Population Development of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) under Simulated UK Glasshouse Conditions

Abstract
:
1. Introduction

2. Experimental Section
2.1. Source of Tuta absoluta and Maintenance of Culture Stock
2.2. Population Development under Various Temperatures
2.3. Sexing Tuta absoluta Moths
2.4. Larval “Wandering” Time on Leaf Surface
2.5. Adult Moth Longevity When Supplied with Food Source
2.6. Data Analysis
3. Results
3.1. Population Development under Various Temperatures
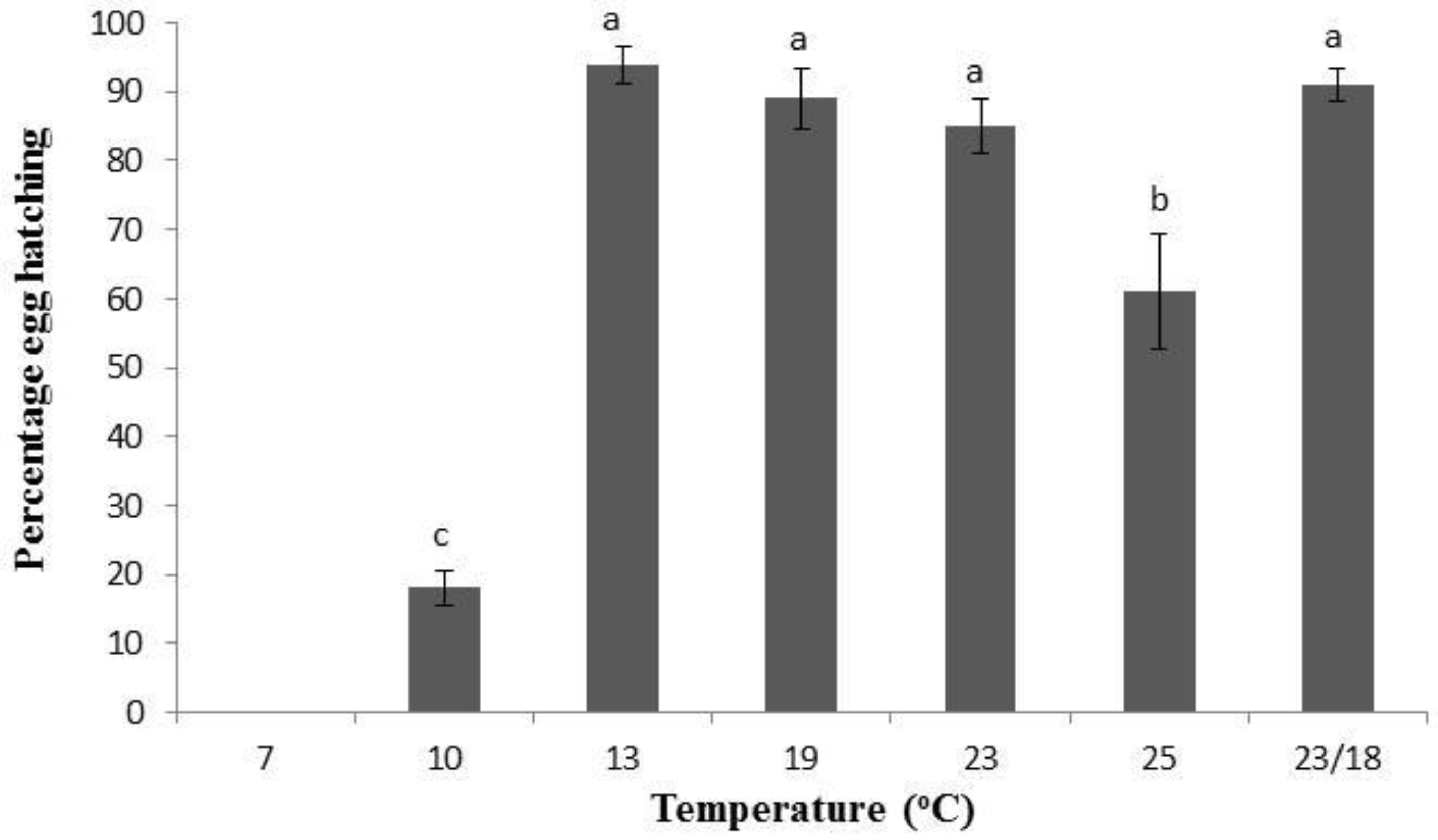
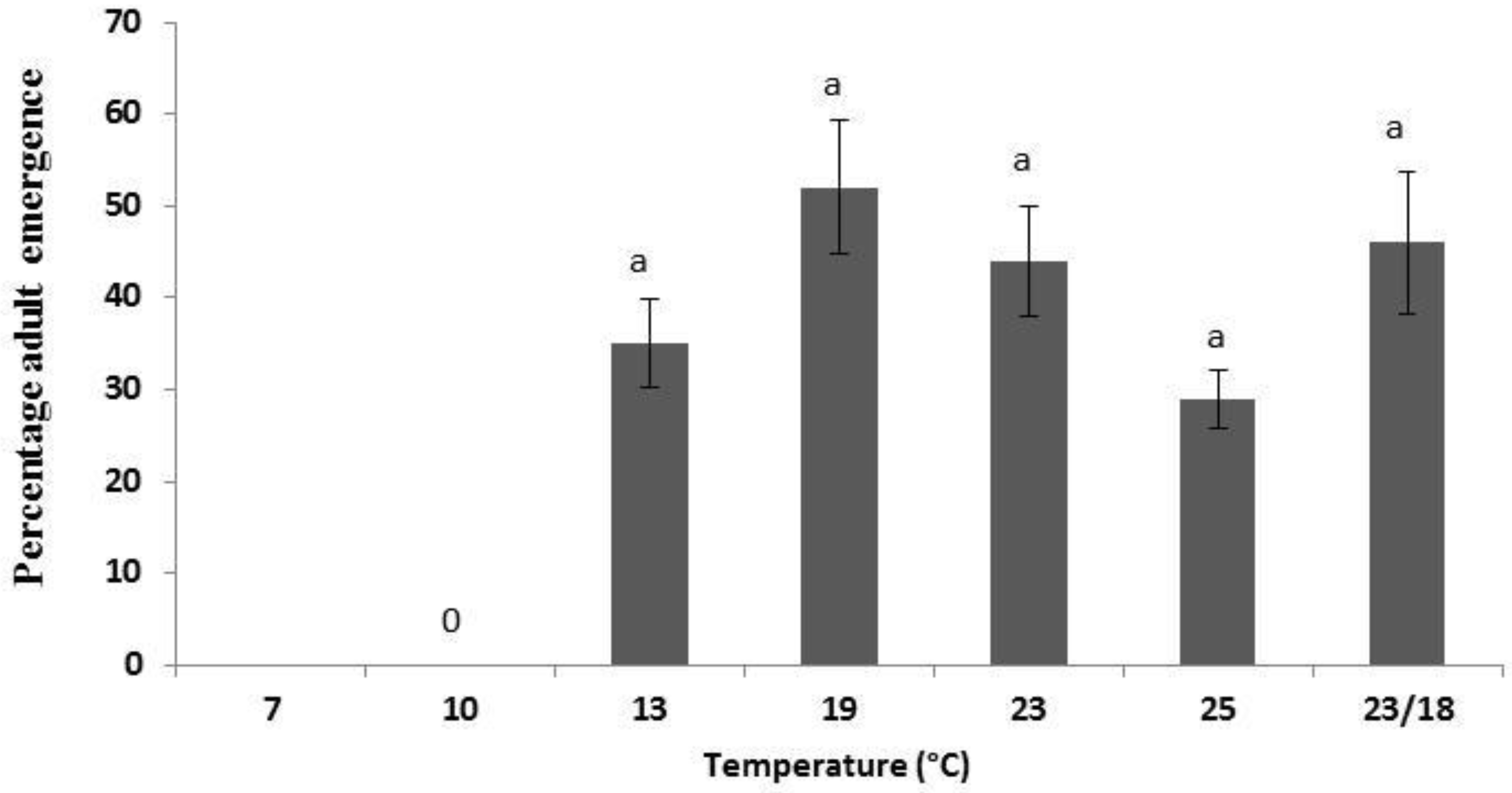

{kind=link}
{kind=link}
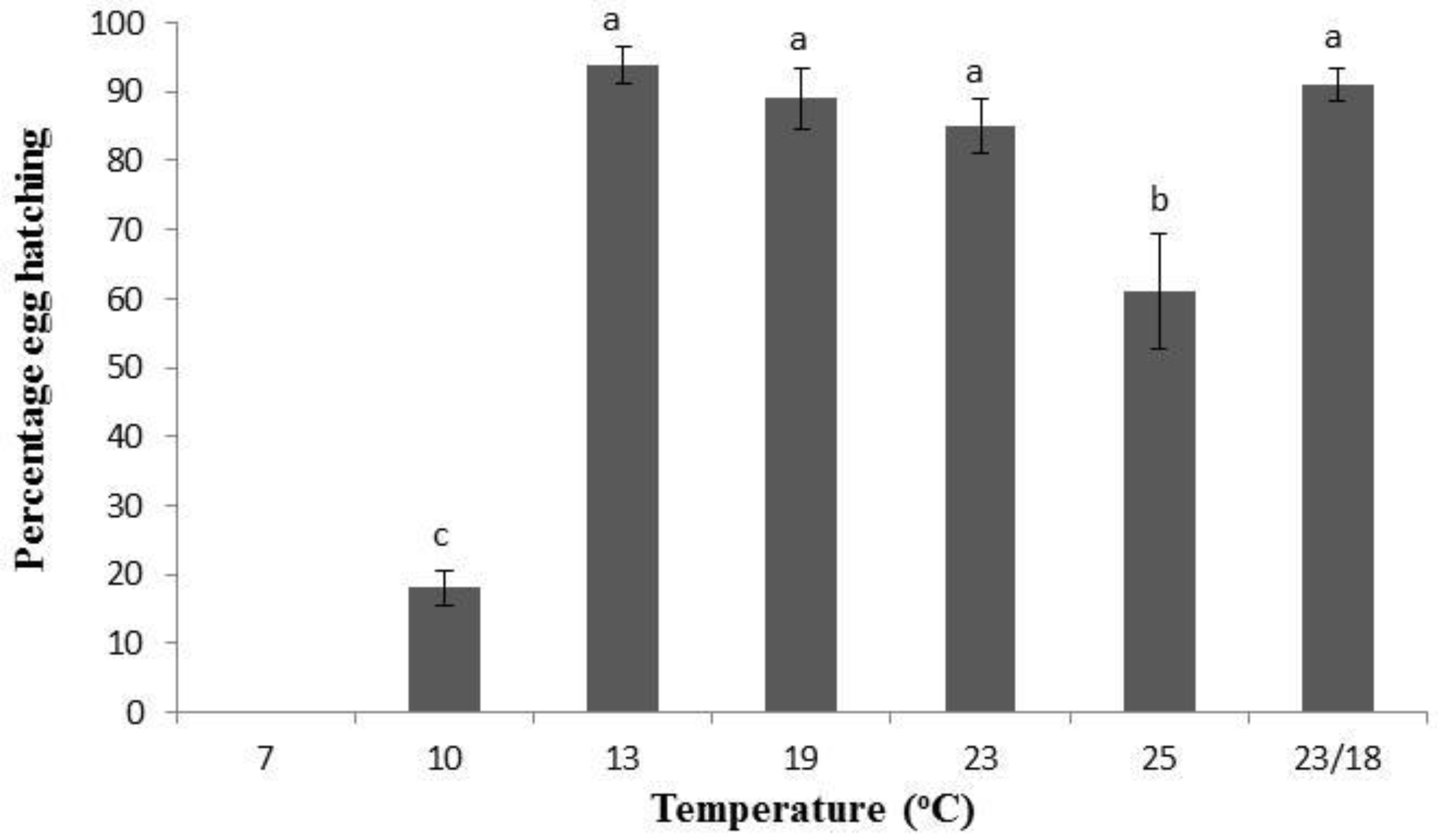{kind=link}
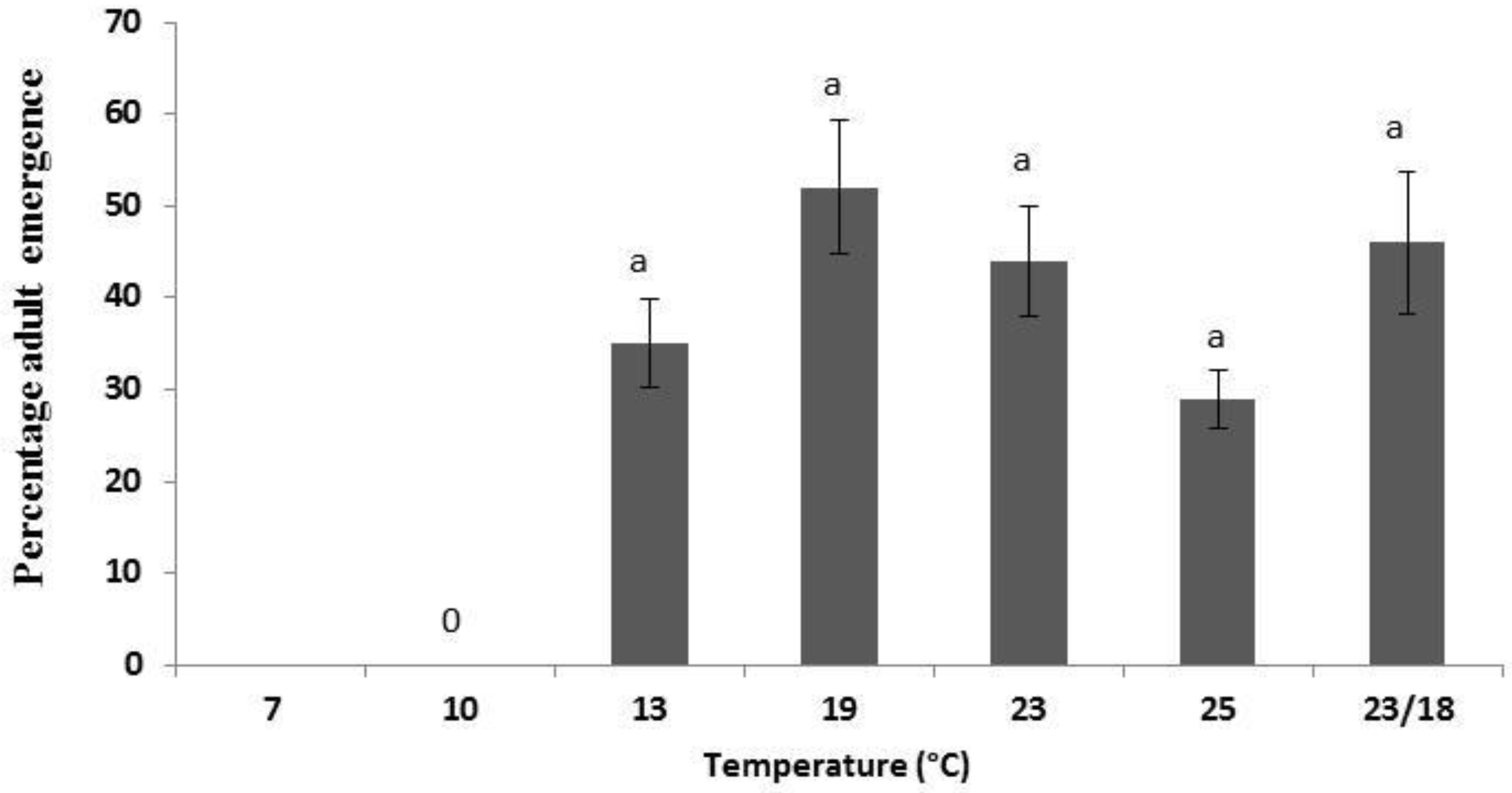{kind=link}
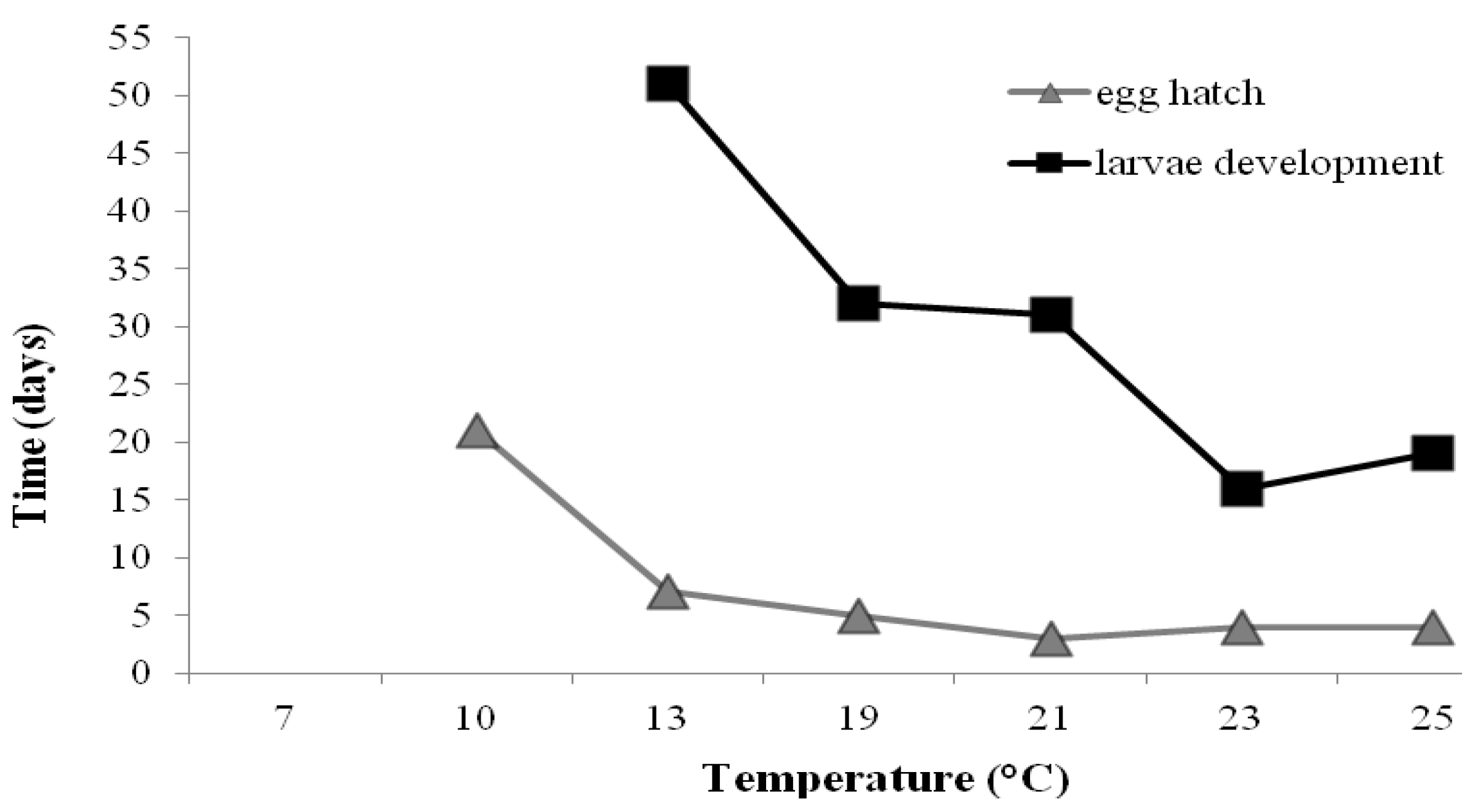
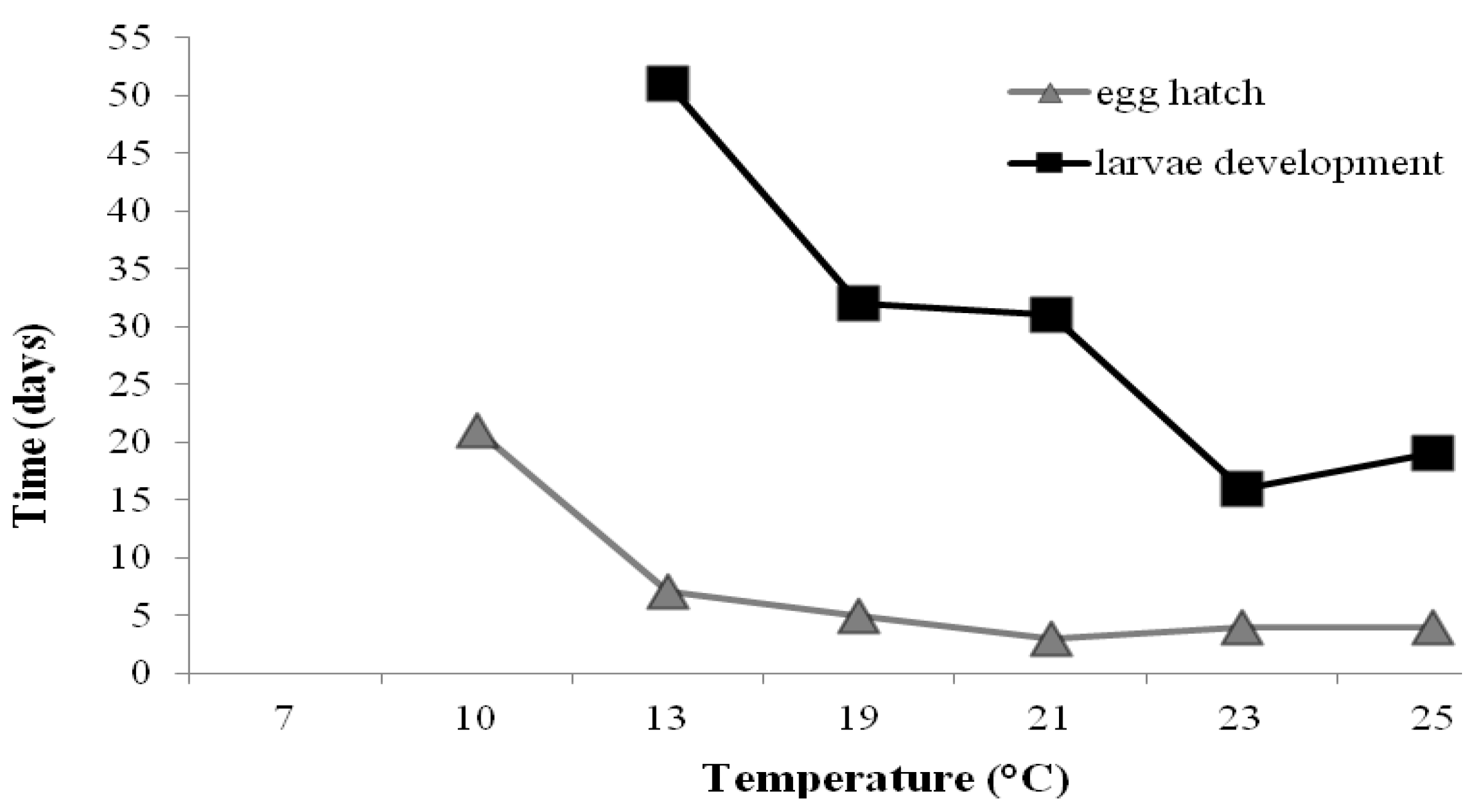{kind=link}
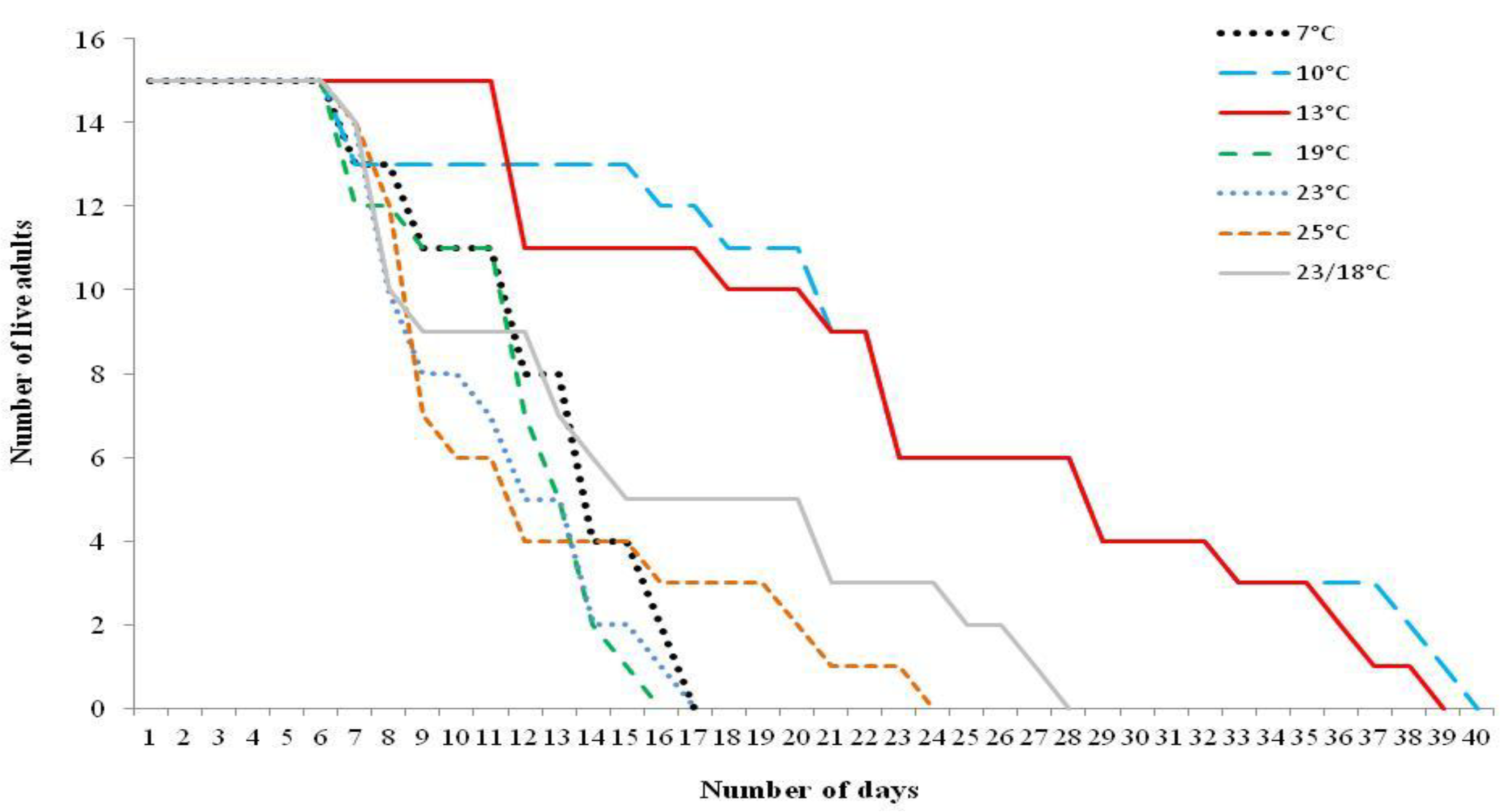
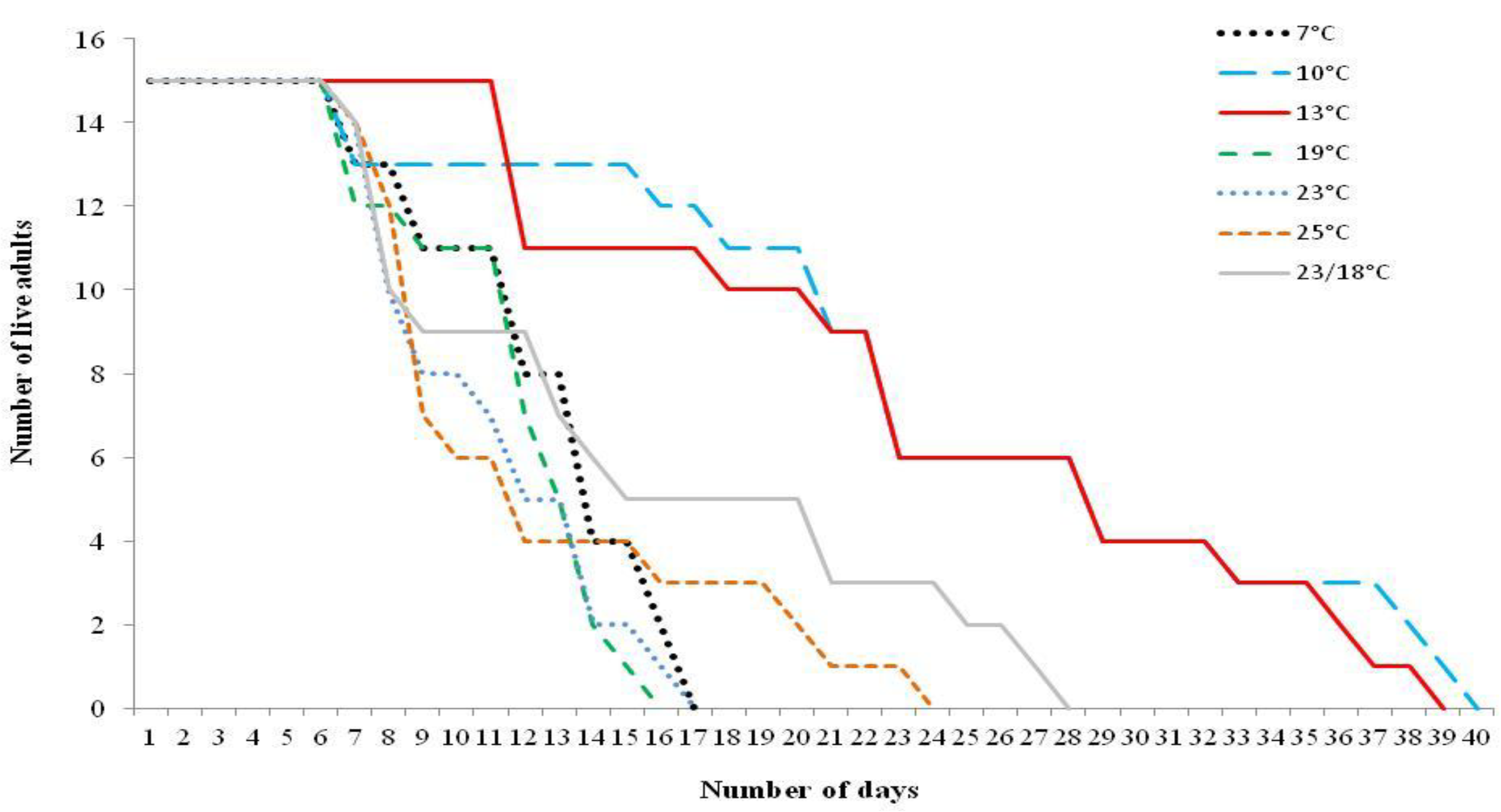{kind=link}
{kind=link}
| Temperature (°C) | Male | Female | Total |
|---|---|---|---|
| 7 | 0 | 0 | 0 |
| 10 | 0 | 0 | 0 |
| 13 | 23 a | 17 a | 40 |
| 19 | 27 a | 25 a | 52 |
| 23 | 25 a | 19 a | 44 |
