing Escherichia coli (STEC) strains in caecal samples from pigs at slaughter in Italy. Vet. Med. Sci. 2019, 5, 462–469. [Google Scholar] [CrossRef]
- Vernozy-Rozand, C.; Montet, M.P.; Bertin, Y.; Trably, F.; Girardeau, J.P.; Martin, C.; Livrelli, V.; Beutin, L. Serotyping, Stx2 subtyping, and characterization of the locus of enterocyte effacement island of Shiga toxin-producing Escherichia coli and E. coli O157:H7 strains isolated from the environment in France. Appl. Environ. Microbiol. 2004, 70, 2556–2559. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fasel, D.; Mellmann, A.; Cernela, N.; Hachler, H.; Fruth, A.; Khanna, N.; Egli, A.; Beckmann, C.; Hirsch, H.H.; Goldenberger, D.; et al. Hemolytic uremic syndrome in a 65-Year-old male linked to a very unusual type of stx2e- and eae-harboring O51:H49 Shiga toxin-producing Escherichia coli. J. Clin. Microbiol. 2014, 52, 1301–1303. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, A.W.; Bielaszewska, M.; Zhang, W.L.; Pulz, M.; Kuczius, T.; Ammon, A.; Karch, H. Escherichia coli harboring Shiga toxin 2 gene variants: Frequency and association with clinical symptoms. J. Infect. Dis. 2002, 185, 74–84. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Ignacio, A.; Martins, F.H.; Rocha, L.B.; Piazza, R.M.F.; Vaz, T.M.I.; Avila-Campos, M.J.; Nakano, V. Presence of Shiga toxin 2e-producing Escherichia coli and atypical enteropathogenic E. coli in an asymptomatic child. JMM Case Rep. 2014, 1, e000001. [Google Scholar] [CrossRef]
- Saupe, A.; Edel, B.; Pfister, W.; Loffler, B.; Ehricht, R.; Rodel, J. Acute diarrhoea due to a Shiga toxin 2e-producing Escherichia coli O8:H19. JMM Case Rep. 2017, 4, e005099. [Google Scholar] [CrossRef]
- Meng, Q.; Bai, X.; Zhao, A.; Lan, R.; Du, H.; Wang, T.; Shi, C.; Yuan, X.; Bai, X.; Ji, S.; et al. Characterization of Shiga toxin-producing Escherichia coli isolated from healthy pigs in China. BMC Microbiol. 2014, 14, 5. [Google Scholar] [CrossRef]
- Sonntag, A.K.; Bielaszewska, M.; Mellmann, A.; Dierksen, N.; Schierack, P.; Wieler, L.H.; Schmidt, M.A.; Karch, H. Shiga toxin 2e-producing Escherichia coli isolates from humans and pigs differ in their virulence profiles and interactions with intestinal epithelial cells. Appl. Environ. Microbiol. 2005, 71, 8855–8863. [Google Scholar] [CrossRef]
- Muniesa, M.; Recktenwald, J.; Bielaszewska, M.; Karch, H.; Schmidt, H. Characterization of a shiga toxin 2e-converting bacteriophage from an Escherichia coli strain of human origin. Infect. Immun. 2000, 68, 4850–4855. [Google Scholar] [CrossRef]
- Hostetter, S.J.; Helgerson, A.F.; Paton, J.C.; Paton, A.W.; Cornick, N.A. Therapeutic use of a receptor mimic probiotic reduces intestinal Shiga toxin levels in a piglet model of hemolytic uremic syndrome. BMC Res. Notes 2014, 7, 331. [Google Scholar] [CrossRef]
- Pinto, G.; Sampaio, M.; Dias, O.; Almeida, C.; Azeredo, J.; Oliveira, H. Insights into the genome architecture and evolution of Shiga toxin encoding bacteriophages of Escherichia coli. BMC Genom. 2021, 22, 366. [Google Scholar] [CrossRef] [PubMed]
