Evidence for Male Horn Dimorphism and Related Pronotal Shape Variation in Copris lunaris (Linnaeus, 1758) (Coleoptera: Scarabaeidae, Coprini)


Abstract
1. Introduction
2. Materials and Methods
2.1. Material
2.2. Data Acquisition
2.3. Morphometric Measurements
2.3.1. Horn Length
2.3.2. Body Size
2.3.3. Shape
2.4. Analysis of Horn Allometry
2.5. Analysis of Shape Variation
2.6. Analysis of Size and Shape Covariation
3. Results
3.1. Horn Allometry
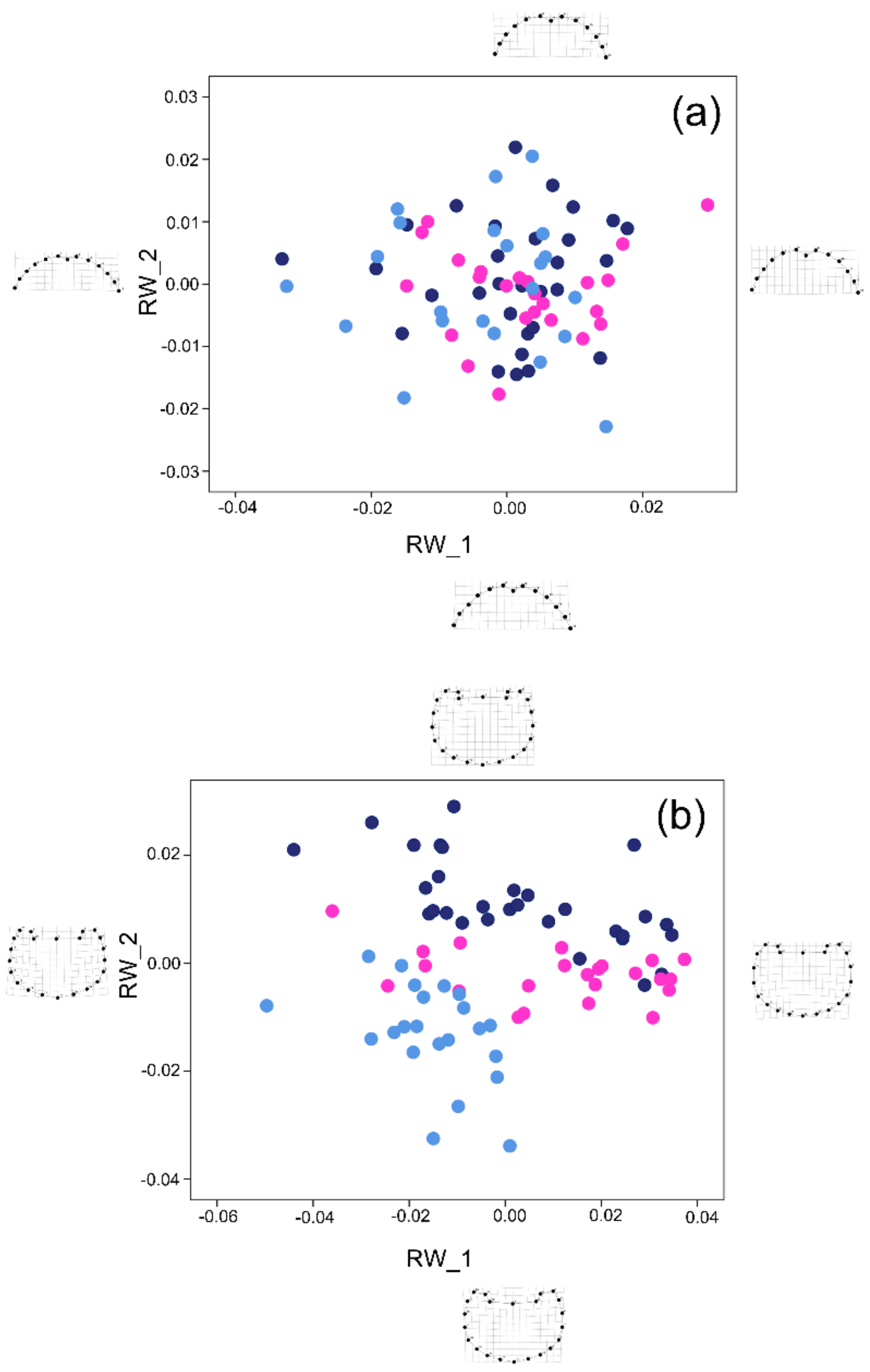
3.2. Shape Analysis
3.3. Shape vs. Size Analysis
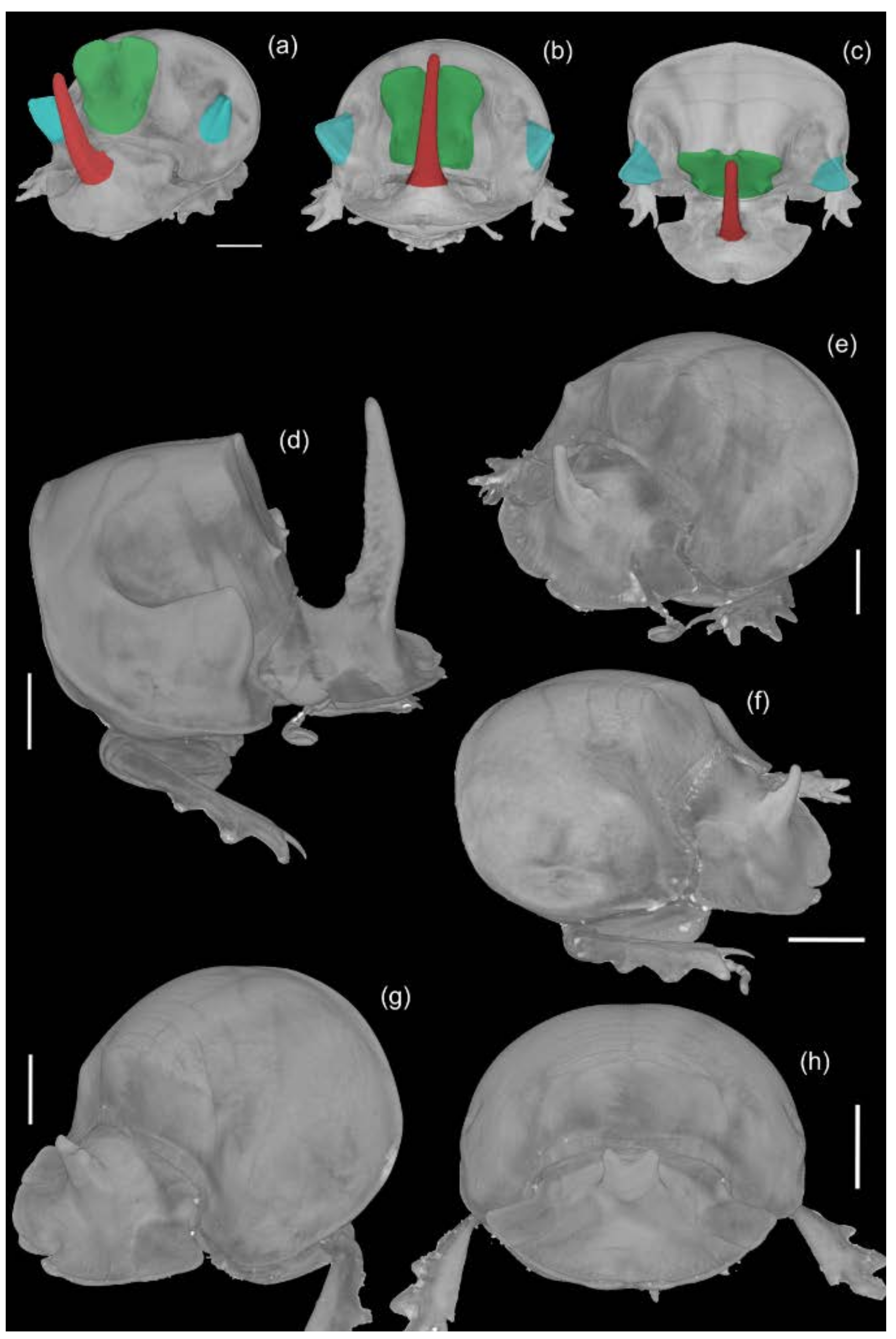
3.4. MicroCT Images
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eberhard, W.G. Beetle horn dimorphism: Making the best of a bad lot. Am. Nat. 1982, 119, 420–426. [Google Scholar] [CrossRef]
- Eberhard, W.G.; Gutiérrez, E.E. Male dimorphisms in beetles and earwigs and the question of developmental constraints. Evolution 1991, 45, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Emlen, D.J.; Nijhout, H.F. The development and evolution of exaggerated morphologies in insects. Annu. Rev. Entomol. 2000, 45, 661–708. [Google Scholar] [CrossRef] [PubMed]
- Buzatto, B.A.; Requena, G.S.; Lourenço, R.S.; Munguía-Steyer, R.; Machado, G. Conditional male dimorphism and alternative reproductive tactics in a Neotropical arachnid (Opiliones). Evol. Ecol. 2011, 25, 331–349. [Google Scholar] [CrossRef]
- Buzatto, B.A.; Machado, G. Male dimorphism and alternative reproductive tactics in harvestmen (Arachnida: Opiliones). Behav. Processes 2014, 109, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Emlen, D.J.; Hunt, J.; Simmons, L.W. Evolution of sexual dimorphism and male dimorphism in the expression of beetle horns: Phylogenetic evidence for modularity, evolutionary lability, and constraint. Am. Nat. 2005, 166 (Suppl. S4), S42–S68. [Google Scholar] [CrossRef]
- Emlen, D.J.; Lavine, L.C.; Ewen-Campen, B. On the origin and evolutionary diversification of beetle horns. Proc. Natl. Acad. Sci. USA 2007, 104, 8661–8668. [Google Scholar] [CrossRef] [PubMed]
- Emlen, D.J. Diet alters male horn allometry in the beetle Onthophagus acuminatus (Coleoptera: Scarabaeidae). Proc. R. Soc. B Sci. 1997, 264, 567–574. [Google Scholar] [CrossRef]