| 25 | 17 a | 12 a | 29 |
| 23/18 | 18 a | 31 a | 49 |
| Total | 110 | 114 | 214 |
| Temperature (°C) | Egg hatch | Larvae | Adult | Total life-span |
|---|---|---|---|---|
| 7 | - | - | - | - |
| 10 | 21 a | 27 a,* | - | - |
| 13 | 7 b | 51 b | 14 a | 72 |
| 19 | 5 b | 32 a | 14 a | 52 |
| 23 | 4 b | 16 a | 15 a | 35 |
| 25 | 4 b | 19 a | 12 a | 35 |
| 23/18 | 3 b | 31 a | 10 a | 44 |
3.2. Adult Moth Longevity When Supplied with Food Source
3.3. Larval “Wandering” Time on Leaf Surface

4. Discussion
5. Conclusion
Acknowledgements
References
- McDonald, J.R.; Bale, J.S.; Walters, K.F.A. Temperature development and establishment potential of Thrips. palmi in the UK. Eur. J. Entomol. 1999, 95, 169–173. [Google Scholar]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Northing, P.; Mathers, J.J.; Luo, W.; Walters, K.F.A. Environmental evaluation of hot water treatments to control Liriomyza. huidobrensis infesting plant material in transit. Int. J. Environ. Sci. Technol. 2009, 6, 167–174. [Google Scholar]
