Skin Color in Apple Fruit (Malus × domestica): Genetic and Epigenetic Insights


 and
and
Abstract
1. Introduction
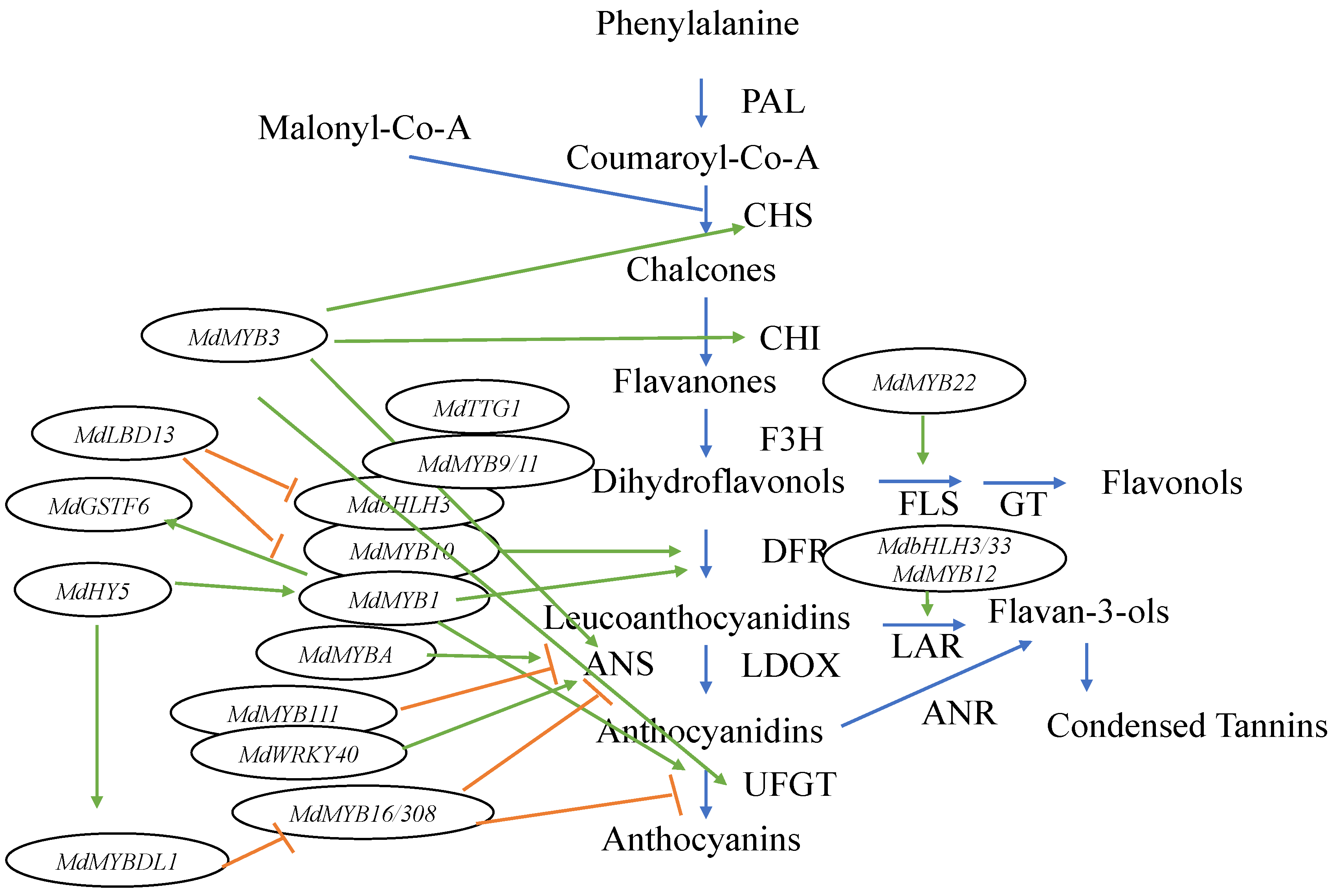
2. Anthocyanin Biosynthesis and Pigment Composition
3. Genetic Determinants of Apple Skin Color
3.1. MYB Transcription Factors Regulate Apple Skin Color
3.2. Genetic Mapping
3.3. Copy Number Variations
3.4. Transposable Elements
4. Epigenetic Determinants of Apple Skin Color
4.1. DNA Methylation and Demethylation
4.2. Histone Modifications
4.3. Small RNAs
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-aza-dC | 5-Aza-2′-deoxycytidine |
| ANS | Anthocyanidin synthase |
| CHI | Chalcone isomerase |
| CHS | Chalcone synthase |
| CNV | Copy number variation |
| CNVRs | Copy number variable regions |
| DEGs | Differentially expressed genes |
| DFR | Dihydroflavonol 4-reductase |
| DMG | Differentially methylated genes |
| DMR | Differentially methylated regions |
| DUS | Distinctness, uniformity, and stability |
| eTMs | Endogenous target mimics |
| F3H | Flavanone-3-hydroxylase |
| FLS | Flavonol synthase |
| GSTs | Glutathione S-transferases |
| LAR | Leucoanthocyanidin reductase |
| LDOX | Leucoanthocyanidin dioxygenase |
| LINE, SINE | Long and short interspersed nuclear elements |
| lncRNA | Long non-coding RNA |
| LRNs | LTR retrotransposons |
| LTR | Long terminal repeats |
| miRNAs | MicroRNAs |
| MRE | MYB recognition element |
| NGS | Next-generation sequencing |
| PA | Proanthocyanidin |
| phasiRNAs | Phased siRNAs |
| QTL | Quantitative trait locus |
| RdDM pathway | RNA-directed DNA methylation pathway |
| siRNAs | Small interfering RNAs |
| SNPs | Single Nucleotide Polymorphisms |
| sRNAs | Small RNAs |
| TE | Transposable elements |
| TF | Transcription factor |
| TSDs | Target site duplications |
| UFGT | UDP-glucose: flavonoid-3-O-glycosyltransferase |
| WGBS | Whole-genome bisulfite sequencing |
References
- Habben, J.; Schulte, E. Plant breeders’ rights for new fruit cultivars. Acta Hortic. 2000, 538, 673–676. [Google Scholar] [CrossRef]
- Bae, R.N.; Lee, S.K. Influence of chlorophyll, internal ethylene, and PAL on anthocyanin synthesis in “Fuji” apple. J. Korean Soc. Hortic. Sci. 1995, 36, 361–370. [Google Scholar]
- Lancaster, J.E.; Grant, J.E.; Lister, C.E.; Taylor, M.C. Skin Color in Apples—Influence of Copigmentation and Plastid Pigments on Shade and Darkness of Red Color in Five Genotypes. J. Am. Soc. Hortic. Sci. 1994, 119, 63–69. [Google Scholar] [CrossRef]
- Treutter, D. Biosynthesis of phenolic compounds and its regulation in apple. Plant Growth Regul. 2001, 34, 71–89. [Google Scholar] [CrossRef]
- Ben-Yehudah, G.; Korchinsky, R.; Redel, G.; Ovadya, R.; Oren-Shamir, M.; Cohen, Y. Colour accumulation patterns and the anthocyanin biosynthetic pathway in “Red Delicious” apple variants. J. Hortic. Sci. Biotechnol. 2005, 80, 187–192. [Google Scholar] [CrossRef]
- Cipollini, M.L.; Levey, D.J. Antifungal activity of Solanum fruit glycoalkaloids: Implications for frugivory and seed dispersal. Ecology 1997, 78, 799–809. [Google Scholar] [CrossRef]
- Steyn, W.J.; Wand, S.J.E.; Jacobs, G.; Rosecrance, R.C.; Roberts, S.C. Evidence for a photoprotective function of low-temperature-induced anthocyanin accumulation in apple and pear peel. Physiol. Plant. 2009, 136, 461–472. [Google Scholar] [CrossRef]
