Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii





Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Colony
2.2. Bacteria Isolation and Culture Conditions
2.3. Larval Oral Bioassay
2.4. Larval and Pupal Dipping Bioassay
2.5. Pupal Direct-Spray Bioassay
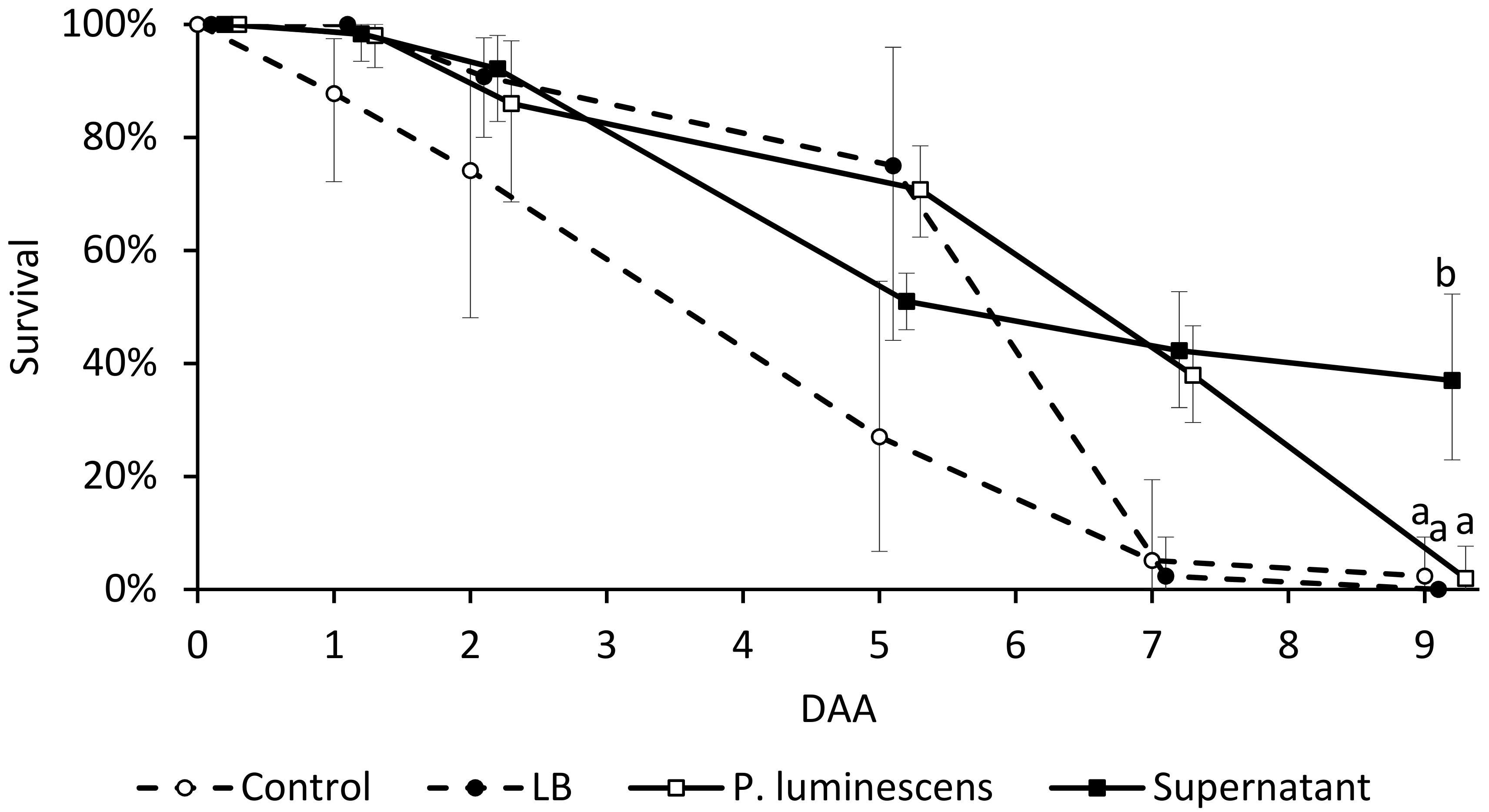
2.6. Adult Insect Bioassay
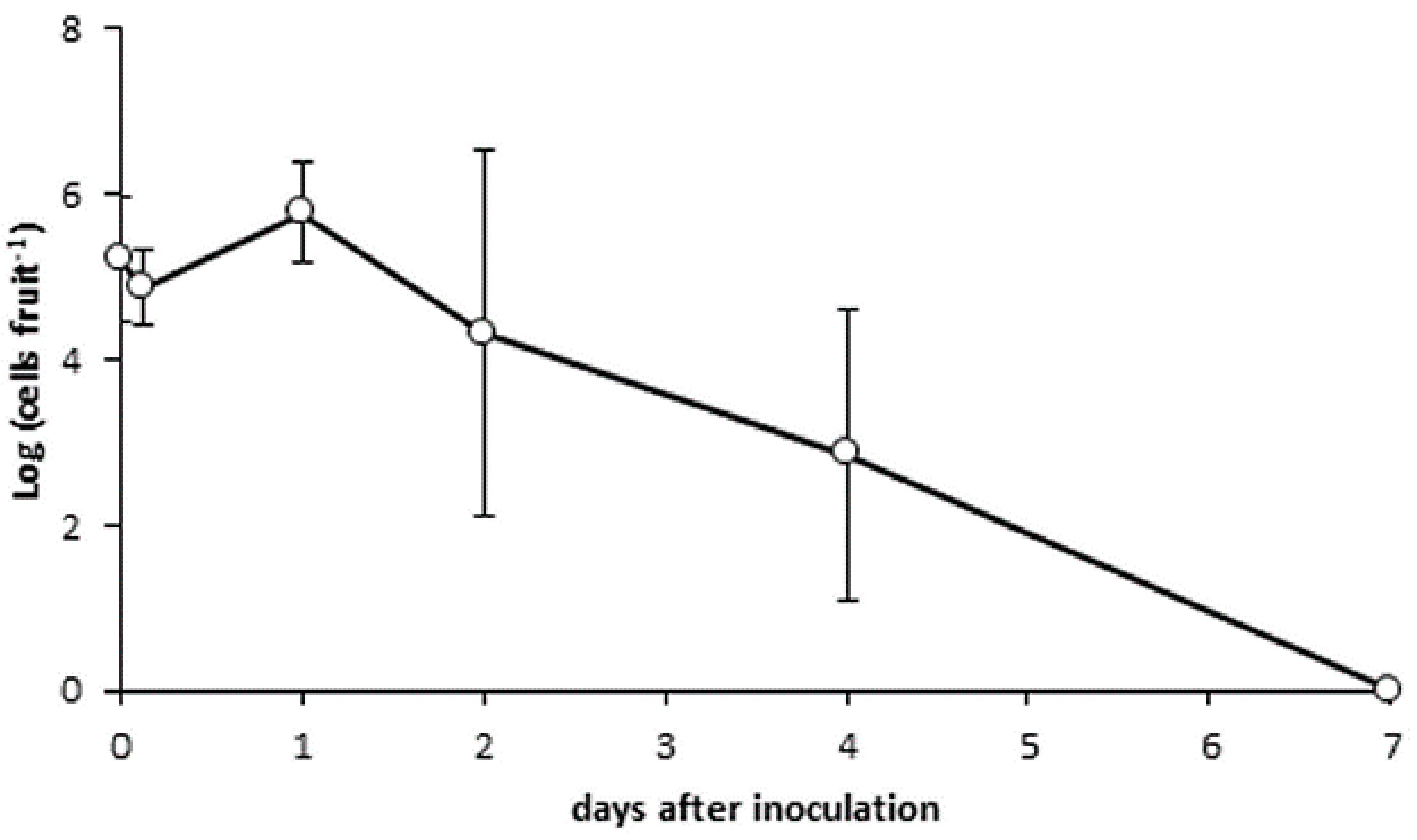
2.7. Bacterial Persistence on Cherry Fruit
2.8. Data Analysis
3. Results
3.1. Larval Oral Bioassay
3.2. Larval and Pupal Dipping Bioassay
3.3. Pupal Direct-Spray Bioassay
3.4. Adult Insect Bioassay
3.5. Bacterial Persistence on Cherry Skin
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cini, A.; Ioratti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.D.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Bellamy, D.E.; Sisterson, M.S.; Walse, S.S. Quantifying host potentials: Indexing postharvest fresh fruits for spotted wing drosophila, Drosophila suzukii. PLoS ONE 2013, 8, e61227. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Van Steenwyk, R.A.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufeld, P. In focus: Spotted wing drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Sato, R. Bionomics of the cherry drosophila, Drosophila suzukii Matsumura (Diptera: Drosophilidae) in Fukushima prefecture (Japan). Ann. Rep. Soc. Plant Prot. North Jpn. 1995, 46, 164–172. [Google Scholar]