- Moczek, A.P.; Emlen, D.J. Proximate determination of male horn dimorphism in the beetle Ontophagus taurus (Coleoptera: Scarabaeidae). J. Evol. Biol. 1999, 12, 27–37. [Google Scholar] [CrossRef]
- Karino, K.; Seki, N.; Chiba, M. Larval nutritional environment determines adult size in Japanese horned beetles Allomyrina dichotoma. Ecol. Res. 2004, 19, 663–668. [Google Scholar] [CrossRef]
- Moczek, A.P. Horn polyphenism in the beetle Onthophagus taurus: Larval diet quality and plasticity in parental investment determine adult body size and male horn morphology. Behav. Ecol. 1998, 9, 636–641. [Google Scholar] [CrossRef]
- Hunt, J.; Simmons, L.W. Maternal and paternal effects on offspring phenotype in the dung beetle Onthophagus taurus. Evolution 2000, 54, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Emlen, D.J. Alternative reproductive tactics and male dimorphism in the horned beetle Onthophagus acuminatus (Coleoptera: Scarabaeidae). Behav. Ecol. Sociobiol. 1997, 41, 335–341. [Google Scholar] [CrossRef]
- Kotiaho, J.S.; Simmons, L.W. Longevity cost of reproduction for males but no longevity cost of mating or courtship for females in the male-dimorphic dung beetle Onthophagus binodis. J. Insect Physiol. 2003, 49, 817–822. [Google Scholar] [CrossRef]
- Gross, M.R. Alternative reproductive strategies and tactics: Diversity within sexes. Trends Ecol. Evol. 1996, 11, 92–98. [Google Scholar] [CrossRef]
- Moczek, A.P.; Emlen, D.J. Male horn dimorphism in the scarab beetle, Onthophagus taurus: Do alternative reproductive tactics favour alternative phenotypes? Anim. Behav. 2000, 59, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Simmons, L.W.; Emlen, D.J.; Tomkins, J.L. Sperm competition games between sneaks and guards: A comparative analysis using dimorphic male beetles. Evolution 2007, 61, 2684–2692. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, S.; Yamaura, Y.; Makihara, H. Sexual and Male Horn Dimorphism in Copris ochus (Coleoptera: Scarabaeidae). Zoolog. Sci. 2007, 24, 1082–1085. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, A.; Citeroni, V.; Mazzone, F.; Dellacasa, M.; Palestrini, C. Are horn morphological patterns able to differentiate the two closely related species Copris klugi Harold and Copris sierrensis Matthews? Neotrop. Entomol. 2015, 44, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Akamine, M. Dimorphism of both head and prothoracic horn morphologies in male Copris acutidens (Coleoptera: Scarabaeoidea). Entomol. Sci. 2016, 19, 82–85. [Google Scholar] [CrossRef]
