Inactivation of Prions by Low-Temperature Sterilization Technology Using Vaporized Gas Derived from a Hydrogen Peroxide–Peracetic Acid Mixture

Abstract
:1. Introduction
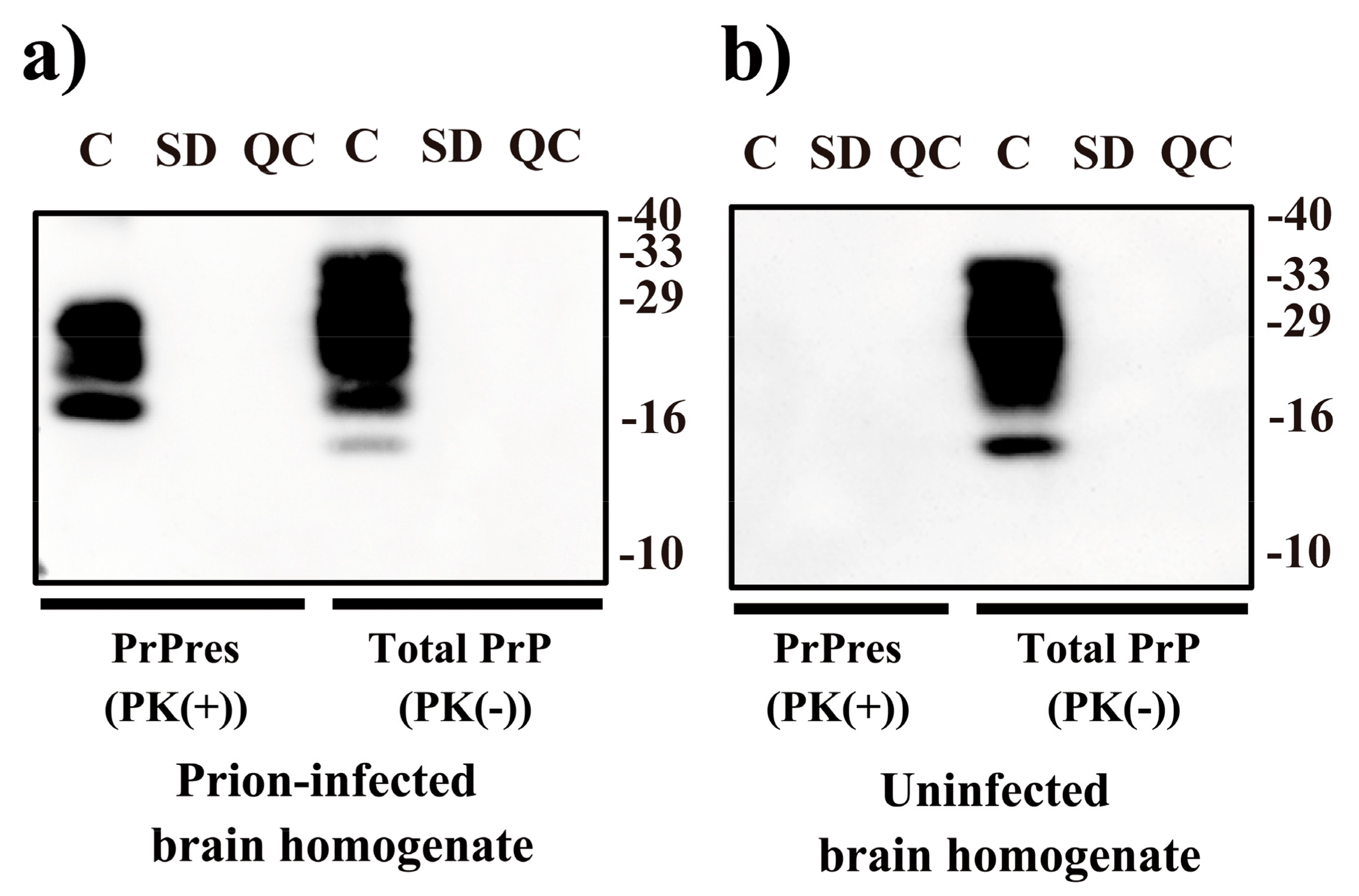
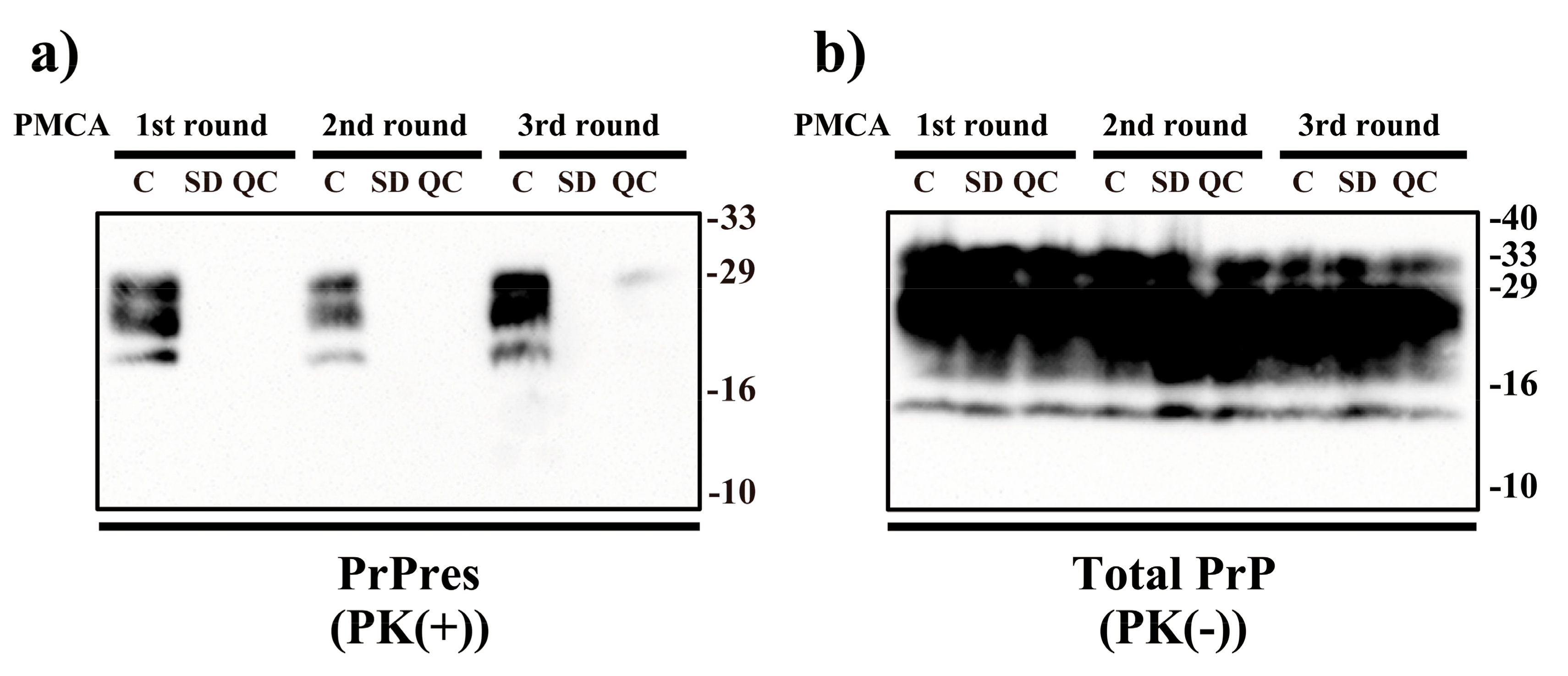
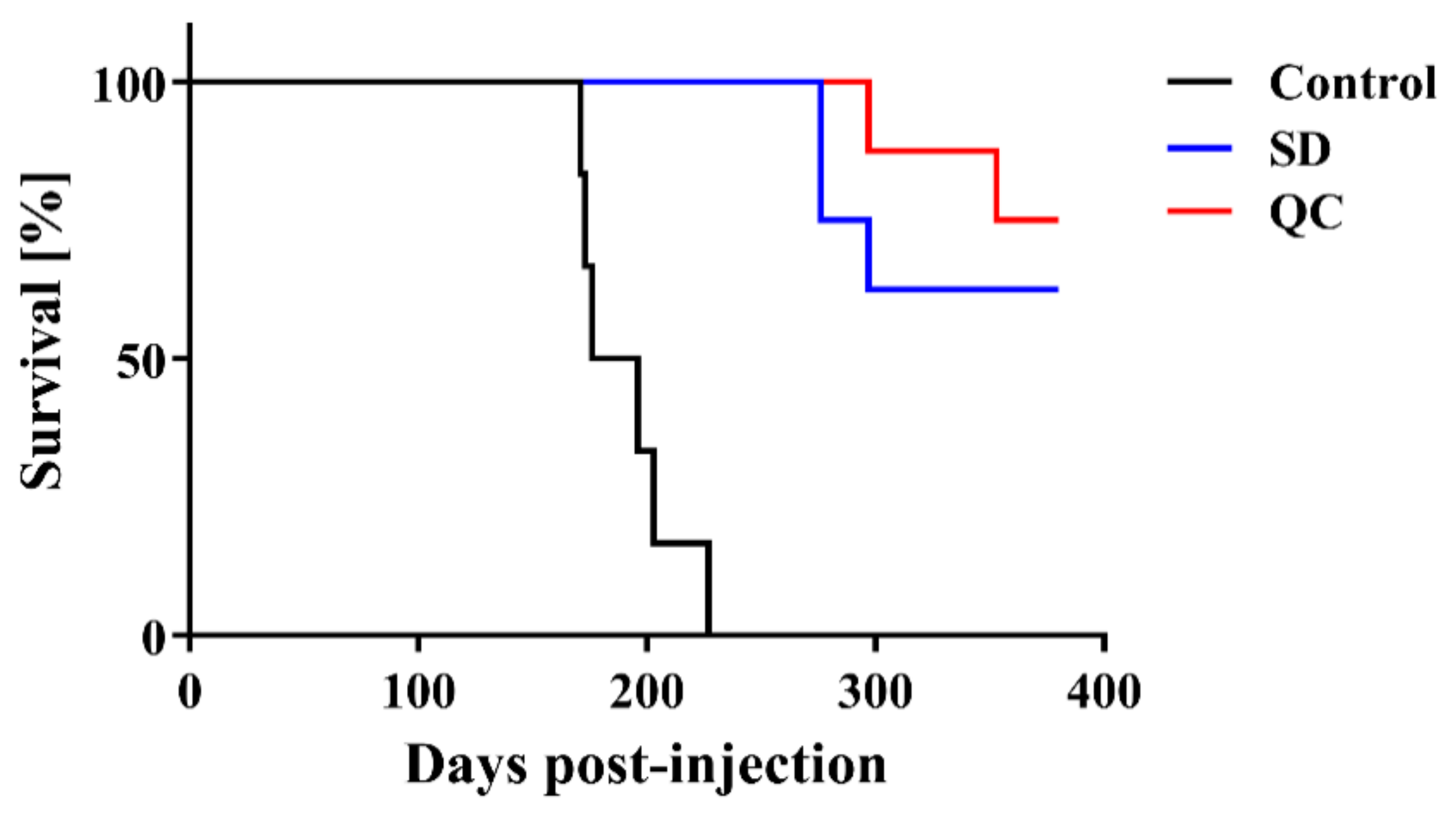
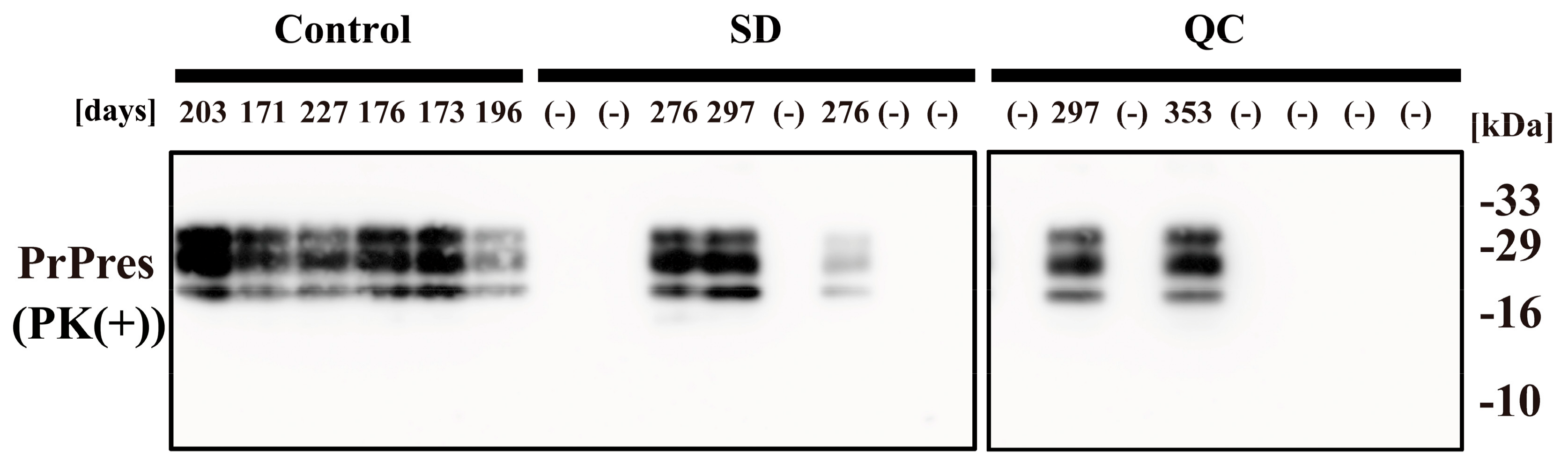
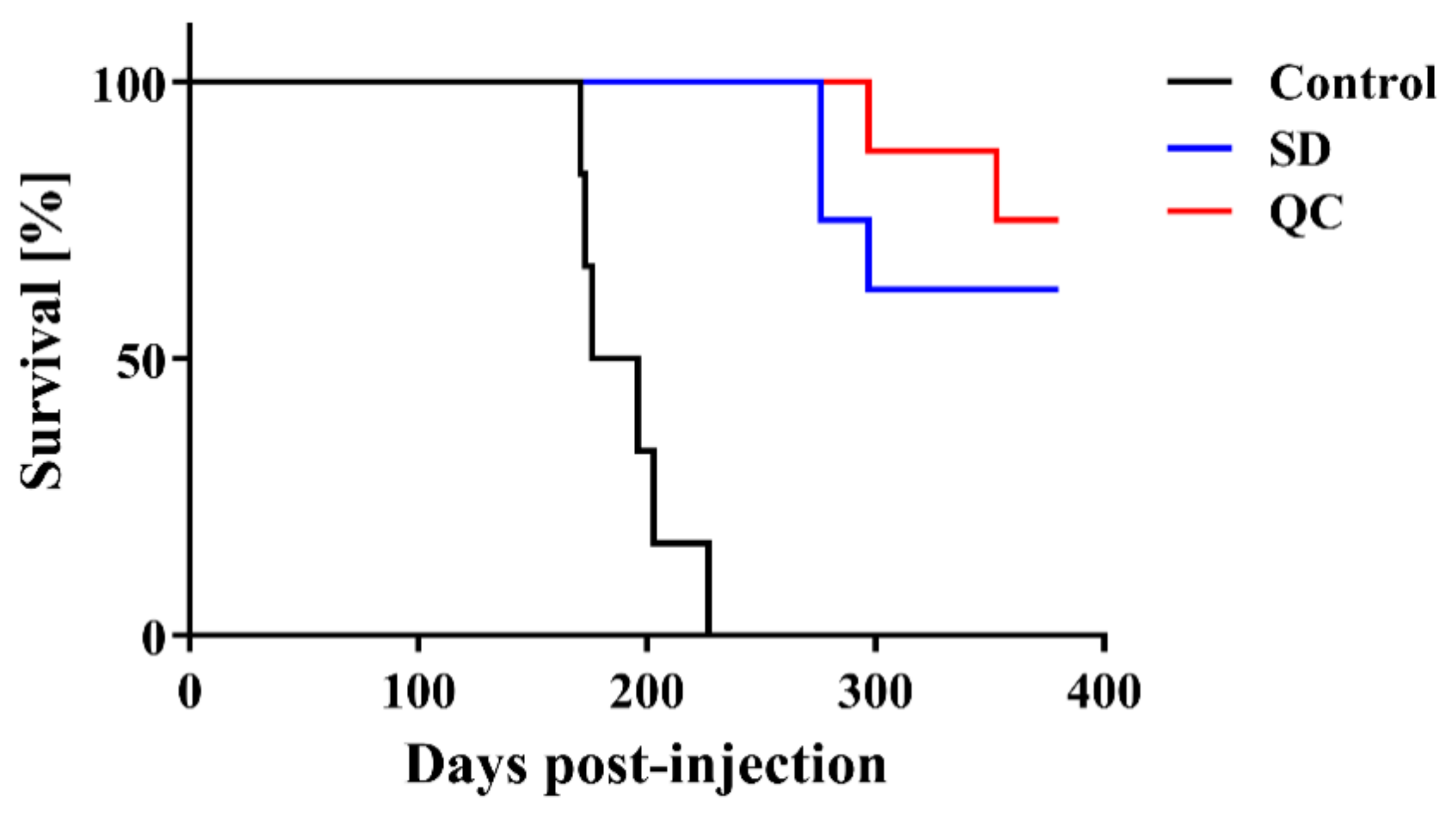
2. Results
3. Discussion
4. Materials and Methods
4.1. VHPPA Treatment of Prions
4.2. Prion Injection
4.3. Immunoblot Analysis
4.4. PMCA
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prusiner, S.B. Prions. Proc. Natl. Acad. Sci. USA 1998, 95, 13363–13383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prusiner, S.B. Molecular biology of prion diseases. Science 1991, 252, 1515–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakudo, A.; Ikuta, K. Prion protein functions and dysfunction in prion diseases. Curr. Med. Chem. 2009, 16, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Fiorini, M.; Bongianni, M.; Monaco, S.; Zanusso, G. Biochemical Characterization of Prions. Prog. Mol. Biol. Transl. Sci. 2017, 150, 389–407. [Google Scholar] [CrossRef] [PubMed]