- Goodhue, R.E.; Bolda, M.; Farnsworth, D.; Williams, J.C.; Zalom, F.G. Spotted wing drosophila infestation of California strawberries and raspberries: Economic analysis of potential revenue losses and control costs. Pest Manag. Sci. 2011, 67, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Haye, T.; Girod, P.; Cuthbertson, A.G.S.; Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Baroffio, C.; Zhang, J.P.; Desneux, N. Current SWD IPM tactics and their practical implementation in fruit crops across different regions around the world. J. Pest Sci. 2016, 89, 643–651. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Haviland, D.R.; Beers, E.H. Chemical control programs for Drosophila suzukii that comply with international limitations on pesticide residues for exported sweet cherries. J. Integr. Pest Manag. 2012, 3, F1–F6. [Google Scholar] [CrossRef]
- Stark, J.D.; Banks, J.E. Population-level effects of pesticides and other toxicants on arthropods. Annu. Rev. Entomol. 2003, 48, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Rahoo, A.M.; Mukhtar, T.; Gowen, S.R.; Pembroke, B. Virulence of entomopathogenic bacteria Xenorhabdus bovienii and Photorhabdus luminescens against Galleria mellonella larvae. Pak. J. Zool. 2011, 43, 543–548. [Google Scholar]
- Miller, B.; Anfora, G.; Buffington, M.; Daane, K.M.; Dalton, D.T.; Hoelmer, K.M.; Rossi Stacconi, M.V.; Grassi, A.; Ioriatti, C.; Loni, A.; et al. Seasonal occurrence of resident parasitoids associated with Drosophila suzukii in two small fruit production regions of Italy and the USA. Bull. Insectol. 2015, 68, 255–263. [Google Scholar]
- Wang, X.-G.; Stewart, T.J.; Biondi, A.; Chavez, B.A.; Ingels, C.; Caprile, J.; Grant, J.A.; Walton, V.M.; Daane, K.M. Population dynamics and ecology of Drosophila suzukii. J. Pest Sci. 2016, 89, 701–712. [Google Scholar] [CrossRef]