- Dortel, E.; Thuiller, W.; Lobo, J.M.; Bohbot, H.; Lumaret, J.P.; Jay-Robert, P. Potential effects of climate change on the distribution of Scarabaeidae dung beetles in Western Europe. J. Insect Conserv. 2013, 17, 1059–1070. [Google Scholar] [CrossRef]
- Carpaneto, G.M.; Piattella, E.; Pittino, R. The scarab beetles of Turkey: An updated checklist and chorotype analysis (Coleoptera, Scarabaeoidea). Biogeographia 2000, 21, 217–240. [Google Scholar] [CrossRef]
- Tonelli, M.; Verdú, J.R.; Zunino, M.E. Effects of grazing intensity and the use of veterinary medical products on dung beetle biodiversity in the sub-mountainous landscape of Central Italy. PeerJ 2017, 5, e2780. [Google Scholar] [CrossRef] [PubMed]
- Klemperer, H.G. Normal and atypical nesting behaviour of Copris lunaris (L.): Comparison with related species (Coleoptera, Scarabaeidae). Ecol. Entomol. 1982, 7, 69–83. [Google Scholar] [CrossRef]
- Klemperer, H.G. Parental behaviour in Copris lunaris (Coleoptera, Scarabaeidae): Care and defence of brood balls and nest. Ecol. Entomol. 1982, 7, 155–167. [Google Scholar] [CrossRef]
- Klemperer, H.G. The evolution of parental behaviour in Scarabaeinae (Coleoptera, Scarabaeidae): An experimental approach. Ecol. Entomol. 1983, 8, 49–59. [Google Scholar] [CrossRef]
- Pizzo, A.; Mercurio, D.; Palestrini, C.; Roggero, A.; Rolando, A. Male differentiation patterns in two polyphenic sister species of the genus Onthophagus Latreille, 1802 (Coleoptera, Scarabaeidae): A geometric morphometric approach. J. Zoolog. Syst. Evol. Res. 2006, 44, 54–62. [Google Scholar] [CrossRef]
- Macagno, A.L.M.; Pizzo, A.; Roggero, A.; Rolando, A.; Palestrini, C. Horn polyphenism and related head shape variation in a single-horned dung beetle: Onthophagus (Palaeonthophagus) fracticornis (Coleoptera: Scarabaeidae). J. Zoolog. Syst. Evol. Res. 2009, 47, 96–102. [Google Scholar] [CrossRef]
- Greco, M.; Jones, A.; Spooner-Hart, R.; Holford, P. X-ray computerised microtomography (MicroCT): A new technique for assessing external and internal morphology of bees. J. Apic. Res. 2008, 47, 286–291. [Google Scholar] [CrossRef]