- Murchie, A.K.; Moore, J.P.; Walters, K.F.A.; Blackshaw, R.P. Invasion of agricultural land by the earthworm predator, Arthurdendyus. triangulata (Deny). Pedobiologia 2003, 47, 920–923. [Google Scholar]
- Barrientos, Z.R.; Apablaza, H.J.; Norero, S.A.; Estay, P.P. Threshold temperature and thermal constant for development of the South American tomato moth, Tuta. absoluta (Lepidoptera, Gelechiidae). Ciencia. Investigacion. Agraria 1998, 25, 133–137. [Google Scholar]
- Miranda, M.M.M.; Picanco, M.; Zanuncio, J.C.; Guedes, R.N.C. Ecological life table of Tuta. absoluta (Meyrick) (Lepidoptera: gelechiidae). Biocontrol. Sci. Technol. 1998, 8, 597–606. [Google Scholar] [CrossRef]
- Torres, J.B.; Faria, C.A.; Evangelista, W.A.; Pratissol, D. Within-plant distribution of the leaf miner Tuta. absoluta (Meyrick) immatures in processing tomatoes, with notes on plant phenology. Int. J. Pest. Manage 2001, 47, 173–178. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization (EPPO). Data sheets on pests recommended for regulation as quarantine pests: Tuta Absoluta. In EPPO Bull.; 2004; 2, pp. 1–3. [Google Scholar]