- Cha, W.; Fratamico, P.M.; Ruth, L.E.; Bowman, A.S.; Nolting, J.M.; Manning, S.D.; Funk, J.A. Prevalence and characteristics of Shiga toxin-producing Escherichia coli in finishing pigs: Implications on public health. Int. J. Food Microbiol. 2018, 264, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Maluta, R.P.; Fairbrother, J.M.; Stella, A.E.; Rigobelo, E.C.; Martinez, R.; de Avila, F.A. Potentially pathogenic Escherichia coli in healthy, pasture-raised sheep on farms and at the abattoir in Brazil. Vet. Microbiol. 2014, 169, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Osek, J.; Gallien, P.; Protz, D. Characterization of shiga toxin-producing Escherichia coli strains isolated from calves in Poland. Comp. Immunol. Microbiol. Infect. Dis. 2000, 23, 267–276. [Google Scholar] [CrossRef]
- Feng, P.C.; Reddy, S. Prevalences of Shiga toxin subtypes and selected other virulence factors among Shiga-toxigenic Escherichia coli strains isolated from fresh produce. Appl. Environ. Microbiol. 2013, 79, 6917–6923. [Google Scholar] [CrossRef]
- Serrano, N.S.; Zweifel, C.; Corti, S.; Stephan, R. Microbiological quality and presence of foodborne pathogens in raw milk cheeses and raw meat products marketed at farm level in Switzerland. Ital. J. Food Saf. 2018, 7, 7337. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Han, D.; Lee, H.T.; Wi, S.M.; Park, J.H.; Jo, J.W.; Cho, Y.J.; Hahn, T.W.; Lee, S.; Kang, B.; et al. Pathogenic and phylogenetic characteristics of non-O157 Shiga toxin-producing Escherichia coli isolates from retail meats in South Korea. J. Vet. Sci. 2018, 19, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Beutin, L. Characteristics of Shiga toxin-producing Escherichia coli from meat and milk products of different origins and association with food producing animals as main contamination sources. Int. J. Food Microbiol. 2011, 146, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Bibbal, D.; Um, M.M.; Diallo, A.A.; Kerouredan, M.; Dupouy, V.; Toutain, P.L.; Bousquet-Melou, A.; Oswald, E.; Brugere, H. Mixing of Shiga toxin-producing and enteropathogenic Escherichia coli in a wastewater treatment plant receiving city and slaughterhouse wastewater. Int. J. Hyg. Environ. Health 2018, 221, 355–363. [Google Scholar] [CrossRef]
- Bai, X.; Zhang, J.; Hua, Y.; Jernberg, C.; Xiong, Y.; French, N.; Lofgren, S.; Hedenstrom, I.; Ambikan, A.; Mernelius, S.; et al. Genomic Insights into clinical Shiga toxin-producing Escherichia coli Strains: A 15-year period survey in Jonkoping, Sweden. Front. Microbiol. 2021, 12, 627861. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, L.; Zhou, Y.; Miao, Z. Prevalence and characterization of virulence genes in Escherichia coli isolated from piglets suffering post-weaning diarrhoea in Shandong Province, China. Vet. Med. Sci. 2020, 6, 69–75. [Google Scholar] [CrossRef]
- Yang, G.Y.; Guo, L.; Su, J.H.; Zhu, Y.H.; Jiao, L.G.; Wang, J.F. Frequency of diarrheagenic virulence genes and characteristics in Escherichia coli Isolates from pigs with diarrhea in China. Microorganisms 2019, 7, 308. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Wang, H.; Xin, Y.; Wei, R.; Tang, X.; Zhao, A.; Sun, H.; Zhang, W.; Wang, Y.; Xu, Y.; et al. Prevalence and characteristics of Shiga toxin-producing Escherichia coli isolated from retail raw meats in China. Int. J. Food Microbiol. 2015, 200, 31–38. [Google Scholar] [CrossRef]