- Renkema, J.M.; Telfer, Z.; Gariepy, T.; Hallett, R.H. Dalotia coriaria as a predator of Drosophila suzukii: Functional responses, reduced fruit infestation and molecular diagnostics. Biol. Control 2015, 89, 1–10. [Google Scholar] [CrossRef]
- Woltz, J.; Donahue, K.; Bruck, D.; Lee, J. Efficacy of commercially available predators, nematodes and fungal entomopathogens for augmentative control of Drosophila suzukii. J. Appl. Entomol. 2015, 139, 759–770. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Collins, D.A.; Blackburn, L.F.; Audsley, N.; Bell, H.A. Preliminary screening of potential control products against Drosophila suzukii. Insects 2014, 5, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, A.G.S.; Audsley, N. Further screening of entomopathogenic fungi and nematodes as control agents for Drosophila suzukii. Insects 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Naranjo-Lázaro, J.M.; Mellín-Rosas, M.A.; González-Padilla, V.D.; Sánchez-González, J.A.; Moreno-Carrillo, G.; Arredondo-Bernal, H.C. Susceptibility of Drosophila suzukii Matsumura (Diptera: Drosophilidae) to entomophatogenic fungi. Southwest. Entomol. 2014, 39, 201–203. [Google Scholar] [CrossRef]
- Wise, J.C.; Van Woerkom, A.H.; Wheeler, C.E.; Isaacs, R. IR-4 Attract N Kill tactics for control of spotted wing drosophila in blueberry. Arthropod Manag. Tests 2018, 43. [Google Scholar] [CrossRef]
- EU-Pesticides-Database. European Union Pesticides Database. 2018. Available online: http://ec.europa.eu/food/plant/pesticides/eu-311 pesticides-database (accessed on 14 August 2018).