- Simonsen, T.J.; Kitching, I.J. Virtual dissections through micro-CT scanning: A method for non-destructive genitalia ‘dissections’ of valuable Lepidoptera materia. Syst. Entomol. 2014, 39, 606–618. [Google Scholar] [CrossRef]
- Wipfler, B.; Courtney, G.W.; Craig, D.A.; Beutel, R.G. First μCT-based 3D reconstruction of a dipteran larva—he head morphology of Protanyderus (Tanyderidae) and its phylogenetic implications. J. Morphol. 2012, 273, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Wipfler, B.; Pohl, H.; Iyavorskaya, M.; Beutel, R.G. A review of methods for analysing insect structures—the role of morphology in the age of phylogenomics. Curr. Opin. Insect Sci. 2016, 18, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Wojcieszek, J.M.; Austin, P.; Harvey, M.S.; Simmons, L.W. Micro-CT scanning provides insight into the functional morphology of millipede genitalia. J. Zool. 2012, 287, 91–95. [Google Scholar] [CrossRef]
- Lowe, T.; Garwood, R.J.; Simonsen, T.J.; Bradley, R.S.; Withers, P.J. Metamorphosis revealed: Time-lapse three-dimensional imaging inside a living chrysalis. J. R. Soc. Interface 2013, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Goyens, J.; Dirckx, J.; Aerts, P. Mechanoreceptor distribution in stag beetle jaws corresponds to the material stress in fights. Arthropod Struct. Dev. 2015, 44, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Alba-Tercedor, J. Microtomographic study on the anatomy of adult male eyes of two mayfly species. Zoosymposia 2016, 11, 101–120. [Google Scholar] [CrossRef]
- Garcia, F.H.; Fischer, G.; Liu, C.; Audisio, T.L.; Economo, E.P. Next-generation morphological character discovery and evaluation: An X-ray micro-CT enhanced revision of the ant genus Zasphinctus Wheeler (Hymenoptera, Formicidae, Dorylinae) in the Afrotropics. ZooKeys 2017, 693, 33–93. [Google Scholar] [CrossRef] [PubMed]
- Poinapen, D.; Konopka, J.K.; Umoh, J.U.; Norley, C.J.D.; McNeil, J.N.; Holdsworth, D.W. Micro-CT imaging of live insects using carbon dioxide gas-induced hypoxia as anesthetic with minimal impact on certain subsequent life history traits. BMC Zool. 2017, 2, 1–9. [Google Scholar] [CrossRef]
- Ziani, S. Morphological revision of the western Palaearctic species of the genus Copris Geoffroy, 1762 with three foretibial external teeth (Coleoptera: Scarabaeoidea: Scarabaeidae). Insecta Mundi 2017, 0528, 1–26. [Google Scholar]