- Hoch, W.A.; Singsaas, E.L.; McCown, B.H. Resorption Protection. Anthocyanins Facilitate Nutrient Recovery in Autumn by Shielding Leaves from Potentially Damaging Light Levels. Plant Physiol. 2003, 133, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W. Anthocyanins in autumn leaf senescence. Adv. Bot. Res. 2002, 37, 147–165. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Xie, S.; Sockovie, E.; Khanizadeh, S. Which polyphenolic compounds contribute to the total antioxidant activities of apple? J. Agric. Food Chem. 2005, 53, 4989–4995. [Google Scholar] [CrossRef]
- Takos, A.M.; Ubi, B.E.; Robinson, S.P.; Walker, A.R. Condensed tannin biosynthesis genes are regulated separately from other flavonoid biosynthesis genes in apple fruit skin. Plant Sci. 2006, 170, 487–499. [Google Scholar] [CrossRef]
- Broun, P. Transcription factors as tools for metabolic engineering in plants. Curr. Opin. Plant Biol. 2004, 7, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Hichri, I.; Deluc, L.; Barrieu, F.; Bogs, J.; Mahjoub, A.; Regad, F.; Gallois, B.; Granier, T.; Trossat-Magnin, C.; Gomès, E.; et al. A single amino acid change within the R2 domain of the VvMYB5b transcription factor modulates affinity for protein partners and target promoters selectivity. BMC Plant Biol. 2011, 11, 117. [Google Scholar] [CrossRef]
- Takos, A.M.; Jaffe, F.W.; Jacob, S.R.; Bogs, J.; Robinson, S.P.; Walker, A.R. Light-Induced Expression of a MYB Gene Regulates Anthocyanin Biosynthesis in Red Apples. Plant Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef] [PubMed]
- Espley, R.V.; Bovy, A.; Bava, C.; Jaeger, S.R.; Tomes, S.; Norling, C.; Crawford, J.; Rowan, D.; McGhie, T.K.; Brendolise, C.; et al. Analysis of genetically modified red-fleshed apples reveals effects on growth and consumer attributes. Plant Biotechnol. J. 2013, 11, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Awad, M.A.; De Jager, A. Formation of flavonoids, especially anthocyanin and chlorogenic acid in “Jonagold” apple skin: Influences of growth regulators and fruit maturity. Sci. Hortic. 2002, 93, 257–266. [Google Scholar] [CrossRef]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef]
- Zipor, G.; Duarte, P.; Carqueijeiro, I.; Shahar, L.; Ovadia, R.; Teper-Bamnolker, P.; Eshel, D.; Levin, Y.; Doron-Faigenboim, A.; Sottomayor, M.; et al. In planta anthocyanin degradation by a vacuolar class III peroxidase in Brunfelsia calycina flowers. New Phytol. 2015, 205, 653–665. [Google Scholar] [CrossRef]
- Rogers, W.S. Rootstock effect on colour and size of apples. Rep. East Malling Res. Stn. 1926, 2, 16–32. [Google Scholar]
- Tukey, H.B.; Brase, K.D. Three year performance of sixteen varieties of apples on Malling IX rootstocks Proc. Am. Soc. Hort. Sci 1941, 38, 321–327. [Google Scholar]
- Blair, D.S. Rootstock and scion relationship in apple trees. Sci. Agric. 1938, 19, 85–94. [Google Scholar]
- Hewetson, F.N. Growth and yield of McIntosh apple trees as influenced by the use of various intermediate stem pieces. Proc. Am. Soc. Hortic. Sci. 1944, 45, 181–186. [Google Scholar]
- Upshalli, W.H. Malling Stocks and French Crab Seedlings as Stocks for Five Varieties of Apples. III. Sci. Agric. 1943, 23, 537–545. [Google Scholar]
- Larsen, F.E.; Fritts, R.; Olsen, K.L. Rootstock influence on “delicious” and “golden delicious” apple fruit quality at harvest and after storage. Sci. Hortic. 1985, 26, 339–349. [Google Scholar] [CrossRef]
- Parvaneh, T.; Abedi, B.; Davarynejad, G.H.; Ganji Moghadam, E. Enzyme activity, phenolic and flavonoid compounds in leaves of Iranian red flesh apple cultivars grown on different rootstocks. Sci. Hortic. 2019, 246, 862–870. [Google Scholar] [CrossRef]
- Ban, Y.; Honda, C.; Hatsuyama, Y.; Igarashi, M.; Bessho, H.; Moriguchi, T. Isolation and functional analysis of a MYB transcription factor gene that is a key regulator for the development of red coloration in apple skin. Plant Cell Physiol. 2007, 48, 958–970. [Google Scholar] [CrossRef] [PubMed]