- Mahar, A.; Jan, N.; Mahar, G.M.; Mahar, A.Q. Control of insects with entomopathogenic bacterium Xenorhabdus nematophila and its toxic secretions. Int. J. Agric. Biol. 2008, 10, 52–56. [Google Scholar]
- Ruiu, L. Insect pathogenic bacteria in integrated pest management. Insects 2015, 6, 352–367. [Google Scholar] [CrossRef] [PubMed]
- Fanning, P.D.; Grieshop, M.J.; Isaacs, R. Efficacy of biopesticides on spotted wing drosophila, Drosophila suzukii Matsumura in fall red raspberries. J. Appl. Entomol. 2018, 142, 26–32. [Google Scholar] [CrossRef]
- Cahenzli, F.; Strack, T.; Daniel, C. Screening of 25 different natural crop protection products against Drosophila suzukii. J. Appl. Entomol. 2018, 142, 563–577. [Google Scholar] [CrossRef]
- Blackburn, M.; Golubeva, E.; Bowen, D.; Ffrench-Constant, R.H. A novel insecticidal toxin from Photorhabdus luminescens, toxin complex a (Tca), and its histopathological effects on the midgut of Manduca sexta. Appl. Environ. Microbiol. 1998, 64, 3036–3041. [Google Scholar] [PubMed]
- Waterfield, N.; Dowling, A.; Sharma, S.; Daborn, P.J.; Potter, U.; Ffrench-Constant, R.H. Oral toxicity of Photorhabdus luminescens W14 toxin complexes in Escherichia coli. Appl. Environ. Microbiol. 2001, 67, 5017–5024. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Fatig, R.O.; Orr, G.L.; Schafer, G.W.; Strickland, J.A.; Sukhapinda, K.; Woodsworth, A.T.; Petell, J.K. Photorhabdus luminescens W-14 insecticidal activity consists of at least two similar but distinct proteins. Purification and characterization of Toxin A and Toxin B. J. Biol. Chem. 1999, 274, 9836–9842. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, R.; Bhatnagar, R.K. Insecticidal toxic proteins produced by Photorhabdus luminescens akhurstii, a symbiont of Heterorhabditis indica. J. Nematol. 2002, 34, 23–27. [Google Scholar] [PubMed]
- Mahar, A.N.; Jan, N.D.; Mahar, A.Q.; Mahar, G.M.; Hullio, M.H.; Lanjar, A.G. Efficacy of entomopathogenic bacterium Photorhabdus luminescens and its metabolites against diamondback moth Plutella xylostella larvae on chinese cabbage and artificial diet. Pak. J. Nematol. 2008, 26, 69–82. [Google Scholar]
- Blackburn, M.B.; Domek, J.M.; Gelman, D.B.; Hu, J.S. The broadly insecticidal Photorhabdus luminescens toxin complex a (Tca): Activity against the Colorado potato beetle, Leptinotarsa decemlineata, and sweet potato whitefly, Bemisia tabaci. J. Insect Sci. 2005, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Dowling, A.; Waterfield, N.R. Insecticidal toxins from Photorhabdus bacteria and their potential use in agriculture. Toxicon 2007, 49, 436–451. [Google Scholar]