- Baral, P.K.; Yin, J.; Aguzzi, A.; James, M.N.G. Transition of the prion protein from a structured cellular form (PrPC)) to the infectious scrapie agent (PrPSc). Protein. Sci. 2019, 28, 2055–2063. [Google Scholar] [CrossRef]
- Pan, K.M.; Baldwin, M.; Nguyen, J.; Gasset, M.; Serban, A.; Groth, D.; Mehlhorn, I.; Huang, Z.; Fletterick, R.J.; Cohen, F.E.; et al. Conversion of alpha-helices into beta-sheets features in the formation of the scrapie prion proteins. Proc. Natl. Acad. Sci. USA 1993, 90, 10962–10966. [Google Scholar] [CrossRef] [Green Version]
- Cohen, F.E.; Prusiner, S.B. Pathologic conformations of prion proteins. Annu. Rev. Biochem. 1998, 67, 793–819. [Google Scholar] [CrossRef]
- Onodera, T.; Sakudo, A. Introduction to current progress in advanced research on prions. Curr. Issues Mol. Biol. 2020, 36, 63–66. [Google Scholar] [CrossRef] [Green Version]
- Castilla, J.; Saa, P.; Morales, R.; Abid, K.; Maundrell, K.; Soto, C. Protein misfolding cyclic amplification for diagnosis and prion propagation studies. Methods Enzym. 2006, 412, 3–21. [Google Scholar] [CrossRef]
- Soto, C.; Saborio, G.P.; Anderes, L. Cyclic amplification of protein misfolding: Application to prion-related disorders and beyond. Trends Neurosci. 2002, 25, 390–394. [Google Scholar] [CrossRef]