- Espley, R.V.; Hellens, R.P.; Putterill, J.; Stevenson, D.E.; Kutty-Amma, S.; Allan, A.C. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 2007, 49, 414–427. [Google Scholar] [CrossRef]
- Li, Y.Y.; Mao, K.; Zhao, C.; Zhao, X.Y.; Zhang, H.L.; Shu, H.R.; Hao, Y.J. MdCOP1 ubiquitin E3 ligases interact with MdMYB1 to regulate light-induced anthocyanin biosynthesis and red fruit coloration in apple. Plant Physiol. 2012, 160, 1011–1022. [Google Scholar] [CrossRef]
- Yuan, K.; Wang, C.; Wang, J.; Xin, L.; Zhou, G.; Li, L.; Shen, G. Analysis of the MdMYB1 gene sequence and development of new molecular markers related to apple skin color and fruit-bearing traits. Mol. Genet. Genom. 2014, 289, 1257–1265. [Google Scholar] [CrossRef]
- Vimolmangkang, S.; Han, Y.; Wei, G.; Korban, S.S. An apple MYB transcription factor, MdMYB3, is involved in regulation of anthocyanin biosynthesis and flower development. BMC Plant Biol. 2013, 13, 176. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Wang, N.; Liu, J.; Qu, C.; Wang, Y.; Jiang, S.; Lu, N.; Wang, D.; Zhang, Z.; Chen, X. The molecular mechanism underlying anthocyanin metabolism in apple using the MdMYB16 and MdbHLH33 genes. Plant Mol. Biol. 2017, 94, 149–165. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, Y.; Sun, J.; Jiang, H.; Xu, H.; Wang, N.; Jiang, S.; Fang, H.; Zhang, Z.; Wang, Y.L.; et al. MdMYBDL1 employed by MdHY5 increases anthocyanin accumulation via repression of MdMYB16/308 in apple. Plant Sci. 2019, 283, 32–40. [Google Scholar] [CrossRef] [PubMed]
- An, X.H.; Tian, Y.; Chen, K.Q.; Liu, X.J.; Liu, D.D.; Xie, X.-B.; Cheng, C.G.; Cong, P.H.; Hao, Y.J. MdMYB9 and MdMYB11 are involved in the regulation of the ja-induced biosynthesis of anthocyanin and proanthocyanidin in apples. Plant Cell Physiol. 2015, 56, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Xu, H.; Jiang, S.; Zhang, Z.; Lu, N.; Qiu, H.; Qu, C.; Wang, Y.; Wu, S.; Chen, X. MYB12 and MYB22 play essential roles in proanthocyanidin and flavonol synthesis in red-fleshed apple (Malus sieversii f. niedzwetzkyana). Plant J. 2017, 90, 276–292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xu, H.; Yang, G.; Wang, N.; Zhang, J.; Wang, Y.; Jiang, S.; Fang, H.; Zhang, Z.; Chen, X. Molecular mechanism of MYB111 and WRKY40 involved in anthocyanin biosynthesis in red-fleshed apple callus. Plant Cell. Tissue Organ Cult. 2019, 139, 467–478. [Google Scholar] [CrossRef]
- Sun, Y.; Li, H.; Huang, J.R. Arabidopsis TT19 functions as a carrier to transport anthocyanin from the cytosol to tonoplasts. Mol. Plant 2012, 5, 387–400. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, I.; Liang, D.; Xu, K. Transcriptome analysis of an apple (Malus × domestica) yellow fruit somatic mutation identifies a gene network module highly associated with anthocyanin and epigenetic regulation. J. Exp. Bot. 2015, 66, 7359–7376. [Google Scholar] [CrossRef]
- Jiang, S.; Chen, M.; He, N.; Chen, X.; Wang, N.; Sun, Q.; Zhang, T.; Xu, H.; Fang, H.; Wang, Y.; et al. MdGSTF6, activated by MdMYB1, plays an essential role in anthocyanin accumulation in apple. Hortic. Res. 2019, 6, 40. [Google Scholar] [CrossRef]
- Li, H.H.; Liu, X.; An, J.P.; Hao, Y.J.; Wang, X.F.; You, C.X. Cloning and elucidation of the functional role of apple MdLBD13 in anthocyanin biosynthesis and nitrate assimilation. Plant Cell. Tissue Organ Cult. 2017, 130, 47–59. [Google Scholar] [CrossRef]