- European and Mediterranean Plant Protection Organization (EPPO). Data sheets on quarantine pests: Tuta absoluta. In EPPO Bull.; 2005; 35, pp. 434–435. [Google Scholar]
- Urbaneja, A.; Vercher, R.; Navarro, V.; Garcia, M.F.; Porcuna, J.L. La polilla del tomate, Tuta. absoluta. Phytoma España 2007, 194, 16–23. [Google Scholar]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; Gonzalez-Cabrera, J.; Catalan Ruescas, D.; Tabone, E.; Frandon, J.; et al. Biological invasion of European tomato crops by Tuta. absoluta: Ecology, geographic expansion and prospects for biological control. J. Pest. Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Urbaneja, A.; González-Cabrera, J.; Arnó, J.; Gabarra, R. Prospects for biological control of Tuta. absoluta in tomatoes of the Mediterranean basin. Pest. Manag. Sci. 2012, 68, 1215–1222. [Google Scholar]
- Cuthbertson, A.G.S. The Food and Environment Research Agency (Fera): York, UK, Unpublished data. 2013.
- Haji, F.N.P.; Parra, J.R.P.; Silva, J.P.; Batista, J.G.D. Biology of the tomato leafminer under laboratory conditions. Pesqui. Agropecu. Bras. 1988, 23, 107–110. [Google Scholar]