- Pierard, D.; Huyghens, L.; Lauwers, S.; Lior, H. Diarrhoea associated with Escherichia coli producing porcine oedema disease verotoxin. Lancet 1991, 338, 762. [Google Scholar] [CrossRef]
- Thomas, A.; Cheasty, T.; Chart, H.; Rowe, B. Isolation of Vero cytotoxin-producing Escherichia coli serotypes O9ab:H- and O101:H-carrying VT2 variant gene sequences from a patient with haemolytic uraemic syndrome. Eur. J. Clin. Microbiol. Infect. Dis. 1994, 13, 1074–1076. [Google Scholar] [CrossRef]
- Beutin, L.; Kruger, U.; Krause, G.; Miko, A.; Martin, A.; Strauch, E. Evaluation of major types of Shiga toxin 2E-producing Escherichia coli bacteria present in food, pigs, and the environment as potential pathogens for humans. Appl. Environ. Microbiol. 2008, 74, 4806–4816. [Google Scholar] [CrossRef]
- Franke, S.; Harmsen, D.; Caprioli, A.; Pierard, D.; Wieler, L.H.; Karch, H. Clonal relatedness of Shiga-like toxin-producing Escherichia coli O101 strains of human and porcine origin. J. Clin. Microbiol. 1995, 33, 3174–3178. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, M.; Friedrich, A.W.; Grundmann, H.; de Boer, R.F.; Croughs, P.D.; Islam, M.A.; Kluytmans-van den Bergh, M.F.; Kooistra-Smid, A.M.; Rossen, J.W. Molecular characterization and phylogeny of Shiga toxin-producing Escherichia coli isolates obtained from two Dutch regions using whole genome sequencing. Clin. Microbiol. Infect. 2016, 22, 642.e1–642.e9. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bai, X.; Zhang, J.; Ambikan, A.; Jernberg, C.; Ehricht, R.; Scheutz, F.; Xiong, Y.; Matussek, A. Molecular characterization and comparative genomics of clinical hybrid Shiga toxin-producing and enterotoxigenic Escherichia coli (STEC/ETEC) strains in Sweden. Sci. Rep. 2019, 9, 5619. [Google Scholar] [CrossRef]
- Kruger, A.; Lucchesi, P.M. Shiga toxins and stx phages: Highly diverse entities. Microbiology 2015, 161, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Steyert, S.R.; Sahl, J.W.; Fraser, C.M.; Teel, L.D.; Scheutz, F.; Rasko, D.A. Comparative genomics and stx phage characterization of LEE-negative Shiga toxin-producing Escherichia coli. Front. Cell. Infect. Microbiol. 2012, 2, 133. [Google Scholar] [CrossRef]
- Lewis, L.A.; Astatke, M.; Umekubo, P.T.; Alvi, S.; Saby, R.; Afrose, J. Soluble expression, purification and characterization of the full length IS2 Transposase. Mob. DNA 2011, 2, 14. [Google Scholar] [CrossRef]
- Peter-Getzlaff, S.; Polsfuss, S.; Poledica, M.; Hombach, M.; Giger, J.; Böttger, E.C.; Zbinden, R.; Bloemberg, G.V. Detection of AmpC beta-lactamase in Escherichia coli: Comparison of three phenotypic confirmation assays and genetic analysis. J. Clin. Microbiol. 2011, 49, 2924–2932. [Google Scholar] [CrossRef]