- Chagné, D.; Krieger, C.; Rassam, M.; Sullivan, M.; Fraser, J.; André, C.; Pindo, M.; Troggio, M.; Gardiner, S.E.; Henry, R.A.; et al. QTL and candidate gene mapping for polyphenolic composition in apple fruit. BMC Plant Biol. 2012, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- McClure, K.A.; Gong, Y.; Song, J.; Vinqvist-Tymchuk, M.; Campbell Palmer, L.; Fan, L.; Burgher-MacLellan, K.; Zhang, Z.; Celton, J.-M.; Forney, C.F.; et al. Genome-wide association studies in apple reveal loci of large effect controlling apple polyphenols. Hortic. Res. 2019, 6, 107. [Google Scholar] [CrossRef] [PubMed]
- Dunemann, F.; Kahnau, R.; Stange, I. Analysis of complex leaf and flower characters in Rhododendron using a molecular linkage map. Theor. Appl. Genet. 1999, 98, 1146–1155. [Google Scholar] [CrossRef]
- Moriya, S.; Kunihisa, M.; Okada, K.; Shimizu, T.; Honda, C.; Yamamoto, T.; Muranty, H.; Denancé, C.; Katayose, Y.; Iwata, H.; et al. Allelic composition of MdMYB1 drives red skin color intensity in apple (Malus × domestica Borkh.) and its application to breeding. Euphytica 2017, 213, 78. [Google Scholar] [CrossRef]
- Chagné, D.; Kirk, C.; How, N.; Whitworth, C.; Fontic, C.; Reig, G.; Sawyer, G.; Rouse, S.; Poles, L.; Gardiner, S.E.; et al. A functional genetic marker for apple red skin coloration across different environments. Tree Genet. Genomes 2016, 12, 67. [Google Scholar] [CrossRef]
- Gardner, K.M.; Brown, P.; Cooke, T.F.; Cann, S.; Costa, F.; Bustamante, C.; Velasco, R.; Troggio, M.; Myles, S. Fast and Cost-Effective Genetic Mapping in Apple Using Next-Generation Sequencing. G3 Genes Genomes Genet. 2014, 4, 1681–1687. [Google Scholar] [CrossRef] [PubMed]
- Lozano, L.; Iglesias, I.; Micheletti, D.; Troggio, M.; Kumar, S.; Volz, R.K.; Allan, A.C.; Chagné, D.; Gardiner, S.E. Feasibility of genome-wide association analysis using a small single nucleotide polymorphism panel in an apple breeding population segregating for fruit skin color. J. Am. Soc. Hortic. Sci. 2014, 139, 619–626. [Google Scholar] [CrossRef]
- Lijavetzky, D.; Cabezas, J.; Ibáñez, A.; Rodríguez, V.; Martínez-Zapater, J.M. High throughput SNP discovery and genotyping in grapevine (Vitis vinifera L.) by combining a re-sequencing approach and SNPlex technology. BMC Genom. 2007, 8, 1–11. [Google Scholar] [CrossRef]
- Chagné, D.; Carlisle, C.M.; Blond, C.; Volz, R.K.; Whitworth, C.J.; Oraguzie, N.C.; Crowhurst, R.N.; Allan, A.C.; Espley, R.V.; Hellens, R.P.; et al. Mapping a candidate gene (MdMYB10) for red flesh and foliage colour in apple. BMC Genom. 2007, 8, 212. [Google Scholar] [CrossRef]
- Lin-Wang, K.; Bolitho, K.; Grafton, K.; Kortstee, A.; Karunairetnam, S.; Mcghie, T.K.; Espley, R.V.; Hellens, R.P.; Allan, A.C. An R2R3 MYB transcription factor associated with regulation of the anthocyanin biosynthetic pathway in Rosaceae. BMC Plant Biol. 2010, 10, 50. [Google Scholar] [CrossRef]
- Zhu, Y.; Evans, K.; Peace, C. Utility testing of an apple skin color MdMYB1 marker in two progenies. Mol. Breed. 2011, 27, 525–532. [Google Scholar] [CrossRef]
- Chagné, D.; Lin-Wang, K.; Espley, R.V.; Volz, R.K.; How, N.M.; Rouse, S.; Brendolise, C.; Carlisle, C.M.; Kumar, S.; De Silva, N.; et al. An Ancient Duplication of Apple MYB Transcription Factors Is Responsible for Novel Red Fruit-Flesh Phenotypes. Plant Physiol. 2013, 161, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Volz, R.; Oraguzie, N.; Whitworth, C.; How, N.; Chagné, D.; Carlisle, C.; Gardiner, S. Red flesh breeding in apple—Progress and challenges. Acta Hortic. 2009, 814, 337–342. [Google Scholar] [CrossRef]