- Emlen, D.J. Artificial selection on horn length-body size allometry in the horned beetle Onthophagus acuminatus. Evolution 1996, 50, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Palestrini, C.; Rolando, A. Body size and paternal investment in the genus Onthophagus (Coleoptera, Scarabaeoidea). J. Zool. 2001, 255, 405–412. [Google Scholar] [CrossRef]
- McCullough, E.L.; Ledger, K.J.; O’Brien, D.M.; Emlen, D.J. Variation in the allometry of exaggerated rhinoceros beetle horns. Anim. Behav. 2015, 109, 133–140. [Google Scholar] [CrossRef]
- Mitteroecker, P.; Gunz, P. Advances in geometric morphometrics. Evol. Biol. 2009, 36, 235–247. [Google Scholar] [CrossRef]
- Webster, M.; Sheets, H.D. A practical introduction to landmark-based geometric morphometrics. Quant. Methods Paleobiol. 2010, 16, 163–188. [Google Scholar]
- Tocco, C.; Roggero, A.; Rolando, A.; Palestrini, C. Inter-specific shape divergence in Aphodiini dung beetles: The case of Amidorus obscurus and A. immaturus. Org. Divers. Evol. 2011, 11, 263–273. [Google Scholar] [CrossRef]
- Palestrini, C.; Roggero, A.; Hernandez Nova, L.K.; Giachino, P.M.; Rolando, A. On the evolution of shape and size divergence in Nebria (Nebriola) ground beetles (Coleoptera, Carabidae). Syst. Biodivers. 2012, 10, 147–157. [Google Scholar] [CrossRef]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. A field comes of age: Geometric morphometrics in the 21st Century. Hystrix 2013, 24, 7–14. [Google Scholar] [CrossRef]
- Roggero, A.; Giachino, P.M.; Palestrini, C. A new cryptic ground beetle species from the Alps characterised via geometric morphometrics. Contrib. Zool. 2013, 82, 171–183. [Google Scholar]
- Wrozyna, C.; Neubauer, T.A.; Meyer, J.; Piller, W.E. Shape variation in neotropical Cytheridella (Ostracoda) using semilandmarks-based geometric morphometrics: A methodological approach and possible biogeographical implications. PLoS ONE 2016, 11, e0168438. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F.J. tpsDig Version 2.31; Digitize Landmarks and Outlines: Stony Brook, NY, USA, 2018; Available online: http://life.bio.sunysb.edu/morph/ (accessed on 20 July 2018).
- Rohlf, F.J. tpsUtil Version 1.76; Digitize Landmarks and Outlines: Stony Brook, NY, USA, 2018; Available online: http://life.bio.sunysb.edu/morph/ (accessed on 20 July 2018).