- Davis, M.A.; Besser, T.E.; Orfe, L.H.; Baker, K.N.; Lanier, A.S.; Broschat, S.L.; New, D.; Call, D.R. Genotypic-phenotypic discrepancies between antibiotic resistance characteristics of Escherichia coli isolates from calves in management settings with high and low antibiotic use. Appl. Environ. Microbiol. 2011, 77, 3293–3299. [Google Scholar] [CrossRef]
- Cointe, A.; Birgy, A.; Bridier-Nahmias, A.; Mariani-Kurkdjian, P.; Walewski, V.; Levy, C.; Cohen, R.; Fach, P.; Delannoy, S.; Bidet, P.; et al. Escherichia coli O80 hybrid pathotype strains producing Shiga toxin and ESBL: Molecular characterization and potential therapeutic options. J. Antimicrob. Chemother. 2020, 75, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Fan, R.; Shao, K.; Yang, X.; Bai, X.; Fu, S.; Sun, H.; Xu, Y.; Wang, H.; Li, Q.; Hu, B.; et al. High prevalence of non-O157 Shiga toxin-producing Escherichia coli in beef cattle detected by combining four selective agars. BMC Microbiol. 2019, 19, 213. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Souvorov, A.; Agarwala, R.; Lipman, D.J. SKESA: Strategic k-mer extension for scrupulous assemblies. Genome Biol. 2018, 19, 153. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A Cross-Platform and Ultrafast Toolkit for FASTA/Q File Manipulation. PLoS ONE 2016, 11, e0163962. [Google Scholar] [CrossRef]
- Chin, C.S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Ingle, D.J.; Valcanis, M.; Kuzevski, A.; Tauschek, M.; Inouye, M.; Stinear, T.; Levine, M.M.; Robins-Browne, R.M.; Holt, K.E. In silico serotyping of E. coli from short read data identifies limited novel O-loci but extensive diversity of O:H serotype combinations within and between pathogenic lineages. Microb. Genom. 2016, 2, e000064. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, J.; Wang, X.; Bai, X.; Ma, J.; Dang, R.; Xiong, Y.; Fanning, S.; Bai, L.; Yang, Z. Characterization of five Escherichia coli isolates co-expressing ESBL and MCR-1 resistance mechanisms from different origins in China. Front. Microbiol. 2019, 10, 1994. [Google Scholar] [CrossRef]
- Jobling, M.G. Novel type II heat-labile enterotoxin produced by human disease outbreak isolates of Escherichia coli Is a member of the LT-IIa Family and should not form a new group: Clarification of nomenclature and precedent. Open Forum Infect. Dis. 2020, 7, ofaa408. [Google Scholar] [CrossRef]
- Ishimaru, K.; Sasaki, M.; Narimatsu, H.; Arimizu, Y.; Gotoh, Y.; Nakamura, K.; Hayashi, T.; Ogura, Y. Escherichia coli O8:H8 carrying a novel variant of the heat-labile enterotoxin LT2 gene caused outbreaks of diarrhea. Open Forum Infect. Dis. 2020, 7, ofaa021. [Google Scholar] [CrossRef]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid phylogenetic analysis of large samples of recombinant bacterial whole genome sequences using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Hu, B.; Bai, X.; Yang, X.; Cao, L.; Liu, Q.; Sun, H.; Li, J.; Zhang, J.; Jin, D.; et al. Antimicrobial resistance of non-O157 Shiga toxin-producing Escherichia coli isolated from humans and domestic animals. Antibiotics 2021, 10, 74. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Source | No. of STECs | No. of Stx2e-STECs (%) | No. of Stx2e-STECs Used for WGS a | |
|---|---|---|---|---|
| Human | Healthy carrier | 5 | 1 (20.0%) | 1 |