- Lopes-Filho, F. Tomate industrial no submedio Săo Francisco e as pragas que limitam sua producao (in Portuguese). Pesqui. Agropecu. Bras. 1990, 25, 283–288. [Google Scholar]
- Apablaza, J. La polilla del tomate y su manejo Tattersal (In Spanish). Pesqui. Agropecu. Bras. 1992, 79, 12–13. [Google Scholar]
- Marris, G.; Cuthbertson, A.G.S.; Mathers, J.J.; Blackburn, L.F. Containing the Small Hive Beetle for research purposes. Bee Craft 2012, 10, 17–21. [Google Scholar]
- Cuthbertson, A.G.S.; Head, J.; Walters, K.F.A.; Gregory, S.A. The efficacy of the entomopathogenic nematode Steinernema feltiae, against instars of Bemisia tabaci. J. Invertebr. Pathol. 2003, 83, 267–269. [Google Scholar] [CrossRef]
- Jacobson, R.J. personal communication. Rob Jacobson Consultancy Ltd.: Leeds, UK, 2012. [Google Scholar]
- Tropea Garzia, G.; Siscaro, G.; Biondi, A.; Zappalà, L. Tuta absoluta, a South American pest of tomato now in the EPPO region: Biology, distribution and damage. EPPO Bull. 2012, 42, 205–210. [Google Scholar] [CrossRef]
- Price, P.W. Insect Ecology, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1984. [Google Scholar]