- Knell, R.J. On the analysis of non-linear allometries. Ecol. Entomol. 2009, 34, 1–11. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Spiess, A.N.; Neumeyer, N. An evaluation of R2 as an inadequate measure for nonlinear models in pharmacological and biochemical research: A Monte Carlo approach. BMC Pharmacol. 2010, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Moczek, A.P.; Nijhout, H.F. Rapid evolution of a polyphenic threshold. Evol. Dev. 2003, 5, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F.J. tpsRelw Version 1.69; Digitize Landmarks and Outlines: Stony Brook, NY, USA, 2017; Available online: http://life.bio.sunysb.edu/morph/ (accessed on 20 July 2018).
- Romiti, F.; Rendolfi de Zan, L.R.; Piras, P.; Carpaneto, G.M. Shape variation of mandible and head in Lucanus cervus (Coleoptera: Lucanidae): A comparison of morphometric approaches. Biol. J. Linn. Soc. Lond. 2016, 120, 836–851. [Google Scholar] [CrossRef]
- Moczek, A.P.; Nijhout, H.F. Trade-offs during the development of primary and secondary sexual traits in a horned beetle. Am. Nat. 2004, 163, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, A.; Macagno, A.L.; Dusini, S.; Palestrini, C. Trade-off between horns and other functional traits in two Onthophagus species (Scarabaeidae, Coleoptera). Zoomorphology 2012, 131, 57–68. [Google Scholar] [CrossRef]
- Piccini, I.; Arnieri, F.; Caprio, E.; Nervo, B.; Pelissetti, S.; Palestrini, C.; Roslin, T.; Rolando, A. Greenhouse gas emissions from dung pats vary with dung beetle species and with assemblage composition. PLoS ONE 2017, 12, e0178077. [Google Scholar] [CrossRef] [PubMed]
- Husak, J.F.; Swallow, J.G. Compensatory traits and the evolution of male ornaments. Behaviour 2011, 148, 1–29. [Google Scholar] [CrossRef]
- Pigliucci, M.; Preston, K. Phenotypic Integration: Studying the Ecology and Evolution of Complex Ohenotypes; Oxford University Press: Oxford, UK, 2004; pp. 1–464. ISBN 9780195160437. [Google Scholar]
- Tomkins, J.L.; Kotiaho, J.S.; Lebas, N.R. Phenotypic plasticity in the developmental integration of morphological trade-offs and secondary sexual trait compensation. Proc. Biol. Sci. 2005, 272, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Palestrini, C.; Pavan, G.; Zunino, M. Acoustic signals in Copris incertus Say (Coleoptera Scarabaeidae Coprinae). Ethol. Ecol. Evol. 1991, 3, 143–146. [Google Scholar] [CrossRef]