- Girirajan, S.; Campbell, C.D.; Eichler, E.E. Human Copy Number Variation and Complex Genetic Disease. Annu. Rev. Genet. 2011, 45, 203–226. [Google Scholar] [CrossRef]
- Boocock, J.; Chagné, D.; Merriman, T.R.; Black, M.A. The distribution and impact of common copy-number variation in the genome of the domesticated apple, Malus × domestica Borkh. BMC Genom. 2015, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Nayak, S.N.; May, G.D.; Jackson, S.A. Next-generation sequencing technologies and their implications for crop genetics and breeding. Trends Biotechnol. 2009, 27, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Espley, R.V.; Brendolise, C.; Chagné, D.; Kutty-Amma, S.; Green, S.; Volz, R.; Putterill, J.; Schouten, H.J.; Gardiner, S.E.; Hellens, R.P.; et al. Multiple repeats of a promoter segment causes transcription factor autoregulation in red apples. Plant Cell 2009, 21, 168–183. [Google Scholar] [CrossRef] [PubMed]
- Brendolise, C.; Espley, R.V.; Lin-Wang, K.; Laing, W.; Peng, Y.; McGhie, T.; Dejnoprat, S.; Tomes, S.; Hellens, R.P.; Allan, A.C. Multiple copies of a simple MYB-binding site confers trans-regulation by specific flavonoid-related R2R3 MYBs in diverse species. Front. Plant Sci. 2017, 8, 1864. [Google Scholar] [CrossRef]
- Kumar, A.; Bennetzen, J.L. Plant Retrotransposons. Annu. Rev. Genet. 1999, 33, 479–532. [Google Scholar] [CrossRef]
- Finnegan, D.J. Transposable elements. Curr. Opin. Genet. Dev. 1992, 2, 861–867. [Google Scholar] [CrossRef]
- Charlesworth, B.; Sniegowski, P.; Stephan, W. The evolutionary dynamics of repetitive DNA in eukaryotes. Nature 1994, 371, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Hadonou, A.M.; Gittins, J.R.; Hiles, E.R.; James, D.J. Two apple repetitive sequence elements: Characterisation and potential use as genetic markers. Euphytica 2003, 131, 177–187. [Google Scholar] [CrossRef]
- Hollister, J.D.; Smith, L.M.; Guo, Y.L.; Ott, F.; Weigel, D.; Gaut, B.S. Transposable elements and small RNAs contribute to gene expression divergence between Arabidopsis thaliana and Arabidopsis lyrata. Proc. Natl. Acad. Sci. USA 2011, 108, 2322–2327. [Google Scholar] [CrossRef] [PubMed]
- Galindo-González, L.; Mhiri, C.; Deyholos, M.K.; Grandbastien, M.A. LTR-retrotransposons in plants: Engines of evolution. Gene 2017, 626, 14–25. [Google Scholar] [CrossRef]
- Amyotte, S.G.; Tan, X.; Pennerman, K.; del Mar Jimenez-Gasco, M.; Klosterman, S.J.; Ma, L.J.; Dobinson, K.F.; Veronese, P. Transposable elements in phytopathogenic Verticillium spp.: Insights into genome evolution and inter- and intra-specific diversification. BMC Genom. 2012, 13, 314. [Google Scholar] [CrossRef] [PubMed]
- Nie, Q.; Qiao, G.; Peng, L.; Wen, X. Transcriptional activation of long terminal repeat retrotransposon sequences in the genome of pitaya under abiotic stress. Plant Physiol. Biochem. 2019, 135, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Rebollo, R.; Romanish, M.T.; Mager, D.L. Transposable Elements: An Abundant and Natural Source of Regulatory Sequences for Host Genes. Annu. Rev. Genet. 2012, 46, 21–42. [Google Scholar] [CrossRef] [PubMed]
- Daccord, N.; Celton, J.M.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E.; Van De Geest, H.; Bianco, L.; Micheletti, D.; Velasco, R.; et al. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 2017, 49, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef]
- Peace, C.P.; Bianco, L.; Troggio, M.; van de Weg, E.; Howard, N.P.; Cornille, A.; Durel, C.E.; Myles, S.; Migicovsky, Z.; Schaffer, R.J.; et al. Apple whole genome sequences: Recent advances and new prospects. Hortic. Res. 2019, 6, 59. [Google Scholar] [CrossRef]