- Fernandez, S.; Montagne, A. Tomato leafminer biology, Scrobipalpula. absoluta (Meyrick.) (Lepidoptera: Gelechiidae). Bull. Entomol. Venez. 1990, 5, 89–99. [Google Scholar]
- Pereyra, P.C.; Sánchez, N.E. Effect of two Solanaceous plants on development and population parameters of the Tomato Leaf Miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Neotrop. Entomol. 2006, 35, 671–676. [Google Scholar] [CrossRef]
- Vercher, R.; Calabuig, A.; Felipe, C. Ecologia, muestreos y umbrales de Tuta absoluta (Meyrick) (in Spanish). Phytoma España 2010, 217, 23–26. [Google Scholar]
- Estay, P. Polilla del Tomate Tuta. absoluta (Meyrick). Available online: http://alerce.inia.cl/docs/Informativos/Informativo09.pdf (accessed on 10th April 2009).
- Curry, G.L.; Feldman, R.M.; Smith, K.C. A stochastic model of a temperature-dependent population. Theor. Popul. Biol. 1978, 13, 197–213. [Google Scholar] [CrossRef]
- Uchoa-Fernandes, M.A.; Della Lucia, T.M.C.; Vilela, E.F. Mating, oviposition and pupation of Scrobipalpula. absoluta (Meyrick) (Lepidoptera: Gelechiidae). An. Soc. Entomol. Brasil. 1995, 24, 159–164. [Google Scholar]