- Giaccone, G.; Moda, F. PMCA Applications for prion detection in peripheral tissues of patients with variant Creutzfeldt-Jakob disease. Biomolecules 2020, 10, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakudo, A. Inactivation methods for prions. Curr. Issues Mol. Biol. 2020, 36, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Fichet, G.; Comoy, E.; Duval, C.; Antloga, K.; Dehen, C.; Charbonnier, A.; McDonnell, G.; Brown, P.; Lasmezas, C.I.; Deslys, J.P. Novel methods for disinfection of prion-contaminated medical devices. Lancet 2004, 364, 521–526. [Google Scholar] [CrossRef]
- Sakudo, A.; Ano, Y.; Onodera, T.; Nitta, K.; Shintani, H.; Ikuta, K.; Tanaka, Y. Fundamentals of prions and their inactivation (review). Int. J. Mol. Med. 2011, 27, 483–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutala, W.A.; Weber, D.J. Society for healthcare epidemiology of America. Guideline for disinfection and sterilization of prion-contaminated medical instruments. Infect. Control Hosp. Epidemiol. 2010, 31, 107–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tateishi, J.; Tashima, T.; Kitamoto, T. Practical methods for chemical inactivation of Creutzfeldt-Jakob disease pathogen. Microbiol. Immunol. 1991, 35, 163–166. [Google Scholar] [CrossRef]
- Rutala, W.A.; Weber, D.J. Healthcare Infection Control Practices Advisory Committee (HICPAC). Guideline for Disinfection and Sterilization in Healthcare Facilities. 2008. Available online: https://www.cdc.gov/infectioncontrol/pdf/guidelines/disinfection-guidelines-H.pdf (accessed on 10 December 2020).
- WHO. WHO Infection Control Guidelines for Transmissible Spongiform Encephalopathies. Report of a WHO Consultation, Geneva, Switzerland, 23–26 March 1999, WHO/CDS/CSR/APH/2000/3.. Available online: http://www.who.int/csr/resources/publications/bse/WHO_CDS_CSR_APH_2000_3/en/ (accessed on 7 December 2020).
- Erana, H.; Perez-Castro, M.A.; Garcia-Martinez, S.; Charco, J.M.; Lopez-Moreno, R.; Diaz-Dominguez, C.M.; Barrio, T.; Gonzalez-Miranda, E.; Castilla, J. A Novel, Reliable and Highly Versatile Method to Evaluate Different Prion Decontamination Procedures. Front. Bioeng. Biotechnol. 2020, 8, 589182. [Google Scholar] [CrossRef]