- Palestrini, C.; Pensati, F.; Barbero, E.; Laiolo, P. Distress signals in Copris hispanus (L., 1764) (Coleoptera: Scarabaeidae). Insect Soc. Life 2000, 3, 185–193. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean ± SE | ||||
|---|---|---|---|---|
| n | Cephalic Horn Length | Pronotal Horn Length | Max Pronotum Width | |
| females | 24 | 1.68 ± 0.27 | 1.94 ± 0.17 | 11.11 ± 0.43 |
| major males | 30 | 5.59 ± 0.80 | 3.86 ± 0.46 | 10.96 ± 0.47 |
| minor males | 22 | 1.55 ± 0.74 | 1.61 ± 0.40 | 9.50 ± 0.49 |
| Classification Cross-Validated Results | ||||||
|---|---|---|---|---|---|---|
| predicted group membership | total | |||||
| 1 | 2 | 3 | ||||
| head | count | 1 | 19 | 6 | 5 | 30 |
| 2 | 5 | 14 | 5 | 24 | ||
| 3 | 4 | 3 | 15 | 22 | ||
| % | 1 | 63.3 | 20.0 | 16.7 | 100 | |
| 2 | 20.8 | 58.3 | 20.8 | 100 | ||
| 3 | 18.2 | 13.6 | 68.2 | 100 | ||
| predicted group membership | total | |||||
| 1 | 2 | 3 | ||||
| pronotum | count | 1 | 25 | 5 | 0 | 30 |
| 2 | 0 | 21 | 2 | 23 | ||
| 3 | 0 | 1 | 21 | 22 | ||
| % | 1 | 83.3 | 16.7 | 0.0 | 100 | |
| 2 | 0.0 | 91.3 | 8.7 | 100 | ||
| 3 | 0.0 | 4.5 | 95.5 | 100 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerman, K.; Roggero, A.; Rolando, A.; Palestrini, C. Evidence for Male Horn Dimorphism and Related Pronotal Shape Variation in Copris lunaris (Linnaeus, 1758) (Coleoptera: Scarabaeidae, Coprini). Insects 2018, 9, 108. https://doi.org/10.3390/insects9030108
Kerman K, Roggero A, Rolando A, Palestrini C. Evidence for Male Horn Dimorphism and Related Pronotal Shape Variation in Copris lunaris (Linnaeus, 1758) (Coleoptera: Scarabaeidae, Coprini). Insects. 2018; 9(3):108. https://doi.org/10.3390/insects9030108
Chicago/Turabian StyleKerman, Kaan, Angela Roggero, Antonio Rolando, and Claudia Palestrini. 2018. "Evidence for Male Horn Dimorphism and Related Pronotal Shape Variation in Copris lunaris (Linnaeus, 1758) (Coleoptera: Scarabaeidae, Coprini)" Insects 9, no. 3: 108. https://doi.org/10.3390/insects9030108
APA StyleKerman, K., Roggero, A., Rolando, A., & Palestrini, C. (2018). Evidence for Male Horn Dimorphism and Related Pronotal Shape Variation in Copris lunaris (Linnaeus, 1758) (Coleoptera: Scarabaeidae, Coprini). Insects, 9(3), 108. https://doi.org/10.3390/insects9030108