- Jacobson, R.J. IPM in Tomatoes. In Biological Control of Arthropod Pests in Protected Culture; Heinz, K., van Driesche, R., Parella, M., Eds.; Ball Publishing: Batavia, CA, USA, 2004; pp. 457–471. [Google Scholar]
- Kabiri, F.; Vila, E.; Cabello, T. Trichogramma. achaeae: An excellent biocontrol agent against Tuta absoluta. Sting (IOBC Newsletter on Biological Control) 2010, 33, 5–6. [Google Scholar]
- Cabello, T.; Gallego, J.R.; Vila, E.; Soler, A.; del Pino, M.; Carnero, A.; Hernandez-Suarez, E.; Polasek, A. Biological control of the South American Tomato Pinworm, Tuta. absoluta (Lep.: Gelechiidae), with releases of Trichogramma. achaeae (Hym.: Trichogrammatidae) in tomato greenhouses of Spain. IOBC/WPRS Bull. 2009, 49, 225–230. [Google Scholar]
- Cabello, T.; Gallego, J.R.; Fernandez, F.J.; Vila, E.; Soler, A.; Parra, A. Applicacion de Parasitoides de Huevos en el Control de Tuta. absoluta en Espana. In Proceedings of Phytoma Conference “La polilla del tomate, un problema en plena expansion”, Valencia, Brazil, 23–24 March 2010; pp. 53–59.