- Noda, M.; Sakai, Y.; Sakaguchi, Y.; Hayashi, N. Evaluation of low-temperature sterilization using hydrogen peroxide gas containing peracetic acid. Biocontrol. Sci. 2020, 25, 185–191. [Google Scholar] [CrossRef]
- Bentley, K.; Dove, B.K.; Parks, S.R.; Walker, J.T.; Bennett, A.M. Hydrogen peroxide vapour decontamination of surfaces artificially contaminated with norovirus surrogate feline calicivirus. J. Hosp. Infect. 2012, 80, 116–121. [Google Scholar] [CrossRef]
- Heckert, R.A.; Best, M.; Jordan, L.T.; Dulac, G.C.; Eddington, D.L.; Sterritt, W.G. Efficacy of vaporized hydrogen peroxide against exotic animal viruses. Appl. Environ. Microbiol. 1997, 63, 3916–3918. [Google Scholar] [CrossRef] [Green Version]
- Berrie, E.; Andrews, L.; Yezli, S.; Otter, J.A. Hydrogen peroxide vapour (HPV) inactivation of adenovirus. Lett. Appl. Microbiol. 2011, 52, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Otter, J.A.; Budde-Niekiel, A. Hydrogen peroxide vapor: A novel method for the environmental control of lactococcal bacteriophages. J. Food Prot. 2009, 72, 412–414. [Google Scholar] [CrossRef]
- Pottage, T.; Richardson, C.; Parks, S.; Walker, J.T.; Bennett, A.M. Evaluation of hydrogen peroxide gaseous disinfection systems to decontaminate viruses. J. Hosp. Infect. 2010, 74, 55–61. [Google Scholar] [CrossRef]