- Shawer, R.; Tonina, L.; Tirello, P.; Duso, C.; Mori, N. Laboratory and field trials to identify effective chemical control strategies for integrated management of Drosophila suzukii in European cherry orchards. Crop Prot. 2018, 103, 73–80. [Google Scholar] [CrossRef]
- Tonina, L.; Mori, N.; Giomi, F.; Battisti, A. Development of Drosophila suzukii. J. Pest Sci. 2016, 89, 667–678. [Google Scholar] [CrossRef]
- Kehres, J.; Denon, D.; Mauléon, H. A simple technique to estimate, in situ, population densities of an entomopathogenic nematode (Heterorhabditis indica) in sandy soils. Nematology 2001, 3, 285–287. [Google Scholar] [CrossRef]
- Sicard, M.; Hering, S.; Schulte, R.; Gaudriault, S.; Schulenburg, H. The effect of Photorhabdus luminescens (Enterobacteriaceae) on the survival, development, reproduction and behaviour of Caenorhabditis elegans (Nematoda: Rhabditidae). Environ. Microbiol. 2007, 9, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Berry, R.E.; Blouin, M.S. Identification of symbiotic bacteria (Photorhabdus and Xenorhabdus) from the entomopathogenic nematodes Heterorhabditis marelatus and Steinernema oregonense based on 16S rDNA sequence. J. Invertebr. Pathol. 2001, 77, 87–91. [Google Scholar] [CrossRef] [PubMed]
- da Silva, O.S.; Prado, G.R.; da Silva, J.L.R.; Silva, C.E.; da Costa, M.; Heermann, R. Oral toxicity of Photorhabdus luminescens and Xenorhabdus nematophila (Enterobacteriaceae) against Aedes aegypti (Diptera: Culicidae). Parasitol. Res. 2013, 112, 2891–2896. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Instructions for Determining the Susceptibility or Resistance of Mosquito Larvae to Insecticides. 1981. Available online: http://apps.who.int/iris/bitstream/handle/10665/69615/WHO?sequence=1 (accessed on 14 August 2018).
- Kanzawa, T. Studies on Drosophila suzukii Mats. Kofu. Rev. Appl. Entomol. 1939, 29, 622. [Google Scholar]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.-S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Yee, W.L.; Alston, D.G. Effects of spinosad, spinosad bait, and chloronicotinyl insecticides on mortality and control of adult and larval western cherry fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 2006, 99, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Waterfield, N.R.; Ciche, T.; Clarke, D. Photorhabdus and a host of hosts. Annu. Rev. Microbiol. 2009, 63, 557–574. [Google Scholar] [CrossRef] [PubMed]
- Sharad-Mohan, A.; Gaur, H.S. Successful management of mango mealy bug, Drosicha mangiferae by Photorhabdus luminescens, a symbiotic bacterium from entomopathogenic nematode Heterorhabditis indica. Int. J. Nematol. 2003, 14, 195–198. [Google Scholar]