- Molla, O.; Monton, H.; Vanaclocha, P.; Beitia, F.; Urbaneja, A. Predation by the mirids, Nesidiocoris. tenuis and Macrolophus. pygmaeus on the tomato borer Tuta absoluta. IOBC/WPRS Bull. 2009, 49, 209–214. [Google Scholar]
- Urbaneja, A.; Monton, H.; Molla, O. Suitability of the tomato borer Tuta. absoluta as prey for Macrolophus. caliginosus and Nesidiocoris. tenuis. J. Appl. Entomol. 2009, 133, 292–296. [Google Scholar]
- Cabello, T.; Gallego, J.R.; Fernandez-Maldonado, F.J.; Soler, A.; Beltran, D.; Parra, A. The damsel bug (Nabis. pseudoferus (Hem.: Nabidae) as a new biological control agent of the South American Tomato Pinworm, Tuta. absoluta (Lep.: Gelechiidae), in tomato crops in Spain. IOBC/WPRS Bull. 2010, 49, 219–223. [Google Scholar]
- González-Cabrera, J.; Mollá, O.; Montón, H.; Urbaneja, A. Efficacy of Bacillus thuringiensis (Berliner) in controlling the tomato borer, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). BioControl 2011, 56, 71–80. [Google Scholar] [CrossRef]
- Biondi, A.; Desneux, N.; Siscaro, G.; Zappalà, L. Using organic-certified rather than synthetic pesticides may not be safer for biological control agents: Selectivity and side effects of 14 pesticides on the predator Orius. laevigatus. Chemosphere 2012, 87, 803–812. [Google Scholar] [CrossRef]
- Zappalà, L.; Siscaro, G.; Biondi, A.; Mollá, O.; González-Cabrera, J.; Urbaneja, A. Efficacy of sulphur on Tuta absoluta and its side effects on the predator Nesidiocoris tenuis. J. Appl. Entomol. 2012, 136, 401–409. [Google Scholar]
- Ferracini, C.; Ingegno, B.L.; Navone, P.; Ferrari, E.; Mosti, M.; Tavella, L.; Alma, A. Adaption of indigenous larval parasitoids to Tuta absoluta (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2012, 105, 1311–1319. [Google Scholar] [CrossRef]
- Zappalà, L.; Bernardo, U.; Biondi, A.; Cocco, A.; Giorgini, M.; Pedata, P.; Rapisarda, C.; Tropea Garzia, G.; Siscaro, G. Recruitment of native parasitoids by the exotic pest Tuta absoluta in Southern Italy. Bull. Insectol. 2012, 65, 51–61. [Google Scholar]
- Jacobson, R.J.; Martin, G. A potential role for entomopathogenic nematodes within IPM of Tuta absoluta (Meyrick) on organic tomato crops. IOBC/WPRS Bull. 2011, 68, 71–74. [Google Scholar]
- Put, K.; Bollens, T.; Wackers, F.L.; Pekas, A. Type and spatial distribution of food supplements impact population development and dispersal of the omnivore predator Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae). Biol. Control. 2012, 63, 172–180. [Google Scholar] [CrossRef]
- Batalla-Carrera, L.; Morton, A.; Garcia-del-Pino, F. Efficacy of entomopathogenic nematodes against tomato leafminer Tuta absoluta in laboratory and greenhouse conditions. BioControl 2010, 55, 523–530. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S. Tuta absoluta—A threat to British tomato crops. Biodivers. News 2012, 58, 11–12. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cuthbertson, A.G.S.; Mathers, J.J.; Blackburn, L.F.; Korycinska, A.; Luo, W.; Jacobson, R.J.; Northing, P. Population Development of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) under Simulated UK Glasshouse Conditions. Insects 2013, 4, 185-197. https://doi.org/10.3390/insects4020185
Cuthbertson AGS, Mathers JJ, Blackburn LF, Korycinska A, Luo W, Jacobson RJ, Northing P. Population Development of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) under Simulated UK Glasshouse Conditions. Insects. 2013; 4(2):185-197. https://doi.org/10.3390/insects4020185
Chicago/Turabian StyleCuthbertson, Andrew G. S., James J. Mathers, Lisa F. Blackburn, Anastasia Korycinska, Weiqi Luo, Robert J. Jacobson, and Phil Northing. 2013. "Population Development of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) under Simulated UK Glasshouse Conditions" Insects 4, no. 2: 185-197. https://doi.org/10.3390/insects4020185
APA StyleCuthbertson, A. G. S., Mathers, J. J., Blackburn, L. F., Korycinska, A., Luo, W., Jacobson, R. J., & Northing, P. (2013). Population Development of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) under Simulated UK Glasshouse Conditions. Insects, 4(2), 185-197. https://doi.org/10.3390/insects4020185