- Goyal, S.M.; Chander, Y.; Yezli, S.; Otter, J.A. Evaluating the virucidal efficacy of hydrogen peroxide vapour. J. Hosp. Infect. 2014, 86, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Fichet, G.; Antloga, K.; Comoy, E.; Deslys, J.P.; McDonnell, G. Prion inactivation using a new gaseous hydrogen peroxide sterilisation process. J. Hosp. Infect. 2007, 67, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Sakudo, A.; Yamashiro, R.; Harata, C. Effect of non-concentrated and concentrated vaporized hydrogen peroxide on scrapie prions. Pathogens 2020, 9, 947. [Google Scholar] [CrossRef] [PubMed]
- Alasri, A.; Roques, C.; Michel, G.; Cabassud, C.; Aptel, P. Bactericidal properties of peracetic acid and hydrogen peroxide, alone and in combination, and chlorine and formaldehyde against bacterial water strains. Can. J. Microbiol. 1992, 38, 635–642. [Google Scholar] [CrossRef]
- Leggett, M.J.; Schwarz, J.S.; Burke, P.A.; McDonnell, G.; Denyer, S.P.; Maillard, J.Y. Resistance to and killing by the sporicidal microbicide peracetic acid. J. Antimicrob. Chemother. 2015, 70, 773–779. [Google Scholar] [CrossRef]
- Leggett, M.J.; Schwarz, J.S.; Burke, P.A.; McDonnell, G.; Denyer, S.P.; Maillard, J.Y. Mechanism of sporicidal activity for the synergistic combination of peracetic acid and hydrogen peroxide. Appl. Environ. Microbiol. 2016, 82, 1035–1039. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Spore germination. Curr. Opin. Microbiol. 2003, 6, 550–556. [Google Scholar] [CrossRef]
- Lambert, R.J.; Johnston, M.D.; Hanlon, G.W.; Denyer, S.P. Theory of antimicrobial combinations: Biocide mixtures—synergy or addition? J. Appl. Microbiol. 2003, 94, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Vadrot, C.; Darbord, J.C. Quantitative evaluation of prion inactivation comparing steam sterilization and chemical sterilants: Proposed method for test standardization. J. Hosp. Infect. 2006, 64, 143–148. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for the Safe Transport of Infectious Substances and Diagnostic Specimens; World Health Organization: Geneva, Switzerland, 1997; WHO/EMC/97.3.; Available online: http://www.who.int/csr/resources/publications/biosafety/WHO_EMC_97_3_EN/en/ (accessed on 7 December 2020).