- Uma, G.P.; Prabhuraj, A. Bio-efficacy of Photorhabdus luminescens, a symbiotic bacterium against Thrips palmi Karny (Thripidae: Thysanoptera). J. Biopestic. 2010, 3, 458–462. [Google Scholar]
- Musser, F.R.; Nyrop, J.P.; Shelton, A.M. Integrating biological and chemical controls in decision making: European corn borer (Lepidoptera: Crambidae) control in sweet corn as an example. J. Econ. Entomol. 2006, 99, 1538–1549. [Google Scholar] [CrossRef] [PubMed]
- Satinder, K.B.; Verma, M.; Tyagi, R.D.; Valéro, J.R. Recent advances in downstream processing and formulations of Bacillus thuringiensis based biopesticides. Process Biochem. 2006, 41, 323–342. [Google Scholar]
- Ramesh, A.K.; Prabhuraj, A.; Ashoka, J.; Hanchinal, S.G.; Sharanagouda, H. Generation and evaluation of nanoparticles of supernatant of Photorhabdus luminescens (Thomas and Poinar) against mite and aphid pests of cotton for enhanced efficacy. Curr. Sci. 2017, 112, 2312–2316. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Concentration (Cells mL−1) | Larvae | Pupae | Adults | Immatures | Total Individuals | |
|---|---|---|---|---|---|---|
| 2 DAA | 4 DAA | 10 DAA | 10 DAA | 10 DAA | 10 DAA | |
| 3.5 × 108 | 10.0 (10.0–10.0) a | 36.7 (8.0–65.0) a | 10.0 (−33.0–53.0) a | 50.0 (25.2–74.8) a | 46.7 (8.7–84.6) a | 96.7 (82.3–111) a |
| 3.5 × 107 | 3.3 (−11–17.6) a | 33.3 (−4.6–71.3) ab | 3.3 (−11.0–177.0) a | 53.3 (−22.6–129.0) a | 36.7 (−15–88.4) a | 90.0 (65.1–114.8) ab |
| 3.5 × 106 | 6.7 (−22–35.4) a | 10.0 (−14.8–34.8) c | 10.0 (−14.8–34.8) a | 53.3 (−22.6–129.0) a | 20.0 (−14.8–34.8) ab | 73.3 (44.6–102.0) bc |
| 3.5 × 105 | 6.7 (−22–35.4) a | 13.3 (−1.0–27.7) bc | 0.0 (0.0–0.0) a | 53.3 (1.6–105.0) a | 13.3 (−1.0–27.7) b | 66.7 (9.3–124.0) c |
| 3.5 × 104 | 0.0 (0.0–0.0) a | 6.7 (−22.0–35.4) c | 6.7 (−22.0–35.4) a | 46.7 (18.0–75.4) a | 13.3 (−15.4–42.0) b | 60.0 (60.0–60.0) c |
| 3.5 × 103 | 0.0 (0.0–0.0) a | 10.0 (−33.0–53.0) c | 3.3 (−11.0–17.7) a | 46.7 (8.7–84.6) a | 13.3 (−24.6–51.3) b | 60.0 (10.3–109.7) c |
| Control | 3.3 (−11.0–17.7) a | 6.7 (−7.6–21.0) c | 0.0 (0.0–0.0) a | 6.7 (−7.7–21.0) b | 6.7 (−7.7–21.0) b | 13.3 (−1.0–27.7) d |
| Treatment on Larvae | Larvae, 4 DAA | Immatures, 10 DAA | Total Individuals, 10 DAA |
| Bacterial supernatant | 24 (11.6–32.4) a | 44 (28.4–55.6) a | 74 (51.6–92.4) a |
| Supernatant 10-fold dilution | 12 (−1.6–25.6) b | 18 (1.8–34.2) b | 34 (15.2–52.8) b |
| Control | 4 (−2.8–10.8) b | 12 (1.6–22.4) b | 28 (17.6–38.4) b |
| Treatment on Pupae | Pupae, 9 DAA | Adults, 9 DAA | Total Individuals, 9 DAA |
| Bacterial supernatant | 38 (17.6–58.4) a | 36 (15.2–56.8) a | 74 (59.8–88.2) a |
| Supernatant 10-fold dilution | 26 (9.3–42.7) a | 16 (9.2–22.8) b | 42 (25.8–58.2) b |