- Telias, A.; Lin-Wang, K.; Stevenson, D.E.; Cooney, J.M.; Hellens, R.P.; Allan, A.C.; Hoover, E.E.; Bradeen, J.M. Apple skin patterning is associated with differential expression of MYb10. BMC Plant Biol. 2011, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, J.; Han, X.; Li, J.; Gao, Y.; Richards, C.M.; Zhang, C.; Tian, Y.; Liu, G.; Gul, H.; et al. A high-quality apple genome assembly reveals the association of a retrotransposon and red fruit colour. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Butelli, E.; Licciardello, C.; Zhang, Y.; Liu, J.; Mackay, S.; Bailey, P.; Reforgiato-Recupero, G.; Martin, C. Retrotransposons control fruit-specific, cold-dependent accumulation of anthocyanins in blood oranges. Plant Cell 2012, 24, 1242–1255. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirochika, H. Retrotransposon-Induced Mutations in Grape Skin Color. Science 2004, 304, 982. [Google Scholar] [CrossRef] [PubMed]
- Fournier-Level, A.; Lacombe, T.; Le Cunff, L.; Boursiquot, J.M.; This, P. Evolution of the VvMybA gene family, the major determinant of berry colour in cultivated grapevine (Vitis vinifera L.). Heredity 2010, 104, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Pelsy, F. Molecular and cellular mechanisms of diversity within grapevine varieties. Heredity 2010, 104, 331–340. [Google Scholar] [CrossRef]
- Russo, V.E.; Martienssen, R.A.; Riggs, A.D. Epigenetic Mechanisms of Gene Regulation; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1996. [Google Scholar]
- Chong, S.; Whitelaw, E. Epigenetic germline inheritance. Curr. Opin. Genet. Dev. 2004, 14, 692–696. [Google Scholar] [CrossRef]
- Basdeki, L.; Hagidimitriou, M. The Role of DNA Methylation in Perennial Plants. Not. Sci. Biol. 2019, 11, 1–11. [Google Scholar] [CrossRef]
- Tollefsbol, T. Handbook of Epigenetics: The New Molecular and Medical Genetics; Academic Press: Cambridge, MA, USA, 2017; ISBN 0128054778. [Google Scholar]
- Reinders, J.; Wulff, B.B.H.; Mirouze, M.; Marí-Ordóñez, A.; Dapp, M.; Rozhon, W.; Bucher, E.; Theiler, G.; Paszkowski, J. Compromised stability of DNA methylation and transposon immobilization in mosaic Arabidopsis epigenomes. Genes Dev. 2009, 23, 939–950. [Google Scholar] [CrossRef]
- Niederhuth, C.E.; Schmitz, R.J. Covering your bases: Inheritance of DNA methylation in plant genomes. Mol. Plant 2014, 7, 472–480. [Google Scholar] [CrossRef]
- Pikaard, C.S.; Scheid, O.M.; Kingston, R.E.; Tamkun, J.W.; Baulcombe, D.C.; Dean, C. Epigenetic Regulation in Plants. Cold Spring Harb. Perspect. Biol. 2014, 6, a019315. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Lang, Z.; Genetics, P.M. The mechanism and function of active DNA demethylation in plants. J. Integr. Plant Biol. 2019, 62, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Quadrana, L.; Almeida, J.; Asís, R.; Duffy, T.; Dominguez, P.G.; Bermúdez, L.; Conti, G.; Corrêa Da Silva, J.V.; Peralta, I.E.; Colot, V.; et al. Natural occurring epialleles determine vitamin e accumulation in tomato fruits. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liu, R.; Niu, Q.; Tang, K.; Zhang, B.; Zhang, H.; Chen, K.; Zhu, J.K.; Lang, Z. Global increase in DNA methylation during orange fruit development and ripening. Proc. Natl. Acad. Sci. USA 2019, 116, 1430–1436. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.H.; Sun, Q.G.; Chen, M.; Wang, N.; Xu, H.F.; Fang, H.C.; Wang, Y.C.; Zhang, Z.Y.; Chen, X. Sen Methylome and transcriptome analyses of apple fruit somatic mutations reveal the difference of red phenotype. BMC Genom. 2019, 20, 1–13. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, N.; Chen, M.; Zhang, R.; Sun, Q.; Xu, H.; Zhang, Z.; Wang, Y.; Sui, X.; Wang, S.; et al. Methylation of MdMYB1 locus mediated by RdDM pathway regulates anthocyanin biosynthesis in apple. Plant Biotechnol. J. 2020, 18, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Jing, C.; Chang, B.; Yan, J.; Liang, B.; Liu, L.; Yang, Y.; Zhao, Z. The effect of promoter methylation on MdMYB1 expression determines the level of anthocyanin accumulation in skins of two non-red apple cultivars. BMC Plant Biol. 2018, 18, 1–14. [Google Scholar] [CrossRef]
- Li, W.F.; Ning, G.X.; Mao, J.; Guo, Z.G.; Zhou, Q.; Chen, B.H. Whole-genome DNA methylation patterns and complex associations with gene expression associated with anthocyanin biosynthesis in apple fruit skin. Planta 2019, 250, 1833–1847. [Google Scholar] [CrossRef] [PubMed]
- Li, W.F.; Mao, J.; Yang, S.J.; Guo, Z.G.; Ma, Z.H.; Dawuda, M.M.; Zuo, C.W.; Chu, M.Y.; Chen, B.H. Anthocyanin accumulation correlates with hormones in the fruit skin of “Red Delicious” and its four generation bud sport mutants. BMC Plant Biol. 2018, 18, 1–15. [Google Scholar] [CrossRef]
- Xu, J.; Tanino, K.K.; Horner, K.N.; Robinson, S.J. Quantitative trait variation is revealed in a novel hypomethylated population of woodland strawberry (Fragaria vesca). BMC Plant Biol. 2016, 16, 1–17. [Google Scholar] [CrossRef]
- Mossman, D.; Kim, K.T.; Scott, R.J. Demethylation by 5-aza-2′-deoxycytidine in colorectal cancer cells targets genomic DNA whilst promoter CpG island methylation persists. BMC Cancer 2010, 10, 366. [Google Scholar] [CrossRef]
- Ma, C.; Liang, B.; Chang, B.; Liu, L.; Yan, J.; Yang, Y.; Zhao, Z. Transcriptome profiling reveals transcriptional regulation by DNA methyltransferase inhibitor 5-aza-2′-deoxycytidine enhancing red pigmentation in bagged “granny smith” apples (Malus domestica). Int. J. Mol. Sci. 2018, 19, 3133. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Tuan, P.A.; Saito, T.; Honda, C.; Hatsuyama, Y.; Ito, A.; Moriguchi, T. Epigenetic regulation of MdMYB1 is associated with paper bagging-induced red pigmentation of apples. Planta 2016, 244, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Kim, G.H.; Choi, C. Differential gene expression and epigenetic analyses between striped and blushed skinned sports of “Fuji” apple. Sci. Hortic. 2020, 261, 108944. [Google Scholar] [CrossRef]
- Springer, N.M. Epigenetics and crop improvement. Trends Genet. 2013, 29, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Hasbún, R.; Valledor, L.; Berdasco, M.; Santamaría, E.; Cañal, M.J.; Rodríguez, R.; Ríos, D.; Sánchez, M. In vitro proliferation and genome dna methylation in adult chestnuts. Acta Hortic. 2005, 693, 333–340. [Google Scholar] [CrossRef]
- Bucher, E.; Kong, J.; Teyssier, E.; Gallusci, P. Epigenetic Regulations of Fleshy Fruit Development and Ripening and Their Potential Applications to Breeding Strategies. Adv. Bot. Res. 2018, 88. [Google Scholar]
- Hu, G.; Cui, K.; Northrup, D.; Liu, C.; Wang, C.; Tang, Q.; Ge, K.; Levens, D.; Crane-Robinson, C.; Zhao, K. H2A.Z facilitates access of active and repressive complexes to chromatin in embryonic stem cell self-renewal and differentiation. Cell Stem Cell 2013, 12, 180–192. [Google Scholar] [CrossRef]
- Dai, X.; Bai, Y.; Zhao, L.; Dou, X.; Liu, Y.; Wang, L.; Li, Y.; Li, W.; Hui, Y.; Huang, X.; et al. H2A.Z Represses Gene Expression by Modulating Promoter Nucleosome Structure and Enhancer Histone Modifications in Arabidopsis. Mol. Plant 2017, 10, 1274–1292. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, M.; Chai, M.; He, Q.; Huang, X.; Zhao, L.; Qin, Y. Epigenetic regulation of anthocyanin biosynthesis by an antagonistic interaction between H2A.Z and H3K4me3. New Phytol. 2018, 221, 295–308. [Google Scholar] [CrossRef]