| Diarrheal patient | 39 | 2 (5.1%) | 2 | |
| Animal | Pig | 147 | 144 (98.0%) | 29 |
| Cattle | 172 | 5 (2.9%) | 3 | |
| Goat | 202 | 2 (1.0%) | 2 | |
| Chicken | 4 | 0 | 0 | |
| Tibetan antelope | 5 | 0 | 0 | |
| Yak | 126 | 0 | 0 | |
| Marmot | 9 | 0 | 0 | |
| Pika | 22 | 0 | 0 | |
| Meat | Pork | 12 | 11 (91.7%) | 11 |
| Mutton | 43 | 6 (14.0%) | 6 | |
| Beef | 27 | 5 (18.5%) | 5 | |
| Chicken | 1 | 0 | 0 | |
| Duck | 1 | 0 | 0 | |
| Environment | Water | 3 | 0 | 0 |
| Total | 818 | 176 (21.5%) | 59 |
| Strain | Total Length (bp) | G + C a Ratio (%) | No. CDS b | No. rRNA | No. tRNA | No. Prophages | No. Plasmids |
|---|---|---|---|---|---|---|---|
| STEC409 | 4,857,389 | 51.82 | 4753 | 22 | 94 | 20 | 2 |
| STEC413 | 4,948,664 | 50.61 | 4826 | 22 | 90 | 17 | 1 |
| STEC509 | 4,871,415 | 50.71 | 5003 | 22 | 87 | 8 | 0 |
| Antimicrobial Classes | AMR Phenotype | AMR Genes | No. of Phenotypic Resistant Isolates | No. of Phenotypic Susceptible Isolates | ||
|---|---|---|---|---|---|---|
| Resistant by Genotype | Susceptible by Genotype | Resistant by Genotype | Susceptible by Genotype | |||
| Tetracyclines | Tetracycline | tet(A), tet(D) | 6 | 38 | 0 | 15 |
| Quinolones | Nalidixic_Acid | oqxA, oqxB, qnrD1, qnrS1 | 13 | 22 | 6 | 18 |
| Trimethoprim | Trimethoprim-Sulfamethoxazole | dfrA12, dfrA14, dfrA15, dfrA17 | 10 | 1 | 19 | 29 |
| Phenicols | Chloramphenicol | cmlA1_1, cmlA6, floR | 10 | 2 | 0 | 47 |
| Colistin | Colistin | mcr-1, mcr-3.1 | 8 | 2 | 1 | 48 |
| Macrolides | Azithromycin | mphA, mrx | 4 | 4 | 0 | 51 |
| Nitrofurans | Nitrofurantoin | -b | 0 | 1 | 0 | 49 |
| β-lactamase | Ampicillin | ampC, blaACC-1c, blaCTX-M-14, blaCTX-M-65, blaTEM-116, blaTEM-1B | 12 | 0 | 41 | 6 |
| Cefotaxime | ||||||
| Aminoglycoside | Amikacin | aac(3)-IIa, aac(3)-IIc, aadA, aadA2, aadA5, aph(3′)-Ia, aph(3″)-Ib, aph(3′)-IIa, aph(6)-Id | 0 | 0 | 32 | 27 |
| Fosfomycins | - a | fosA3 | 0 | 0 | 1 | 58 |
| Sulfonamides | - a | sul1, sul2, sul3 | 0 | 0 | 31 | 28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Wu, Y.; Liu, Q.; Sun, H.; Luo, M.; Xiong, Y.; Matussek, A.; Hu, B.; Bai, X. Genomic Characteristics of Stx2e-Producing Escherichia coli Strains Derived from Humans, Animals, and Meats. Pathogens 2021, 10, 1551. https://doi.org/10.3390/pathogens10121551
Yang X, Wu Y, Liu Q, Sun H, Luo M, Xiong Y, Matussek A, Hu B, Bai X. Genomic Characteristics of Stx2e-Producing Escherichia coli Strains Derived from Humans, Animals, and Meats. Pathogens. 2021; 10(12):1551. https://doi.org/10.3390/pathogens10121551
Chicago/Turabian StyleYang, Xi, Yannong Wu, Qian Liu, Hui Sun, Ming Luo, Yanwen Xiong, Andreas Matussek, Bin Hu, and Xiangning Bai. 2021. "Genomic Characteristics of Stx2e-Producing Escherichia coli Strains Derived from Humans, Animals, and Meats" Pathogens 10, no. 12: 1551. https://doi.org/10.3390/pathogens10121551
APA StyleYang, X., Wu, Y., Liu, Q., Sun, H., Luo, M., Xiong, Y., Matussek, A., Hu, B., & Bai, X. (2021). Genomic Characteristics of Stx2e-Producing Escherichia coli Strains Derived from Humans, Animals, and Meats. Pathogens, 10(12), 1551. https://doi.org/10.3390/pathogens10121551