| Control | 26 (19.2–32.8) a | 6 (−0.8–12.8) b | 32 (26.4–37.6) b |
| Concentration (Cells mL−1) | Larvae | Pupae | Adults | Immatures | Total Individuals | |
|---|---|---|---|---|---|---|
| 2 DAA | 4 DAA | 9 DAA | 10 DAA | 10 DAA | 10 DAA | |
| 3.5 × 108 | 6.7 (−7.7–21.0) a | 10.0 (10.0–10.0) a | 60.0 (−5.0–125.7) a | 16.7 (−35.0–68.4) a | 70.0 (4.3–135.7) a | 86.7 (58.0–115.4) a |
| 3.5 × 107 | 3.3 (−11.0–17.7) a | 10.0 (−14.0–34.8) a | 36.7 (22.3–51) ab | 16.7 (2.3–31.0) a | 46.7 (8.7–84.6) ab | 63.3 (11.6–115) ab |
| 3.5 × 106 | 3.3 (−11.0–17.7) a | 3.3 (−11.0–17.7) a | 16.7 (−55.0–88.4) b | 30.0 (−13.0–73.0) a | 20.0 (−45.7–85.7) b | 50.0 (−15.7–115.7) bc |
| Control | 3.3 (−11.0–17.7) a | 3.3 (−11.0–17.7) a | 13.3 (−1.0–27.7) b | 10.0 (10.0–10.0) a | 16.7 (2.3–31) b | 26.7 (12.3–41.0) c |
| Concentration (Cells mL−1) | Pupae | Adults | Total Individuals |
|---|---|---|---|
| 3.5 × 108 | 46.7 (32.3–61.0) a | 26.7 (−45.0–98.4) a | 73.3 (16.0–130.7) a |
| 3.5 × 107 | 46.7 (8.7–84.6) a | 23.3 (−39.2–85.8) a | 70.0 (4.3–135.7) a |
| 3.5 × 106 | 43.3 (5.4–81.3) ab | 23.3 (9.0–37.7) a | 66.7 (15.0–118.4) ab |
| Control | 23.3 (9.0–37.7) b | 6.7 (−7.7–21.0) a | 30.0 (5.2–54.8) b |
| Concentration (Cells mL−1) | Pupae | Adults | Total Individuals |
|---|---|---|---|
| 3.5 × 108 | 63.3 (6.0–120.7) a | 36.7 (−20.7–94.0) a | 100.0 (100.0–100.0) a |
| 3.5 × 107 | 50.0 (−15.7–115.7) a | 50.0 (−15.7–115.7) a | 100.0 (100.0–100.0) a |
| 3.5 × 106 | 63.3 (34.6–92.0) a | 30.0 (5.2–54.8) ab | 93.3 (79.0–107.7) ab |
| 3.5 × 105 | 63.3 (49.0–77.7) a | 26.7 (12.3–41.0) ab | 90.0 (65.2–114.8) ab |
| 3.5 × 104 | 56.7 (42.3–71.0) a | 30.0 (5.2–54.8) ab | 86.7 (58.0–115.4) ab |
| 3.5 × 103 | 50.0 (7.0–93.0) a | 33.3 (19.0–47.7) ab | 83.3 (45.4–121.3) b |
| Control | 10.0 (10.0–10.0) b | 10.0 (10.0–10.0) b | 20.0 (20.0–20.0) c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shawer, R.; Donati, I.; Cellini, A.; Spinelli, F.; Mori, N. Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii. Insects 2018, 9, 148. https://doi.org/10.3390/insects9040148
Shawer R, Donati I, Cellini A, Spinelli F, Mori N. Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii. Insects. 2018; 9(4):148. https://doi.org/10.3390/insects9040148
Chicago/Turabian StyleShawer, Rady, Irene Donati, Antonio Cellini, Francesco Spinelli, and Nicola Mori. 2018. "Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii" Insects 9, no. 4: 148. https://doi.org/10.3390/insects9040148
APA StyleShawer, R., Donati, I., Cellini, A., Spinelli, F., & Mori, N. (2018). Insecticidal Activity of Photorhabdus luminescens against Drosophila suzukii. Insects, 9(4), 148. https://doi.org/10.3390/insects9040148