- Zheng, T.; Tan, W.; Yang, H.; Zhang, L.; Li, T.; Liu, B.; Zhang, D.; Lin, H. Regulation of anthocyanin accumulation via MYB75/HAT1/TPL-mediated transcriptional repression. PLoS Genet. 2019, 15, 1–28. [Google Scholar] [CrossRef]
- Hernandez, J.M.; Feller, A.; Morohashi, K.; Frame, K.; Grotewold, E. The basic helix-loop-helix domain of maize R links transcriptional regulation and histone modifications by recruitment of an EMSY-related factor. Proc. Natl. Acad. Sci. USA 2007, 104, 17222–17227. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zeng, Z.; Liu, Z.; Xia, R. Small RNAs, emerging regulators critical for the development of horticultural traits. Hortic. Res. 2018, 5, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Xia, R.; Zhu, H.; An, Y.-Q.; Beers, E.P.; Liu, Z. Apple miRNAs and tasiRNAs with novel regulatory networks. Genome Biol. 2012, 13, R47. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Shen, J.; Liu, H.; Li, F.; Ding, N.; Gao, C.; Pattanaik, S.; Patra, B.; Li, R.; Yuan, L. Small tandem target mimic-mediated blockage of microRNA858 induces anthocyanin accumulation in tomato. Planta 2015, 242, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Ma, H.; Zhang, J.; Wu, T.; Song, T.; Tian, J.; Yao, Y. Systematic identification of long noncoding RNAs expressed during light-induced anthocyanin accumulation in apple fruit. Plant J. 2019, 100, 572–590. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Related Gene | CNV Length | LG | Reference |
|---|---|---|---|
| MdMYB10 | Five direct tandem repeats of a 23bp sequence | 9 | Espley et al. [28] |
| MdMYB110a | 930-bp region upstream | 17 | Chagné et al. [52] |
| Related Gene | Over (+) or under (−) Methylated | Effect on Transcription | Reference |
| MdMYB10 | + | Reduces MdMYB10 transcription and anthocyanin production. | Telias et al. [71]; El-Sharkawy et al. [38] |
| MdANS, MdF3H | + | Modifies both regulatory and structural genes in the anthocyanin pathway. | Jiang et al. [87] |
| MdMYB114 | + | Regulates anthocyanin biosynthesis in fully red apples | Jiang et al. [87] |
| MdMYB1 | − | Formation of red pigment in the skin of Granny Smith. | Ma et al. [89] |
| CHS, PAL, F3’H, PER,4CL, CYP98A, CCoAOMT | − | Transcriptional activation and increase in anthocyanin accumulation. | Li et al. [90] |
| MdbHLH74 | − | Transcriptional repression, inhibiting accumulation of anthocyanin. | Li et al. [90] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Celton, J.-M.; Buck-Sorlin, G.; Balzergue, S.; Bucher, E.; Laurens, F. Skin Color in Apple Fruit (Malus × domestica): Genetic and Epigenetic Insights. Epigenomes 2020, 4, 13. https://doi.org/10.3390/epigenomes4030013
Wang W, Celton J-M, Buck-Sorlin G, Balzergue S, Bucher E, Laurens F. Skin Color in Apple Fruit (Malus × domestica): Genetic and Epigenetic Insights. Epigenomes. 2020; 4(3):13. https://doi.org/10.3390/epigenomes4030013
Chicago/Turabian StyleWang, Wuqian, Jean-Marc Celton, Gerhard Buck-Sorlin, Sandrine Balzergue, Etienne Bucher, and François Laurens. 2020. "Skin Color in Apple Fruit (Malus × domestica): Genetic and Epigenetic Insights" Epigenomes 4, no. 3: 13. https://doi.org/10.3390/epigenomes4030013
APA StyleWang, W., Celton, J.-M., Buck-Sorlin, G., Balzergue, S., Bucher, E., & Laurens, F. (2020). Skin Color in Apple Fruit (Malus × domestica): Genetic and Epigenetic Insights. Epigenomes, 4(3), 13. https://doi.org/10.3390/epigenomes4030013