- Rogez-Kreuz, C.; Yousfi, R.; Soufflet, C.; Quadrio, I.; Yan, Z.X.; Huyot, V.; Aubenque, C.; Destrez, P.; Roth, K.; Roberts, C.; et al. Inactivation of animal and human prions by hydrogen peroxide gas plasma sterilization. Infect. Control Hosp. Epidemiol. 2009, 30, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Krebs, M.C.; Bécasse, P.; Verjat, D.; Darbord, J.C. Gas plasma sterilization: Relative efficacy of the hydrogen peroxide phase compared with that of the plasma phase. Int. J. Pharm. 1998, 160, 75–81. [Google Scholar] [CrossRef]
- Shintani, H. Is SteradR from J&J is truly plasma gas sterilizer? Pharmaceut. Reg. Aff. 2013, 3, e124. [Google Scholar] [CrossRef] [Green Version]
- Sakudo, A.; Iwamaru, Y.; Furusaki, K.; Haritani, M.; Onishi, R.; Imamura, M.; Yokoyama, T.; Yoshikawa, Y.; Onodera, T. Inactivation of scrapie prions by the electrically charged disinfectant CAC-717. Pathogens 2020, 9, 536. [Google Scholar] [CrossRef] [PubMed]
- Flechsig, E.; Hegyi, I.; Enari, M.; Schwarz, P.; Collinge, J.; Weissmann, C. Transmission of scrapie by steel-surface-bound prions. Mol. Med. 2001, 7, 679–684. [Google Scholar] [CrossRef]
- Murdoch, H.; Taylor, D.; Dickinson, J.; Walker, J.T.; Perrett, D.; Raven, N.D.; Sutton, J.M. Surface decontamination of surgical instruments: An ongoing dilemma. J. Hosp. Infect. 2006, 63, 432–438. [Google Scholar] [CrossRef]
- Pattison, I.H. The relative susceptibility of sheep, goats and mice to two types of the goat scrapie agent. Res. Vet. Sci. 1966, 7, 207–212. [Google Scholar] [CrossRef]
- Prusiner, S.B.; Cochran, S.P.; Groth, D.F.; Downey, D.E.; Bowman, K.A.; Martinez, H.M. Measurement of the scrapie agent using an incubation time interval assay. Ann. Neurol. 1982, 11, 353–358. [Google Scholar] [CrossRef]
- Derkinderen, P. Could it be that neurodegenerative diseases are infectious? Rev. Neurol. (Paris.) 2019, 175, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.G. Molecular pathology of neurodegenerative diseases: Principles and practice. J. Clin. Pathol. 2019, 72, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Jaunmuktane, Z.; Brandner, S. Invited review: The role of prion-like mechanisms in neurodegenerative diseases. Neuropathol. Appl. Neurobiol. 2020, 46, 522–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prusiner, S.B. Cell biology. A unifying role for prions in neurodegenerative diseases. Science 2012, 336, 1511–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraya Co., Ltd. Hydrogen Peroxide Gas Sterilizer STERIACE 100 SA-100. Available online: https://med.saraya.com/products/kigusenjo/45287.html (accessed on 19 October 2020).
- Sakudo, A.; Imanishi, Y.; Hirata, A.; Koga, Y.; Shintani, H. Effect of nitrogen gas plasma generated by a fast-pulsed power supply using a static induction thyristor on scrapie prion. Pathogens 2020, 9, 819. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Mean Incubation Time ± SEM 1 | N/N02 |
|---|---|---|
| Control | 191.0 ± 9.0 days | 6/6 |
| SD | >276 days 3 | 3/8 |
| QC | >297 days 4 | 2/8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakudo, A.; Anraku, D.; Itarashiki, T. Inactivation of Prions by Low-Temperature Sterilization Technology Using Vaporized Gas Derived from a Hydrogen Peroxide–Peracetic Acid Mixture. Pathogens 2021, 10, 24. https://doi.org/10.3390/pathogens10010024
Sakudo A, Anraku D, Itarashiki T. Inactivation of Prions by Low-Temperature Sterilization Technology Using Vaporized Gas Derived from a Hydrogen Peroxide–Peracetic Acid Mixture. Pathogens. 2021; 10(1):24. https://doi.org/10.3390/pathogens10010024
Chicago/Turabian StyleSakudo, Akikazu, Daiki Anraku, and Tomomasa Itarashiki. 2021. "Inactivation of Prions by Low-Temperature Sterilization Technology Using Vaporized Gas Derived from a Hydrogen Peroxide–Peracetic Acid Mixture" Pathogens 10, no. 1: 24. https://doi.org/10.3390/pathogens10010024